蕾期低溫脅迫對棉花葉綠素熒光特性的影響
2022-02-21 07:17:16鐘信念梁其干魏亦農趙建華馬海霞李志博
新疆農業科學 2022年9期
鐘信念,梁其干,魏亦農,趙建華,馬海霞,李志博
(1.石河子大學農學院/新疆兵團綠洲生態農業重點實驗室,新疆石河子 832003;2.海南省澄邁縣熱帶作物服務中心,海南澄邁 571900;3.新疆農業科學院/海南三亞農作物育種試驗中心,海南三亞 572014)
0 引言
【研究意義】低溫是影響植物生長和分布的自然災害因素之一[1-2]。棉花屬于錦葵科棉屬植物,容易受到低溫冷害的影響[3]。新疆農作物常常遭到低溫冷害[4]。低溫冷害不僅常發生在棉花的播種期和收獲期,也會發生在棉花開花結蕾期。苗期棉花溫度在15℃以下,會使棉花生長發育速度減緩或者是停止生長,后期溫度在20℃以下,對纖維成熟不利[5]。低溫使棉花生育延遲、花蕾脫落,品質和產量嚴重下降[6]。近些年在新疆棉花生長中期也出現了連續低溫冷害極端天氣[4]。【前人研究進展】低溫顯著加重了光合系統的損傷程度和光抑制,且明顯抑制了NPQ[4]。低溫脅迫下棉花幼苗的qN反應比qP靈敏[7]。植株最大光合速率隨脅迫時間延長而下降、光化學淬滅系數qP呈現先降后升趨勢[8]。5和10℃處理8 d對光合反應系統及Fv/Fm并無顯著影響;5處理下Fv/Fm隨處理時間的延長而降低,且回升至與對照相同的水平需要4 d[9]。22℃/17℃低溫下qP、Fv/Fm均呈先降后升的趨勢,而NPQ則于低溫處理第6 d達最大值[10]。胡倩等[11]以2年生油茶幼苗為試驗材料,設置(4℃和-4℃)2個低溫脅迫組,溫脅迫雖然降低了油茶葉片的SPAD值,但未達到顯著水平。植物的葉綠素熒光變化,一定程度上可以反映逆境對植物光合作用的影響[12]。低溫脅迫下4個產地綠竹的Fv、Fm、Fm/Fv和Fv/Fo均呈現下降的趨勢,Fv變化的趨勢為先降后升,葉片Fo升高[13]。【本研究切入點】關于低溫對棉花的影響已有相關研究[14-15]。目前大部分研究主要針對低溫水平對苗期棉花的影響[16-17];或研究抗寒劑包衣對棉花種子萌芽的影響[18-19]及耐寒性鑒定[20-21];有關低溫持續期與低溫強度對棉花光合參數綜合效應的研究較少。即使有少量的研究同時考慮了低溫水平和低溫持續時間的綜合效應[22],但其溫度的設置有待進一步完善。