葉爾羌河流域長身高原鰍寄生線蟲的感染情況及分布類型
2022-02-23 07:57:26田勝利郝翠蘭張文潤容夢婕穆妮熱喀迪爾馮龍基高仕文
淡水漁業 2022年1期
田勝利,郝翠蘭,張文潤,容夢婕,穆妮熱·喀迪爾,馮龍基,高仕文,張 航,岳 城
(新疆農業大學動物醫學學院,烏魯木齊 830052)
葉爾羌河發源于喀喇昆侖山脈,為冰雪補給型河流,是我國最大內陸河塔里木河的源流之一。葉爾羌河流域位于新疆南部,東鄰塔克拉瑪干沙漠,西為帕米爾高原,南界為喀喇昆侖山脈,北與天山南麓沖擊扇相接,屬干旱半干旱氣候。由于其特殊的地理位置及氣候,生存著一些特殊的土著魚類,如塔里木裂腹魚()、寬口裂腹魚()、隆額高原鰍()、長身高原鰍()等。長身高原鰍隸屬于鯉目()鰍科()條鰍亞科()高原鰍屬(),對高原的適應能力極強,廣泛分布于新疆塔里木河水系和甘肅的黑河等,在分布地是優勢種群,對維護生態環境穩定起著重要作用。近年來,由于外來物種帶入,人為干擾,長身高原鰍等特有土著魚類瀕危。目前關于長身高原鰍的報道主要集中于生物學、生長特性、人工繁殖,關于寄生蟲病未見報道。
桿咽線蟲(sp.)隸屬于尾感器亞綱旋尾目桿咽科桿咽屬,而魯道夫對盲囊線蟲()隸屬于尾感器亞綱蛔目異尖科對盲囊屬。桿咽線蟲和對盲囊線蟲在高原鰍魚類中廣泛寄生,對魚體組織和器官造成輕微損傷,使其他病菌侵入,從而使魚致病、甚至死亡,而且有些對盲囊線蟲是人獸共患疾病的病原,對人類健康有危害。本研究對長身高原鰍體內寄生線蟲的感染情況及分布類型進行研究,旨在闡明長身高原鰍寄生線蟲的感染情況及種群分布情況,為長身高原鰍的人工繁育和物種保護提供線蟲病防控的基礎資料。
1 材料與方法
1.1 樣本采集與鑒定
于2019年5月-2020年7月分3次對葉爾羌河塔什庫爾干塔吉克自治縣河段進行采樣,其中捕獲到長身高原鰍414尾。參照《魚病調查手冊》的方法對魚體進行編號、測量、檢查、蟲體采集、保存等。依據《新疆魚類志》對魚類樣本進行形態鑒定,用乳酚透明法進行線蟲制片,依據《魚類寄生蟲學》進行形態鑒定。根據文獻[14-16]對魚類和線蟲進行分子生物學鑒定。
1.2 長身高原鰍體長分組
參考文獻[17]將414尾長身高原鰍分為五個體長組:≤8 cm、8 cm<≤10 cm、10 cm<≤12 cm、12 cm<≤14 cm、>14 cm。
1.3 統計方法
參考BUSH等方法,統計不同體長段長身高原鰍感染率、感染強度、平均豐度、方差、均值、方均比、負二項參數、擴散性指數、平均擁擠度等數據,分析線蟲在長身高原鰍體內的感染情況,種群空間分布格局判斷指標見表1。單因素方差分析(One-way ANOVA)比較各體長組平均感染強度水平(>0.05,差異不顯著;0.01<<0.05,差異顯著;<0.01,差異極顯著)。

表1 種群空間分布判別指標
線蟲的種群平均豐度()與宿主體長()的關系用冪函數=(,為參數)進行擬合。相關公式的擬合程度由相關系數來判斷。相關數據分析和構圖借助SPSS21.0、EXCEL、Originpro 2018、Graphpad prism 8軟件。
1.4 頻率分布統計
預先設定并統計感染0、1~4、5~8、9~12、13~16、17~20和20條以上線蟲的各體長組宿主受感染的百分率。
2 結果和分析
2.1 感染情況
從捕獲的414尾長身高原鰍中共檢獲線蟲2種,分別為桿咽線蟲和魯道夫對盲囊線蟲。線蟲的總感染率為53.38%,平均感染豐度為(7.00±0.99),平均感染強度為(13.10±0.56),共檢出線蟲2 895條,單尾檢出蟲體最多為164條(表2)。桿咽線蟲的感染率、平均感染強度、平均感染豐度均為最高,魯道夫對盲囊線蟲感染程度較輕。