目前對植物葉綠素熒光參數受低溫脅迫影響的研究主要集中于蔬菜類和其他果樹類[23-25],而研究低溫脅迫對蕾期棉花葉綠素熒光參數影響的相關研究目前為止仍未見相關報道。需研究蕾期低溫脅迫對棉花葉綠素熒光特性的影響。【擬解決的關鍵問題】采用人工控溫、控光等條件的方法,選用棉花蕾期耐冷性相關指標,研究3個低溫梯度低溫脅迫對棉花蕾期光合特性的影響,為棉花蕾期遭遇逆境條件進行溫度調控提供理論依據。
1 材料與方法
1.1 材料
供試材料為新疆北疆不同年代代表性的3個陸地棉棉花主栽品種:新陸早61號、新陸早36號、新陸早10號,棉花種子均由新疆兵團綠洲生態農業重點實驗室棉花育種室提供。
1.2 方法
1.2.1 棉苗培養及處理
各品種挑選飽滿健康且經硫酸脫絨的種子,用0.4%的高錳酸鉀溶液對種子消毒10~15 min,用蒸餾水沖洗3~4次,沖洗后放置于經75%酒精消毒的 燒杯中備用。放置于墊有干凈濾紙(防止種子根部穿過紗布網眼)及紗布的發芽盒中,用56℃溫水泡種20~25 min后在28℃培養箱中發芽,每個發芽盒放40粒種子發芽過程中加少量水保持發芽床濕潤。
當種子芽長1 cm時移栽到盆徑(320×250)mm的花盆進行播種育苗。培養土采用花卉市場采購的花土與蛭石以4∶1的比例混配。培養光照強度設置為50 000 lx、晝夜溫度為28℃/20℃、光周期16 h、濕度為60%。待棉苗長至初始現蕾期時,分別移到同一型號的人工智能氣候箱內開始進行不同低溫脅迫處理培養。低溫強度設輕度(T1)、中度(T2)、重度(T3)3個水平,每個水平下的氣候箱培養條件為,T1光照強度20 000 lx,晝夜溫度20℃/15℃光周期16 h;T2光照強度15 000 lx晝夜溫度15℃/10℃光周期16 h;T3光照強度10 000 lx晝夜溫度10℃/5℃光周期16 h3個低溫梯度處理。以正常培養條件下為對照(CK):50 000 lx、晝夜溫度為28℃/20℃、光周期16 h、濕度為60%。每盆為1重復,每處理3次重復。每低溫強度下棉花低溫處理15 d,每隔5 d選取棉花倒4功能葉進行葉綠素熒光參數測定。圖1

圖1 不同低溫脅迫下蕾期棉花表型變化Fig.1 Effect of low temperature stress for 15 days on cotton phenotype at bud stage
1.2.2 葉綠素熒光參數指標測量
葉綠素測定采用葉綠素儀(SPAD-502,日本)。葉綠素熒光參數測定采用美國OS5-FL型飽和脈沖式葉綠素熒光分析儀(kinetic模式),測定前葉片暗適應30 min后,暴露于3 000μmol/(m2·s)(PFD)飽和脈沖光下2 s左右。分別在20℃/15℃、15℃/10℃、10℃/5℃培養0、5、10和15 d的10:00~12:00測定其各快速葉綠素熒光參數;選取蕾期棉花倒四片功能葉做標記,測定時避開葉脈。取重復測定5次的平均值。之后每次測定使用相同葉片,每枚葉片測定2次,每個處理設置6株重復。各熒光參數及具體操作參照[20]。
1.3 數據處理
使用Excel 2019進行試驗數據統計,SPSS 26.0進行差異性分析,P<0.05表示差異顯著,應用origin2021等畫圖軟件綜合分析作圖。
2 結果與分析
2.1 蕾期低溫脅迫對初始熒光(F0)的影響
研究表明,T2、T3處理下的蕾期棉花隨著低溫脅迫持續時間的延長Fo逐漸增大,且與對照相比,Fo均顯著提高,低溫脅迫使蕾期棉花PSII反應中心受到破壞。脅迫5 d時T1處理組與CK對照組之間差異不顯著。各品種在T3脅迫第5 d及新陸早61號在T2脅迫受影響較大,T2、T3低溫脅迫超過5 d可使對蕾期棉花PSII反應中心造成傷害。低溫脅迫時新陸早61號初始熒光Fo的呈現先升高后下再升高的趨勢。3種不同梯度低溫脅迫處理下,CK和T1試驗組的初始熒光(Fo)差異較小,而T2、T3的差異較大,T2、T3試驗組的初始熒光(Fo)顯著高于CK和T1試驗組,T2、T3的低溫脅迫的強度對要比蕾期棉花PSII反應中心的傷害要比T1大。圖2
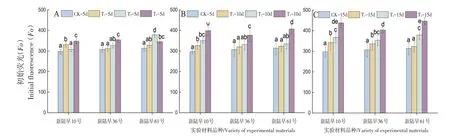
圖2 不同低溫脅迫下初始熒光Fo變化Fig.2 T Effects of low temperature stress on initial fluorescence Fo
2.2 蕾期低溫脅迫對最大熒光產量(Fm)的影響
研究表明,T1和T2處理組中隨著低溫脅迫強度的升高,蕾期棉花最大熒光產量隨之出現了先升高后降低再升高的趨勢;而在T3處理組中,蕾期棉花的最大熒光產量發生急劇下降,且不同處理天數之間差異顯著。T3處理15 d時蕾期棉花的最大熒光產量達到最低。蕾期棉花在遭受不同程度的低溫脅迫時,最大熒光產量的變化趨勢有所差異。在不同的低溫脅迫處理組中新陸早10號受到的影響最大,新陸早10號的抗性較差。T1、T2低溫處理5 d時新陸早61號發生了一次躍升,且T3處理組也大于新陸早10號、新陸早36號;而T1、T2處理5 d時新陸早10號的最大熒光產量一直較低。圖3

圖3 不同低溫脅迫下最大熒光產量Fm變化Fig.3 Effects of low temperature stress on maximum fluorescence yield Fm
2.3 蕾期低溫脅迫對可變熒光(Fv)的影響
研究表明,T1和T2處理組的可變熒光Fv大于CK組,且脅迫程度越大可變熒光Fv越大,而T3組的可變熒光Fv則顯著低于CK,蕾期棉花在T1和T2為蕾期棉花植株可調節恢復的范圍,在T1和T2脅迫強度下蕾期棉花可通過提升可變熒光Fv來增強植株的抗冷性。T3脅迫程度處理的蕾期棉花的可變熒光急劇降低,T3的脅迫程度遠遠超過了植株的抵抗范圍。
T1和T2處理15 d內和CK的可變熒光之間差異不大。當T3脅迫15 d時為負值,T3的低溫脅迫處理對蕾期棉花可造成較大的影響。T3處理組的可變熒光表現為0 d>5 d>10 d>15 d,重度低溫脅迫對蕾期棉花的影響隨低溫脅迫持續時間的延長而變大。在相同脅迫天數條件下,各處理組可變熒光呈T2>T1>CK>T3,低溫脅迫處理抑制了棉花的正常生長。圖4
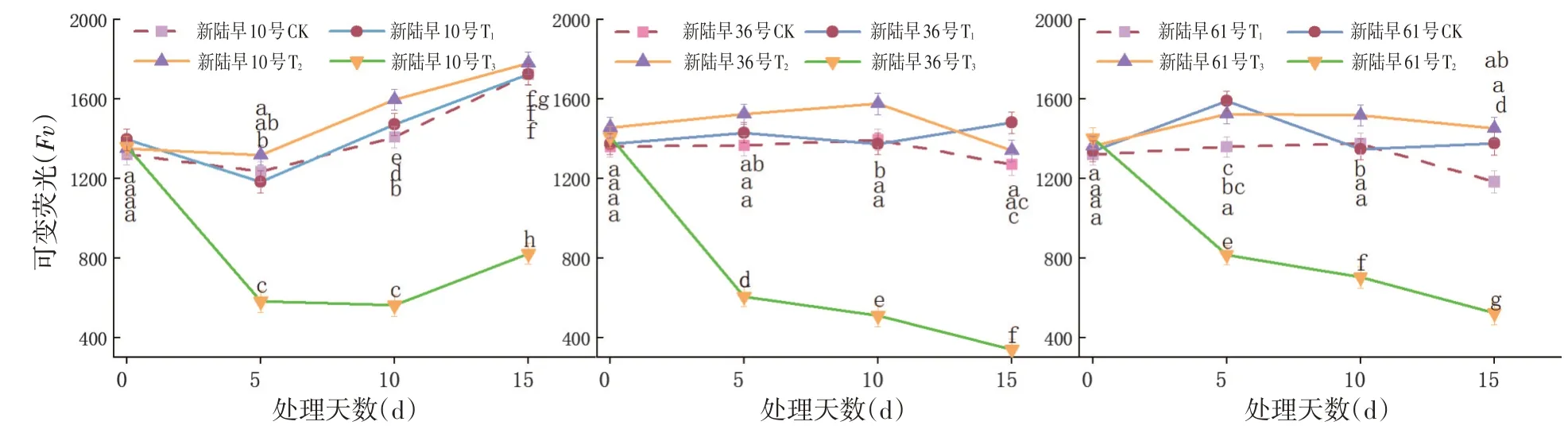
圖4 不同低溫脅迫下可變熒光Fv變化Fig.4 Effects of low temperature stress on i variable fluorescent Fv
2.4 蕾期低溫脅迫對光化學效率(Fv/Fo)的影響
研究表明,T1和T2脅迫程度下的新陸早10號、新陸早36號的光化學效率與CK組差異不大,而新陸早61號T1處理的光化學效率略大于CK。3個品種在經過T1和T2脅迫處理15 d后的光化學效率均大于CK組,T1和T2處理可使蕾期棉花通過提升光化學效率來增強植株的抗冷性,在T1和T2脅迫強度下蕾期棉花可通過提升光化學效率來增強植株的抗冷性。
T3處理組的光化學效率急劇降低,T3的脅迫程度遠遠超過了植株的抵抗范圍,且T3處理組的光化學效率表現為0 d>5 d>10 d>15 d,重度低溫脅迫對蕾期棉花的影響隨低溫脅迫持續時間的延長而變大;重度低溫脅迫對蕾期棉花的影響隨低溫脅迫持續時間的延長而變大。低溫脅迫對蕾期棉花的傷害隨著脅迫時間的延長逐漸加大,低溫脅迫處理嚴重抑制了棉花的正常生長。在相同脅迫天數條件下,各處理組可變熒光呈T2>T1>CK>T3。新陸早10號、新陸早36號T1和T2處理15 d內的光化學效率和CK的光化學效率之間差異不大。新陸早61號則表現為波動較大。圖5
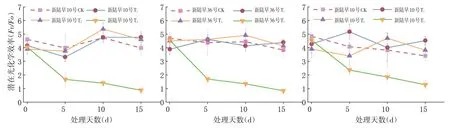
圖5 不同低溫脅迫下光化學效率(Fv/Fo)變化Fig.5 Effect of low temperature stress on potential photochemical efficiency(Fv/Fo)
2.5 蕾期低溫脅迫對花PS II最大光化學效率(Fv/Fm)的影響
研究表明,而T3處理的最大光化學效率則與CK組差異顯著。3個品種在經過T1和T2脅迫處理15 d后的最大光化學效率均與CK組差異不大,T1和T2的脅迫程度還沒有達到蕾期棉花的至死水平,而T3的脅迫程度則嚴重超過了蕾期棉花的承受水平。T1和T2脅迫程度下蕾期棉花可通過提升最大光化學效率來增強植株的抗冷性。與低溫脅迫對可變熒光、光化學效率的影響趨勢一樣。
T3處理組的最大光化學效率急劇降低,T3的脅迫程度遠遠超過了植株的抵抗范圍,且T3處理組的最大光化學效率的變化幅度表現為10 d>15 d>5 d,重度低溫脅迫對蕾期棉花的傷害在脅迫處理第10 d達到最大程度。T3處理下3個品種的蕾期棉花均隨著脅迫時間的延長最大光化學效率逐漸降低,長時間低溫脅迫導致蕾期棉花抗寒性降低,低溫脅迫處理嚴重抑制了棉花的正常生長。
在相同脅迫天數條件下,各品種的最大光化學效率變化大小表現為新陸早10號>新陸61號>新陸早36號,新陸早10號的抗冷性較差。3個品種在T1和T2處理15 d時的最大光化學效率和CK的最大光化學效率之間差異不大,其中新陸早10號不同處理組之間的的最大光化學效率波動較大。圖6
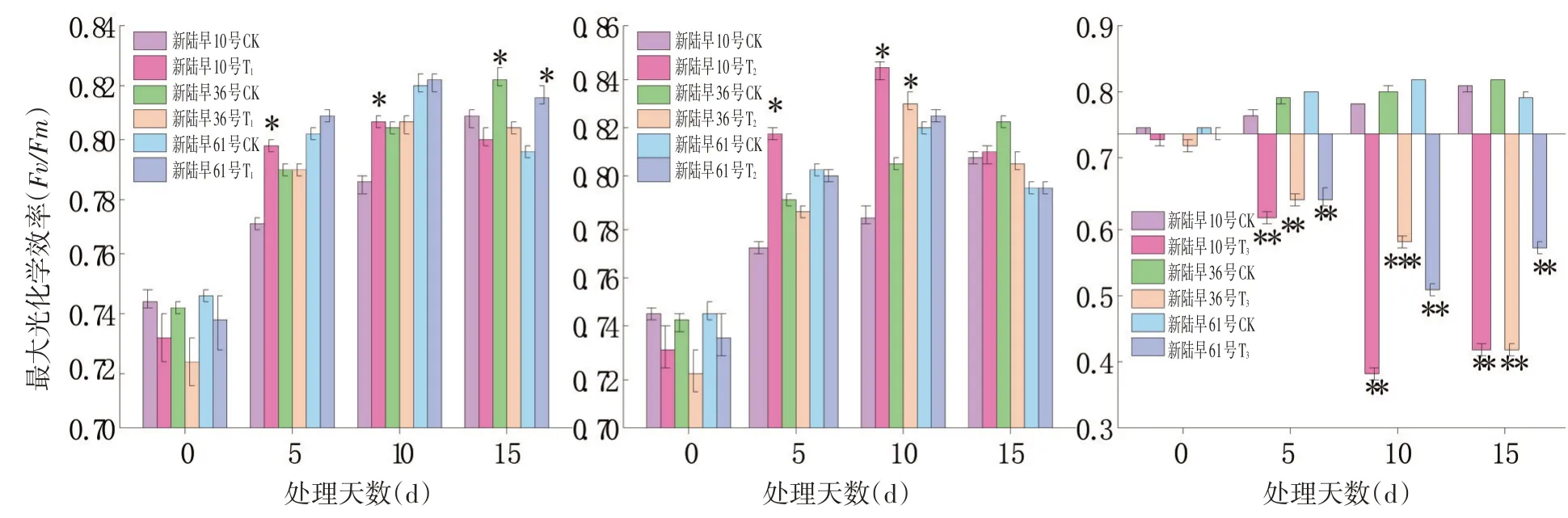
圖6 不同低溫脅迫下PSⅡ最大光化學效率(Fv/Fm)變化Fig.6 Effect of low temperature stress on maximum photochemical efficiency(Fv/Fm)of PS II
2.6 蕾期低溫脅迫對光化學猝滅速率(qP)影響
研究表明,不同低溫脅迫程度均造成3個品種蕾期棉花的光化學猝滅速率下降,光化學猝滅速率值降低,以T3-10℃/5℃條件降幅最大,T2-15℃/10℃條件次之,T1-20℃/15℃條件降幅最小。各品種在相同時段之間的光化學猝滅速率光化學猝滅速率差別不大。T1處理組下新陸早10號的光化學猝滅速率相比于CK下降最大,新陸早61號次之,新陸早36號的光化學猝滅速率下降最小;且均呈下降趨勢。T1脅迫程度下新陸早10號的抗冷性較差。
不同處理組的光化學猝滅速率值均顯著下降,并且光化學猝滅速率值下降幅度隨著溫度的降低明顯增加,低溫情況下,蕾期棉花葉片PSⅡ反應中心的開放程度降低。低溫脅迫對蕾期棉花同脅迫程度的不同品種間的qN的影響并不明顯,但和CK組相比較都呈現出顯著性差異。蕾期棉花的光化學猝滅速率表現雖然表現出了整體下降的趨勢,但各脅迫程度間均表現出顯著性的差異,低溫脅迫下的蕾期棉花通過熱耗散的形式轉移光能。新陸早36號次的光化學猝滅速率值整體下降較小,新陸早36號能夠有效抵抗低溫脅迫。圖7
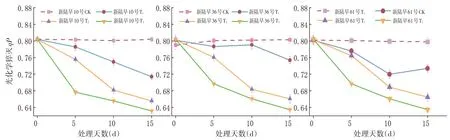
圖7 不同低溫脅迫下光化學猝滅速率qP變化Fig.7 Effect of low temperature stress on photochemical quenching rate qP
2.7 蕾期低溫脅迫對非光化學猝滅速率(NPQ)的影響
研究表明,各個處理組的非化學猝滅速率均呈上升的趨勢,低溫脅迫顯著抑制了蕾期棉花葉片的光合作用,材料在遭受低溫脅迫時啟動了自我保護機制,且通過非化學猝滅速率熱耗散等消耗多余能量的途徑避免PSⅡ功能損傷。隨著低溫脅迫時間的延長,蕾期棉花的非化學猝滅速率均持續上升,蕾期棉花具備通過熱耗散上升來自我保護的能力。低溫脅迫抑制蕾期棉花的光能利用率,促使PSⅡ的能量耗散增加,低溫脅迫能提高蕾期棉花在脅迫條件下的光能利用率。T2組的新陸早10號相比于其他兩個品種的非化學猝滅速率曲線上升速度快。T3處理組的非化學猝滅速率曲線上升幅度最大,T1組的曲線上升幅度最小;T3處理組的非化學猝滅速率急劇上升,低溫脅迫對蕾期棉花的影響隨低溫脅迫持續時間的延長及脅迫強度的加大而變大。同一個低溫脅迫處理組的非化學猝滅速率的變化幅度表現為5 d>10 d>15 d,重度低溫脅迫對蕾期棉花的傷害在脅迫處理第5和10 d最大。圖8
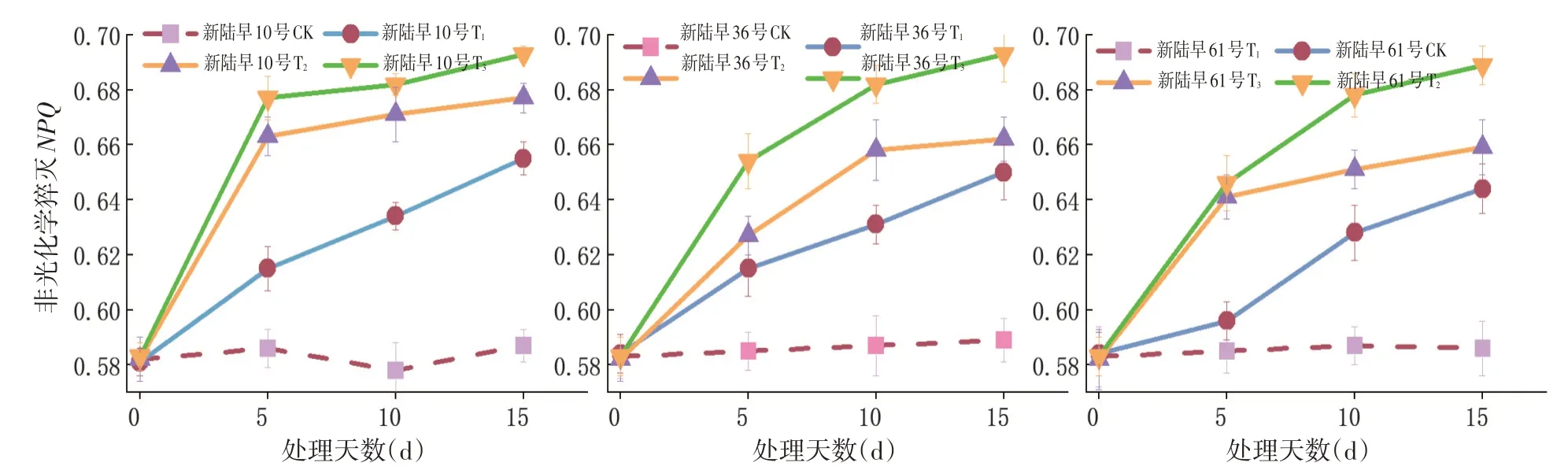
圖8 不同低溫脅迫下NPQ變化Fig.8 Effect of low temperature stress on NPQ
2.8 蕾期低溫脅迫對葉綠素(SPAD)的影響
研究表明,T1的葉綠素含量最高,T2次之,T3的葉綠素含量最低,蕾期棉花葉片的葉綠素含量隨著處理時間的延長及脅迫強度的增加,整體呈下降的趨勢。低溫脅迫強度越大葉綠素含量降低越快,低溫脅迫時間越長葉綠素含量降低幅度越大。T3的葉綠素含量顯著低于其它處理,葉片中葉綠素的含量隨低溫脅迫程度的加重而顯著降低。在相同脅迫天數條件下,各處理組葉綠素含量為T1>T2>T3,蕾期棉花在低溫脅迫下葉綠素受到嚴重的破壞。低溫脅迫強度越大蕾期棉花葉片的葉綠素含量越降低,低溫脅迫處理抑制了蕾期棉花葉片的正常生長。
對照棉花葉片的葉綠素值一直呈升高的趨勢,但在處理后的第9 d,與處理前相比才表現出極顯著差異。而脅迫棉花葉片的葉綠素值卻呈下降的走勢,且第3 d其葉綠素值就顯著下降。對照和脅迫棉花葉片的葉綠素值第1 d達到顯著差異,第3 d就達到了極顯著下差異,棉花葉片的葉綠素值對低溫弱光很敏感。圖9
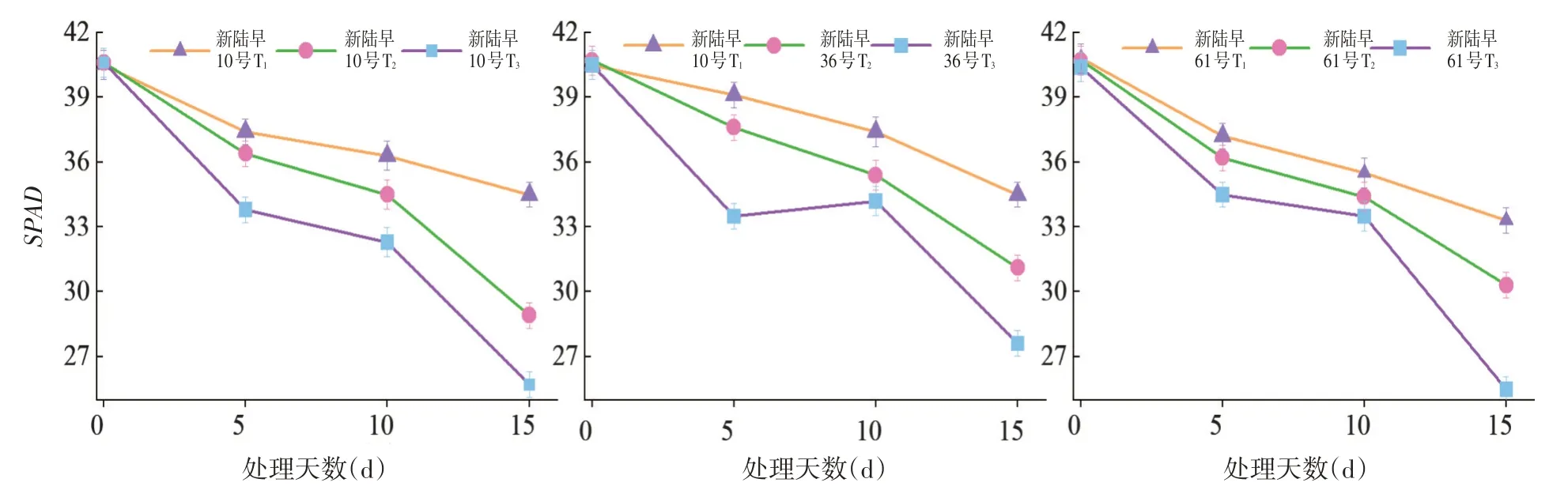
圖9 不同低溫脅迫下葉綠素SPAD變化Fig.9 Effects of low temperature stress on chlorophyll SPAD
3 討論
研究表明,新陸早61號對T2脅迫5 d至10 d有一定的耐受范圍,而當脅迫時長達到15 d時則超過蕾期棉花對低溫脅迫的耐受范圍。當低溫脅迫時間的延長到10、15 d時,T2、T3的Fo隨著低溫脅迫持續時間的延長而愈加增大。王兆[26]通過對3個彩葉草品種進行低溫脅迫研究,發現Fv/Fm和PSII都有不同程度的下降,此現象為彩葉草PSII結構受到了低溫脅迫的傷害。與研究中蕾期棉花在低溫脅迫時Fv/Fm下降的結果一致。研究蕾期棉花在輕度、中度低溫脅迫時Fv/Fm均大于CK組(呈升-降趨勢),重度低溫脅迫時Fv/Fm則小于CK組(呈大幅下降-小幅上升趨勢)且差異極其顯著;Fv/Fo輕度、中度低溫脅迫時與CK組差異不明顯(呈降-升-降趨勢),重度低溫脅迫時Fv/Fm則低于CK組(呈急劇下降趨勢)且差異極其顯著,與已有的研究4份諾麗種質幼苗葉片的Fv/Fo和Fv/Fm隨著脅迫強度加大,呈降-升-降趨勢,不同處理間差異顯著的研究結果基本相一致[25],差異可能是由于實驗材料及低溫設置不同導致。
王春萍等[27]研究發現,低溫弱光處理5 d后辣椒幼苗的SPAD值顯著下降。實驗中蕾期棉花SPAD值同樣隨低溫脅迫時間的延長及脅迫強度增加呈下降趨勢。(25、12、9、6、3、0℃)低溫處理1、3、5 d下,諾麗幼苗葉片非光化學猝滅系數(NPQ)均顯著升高[25];與研究中遭遇低溫脅迫時NPQ升高的結果一致。但也有對咖啡在8℃和0℃低溫脅迫處理72 h進行的研究,隨著處理溫度的降低和時間的延長,咖啡葉片的非光化學浮滅系數(NPQ)呈下降趨勢[28]。以上出現NPQ大幅度降低的現象可能是由于該實驗設置的溫度過低引起的。
郭菊蘭等[13]研究發現5℃低溫脅迫下,可變熒光(Fv)低于CK,且隨低溫時間的延長呈現下降趨勢,在低溫第5 d,顯著低于CK(P<0.05)。與研究中中度、重度脅迫下可變熒光(Fv)呈現下降趨勢的現象相一致。越冬低溫脅迫下可變熒光(Fv)表現為先下降后上升、最大熒光(Fm)表現為先下降后上升[23];與研究中輕度脅迫下可變熒光(Fv)同樣表現為先下降后上升的趨勢相一致;而與本研究中最大熒光(Fm)在輕度、中度脅迫下最大熒光(Fm)先上升后下降,重度脅迫下最大熒光(Fm)呈現逐漸下降趨勢的結果有出入,不同的實驗材料,最大熒光(Fm)、可變熒光(Fv)對低溫脅迫的響應趨勢不一致。
4 結論
4.1 可根據蕾期棉花的Fm、Fo等熒光參數來判斷其受到的低溫脅迫程度。
4.2 低溫脅迫對蕾期棉花的生長有抑制的作用,脅迫溫度越低,對蕾期棉花生長的抑制作用越明顯,且相同低溫度值處理下,低溫處理的時間持續越長,對蕾期棉花生長的抑制作用越明顯。T1脅迫時新陸早61號的耐低溫性較好,T2、T3脅迫時新陸早36號相比于新陸早61號和新陸早10號耐低溫性更好。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
甘肅教育(2020年14期)2020-09-11 07:57:42
中學生數理化(高中版.高考數學)(2020年5期)2020-06-02 09:19:08
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
商周刊(2017年9期)2017-08-22 02:57:49
時代英語·高二(2015年1期)2015-03-16 00:08:11
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
中國衛生(2014年11期)2014-11-12 13:11:32
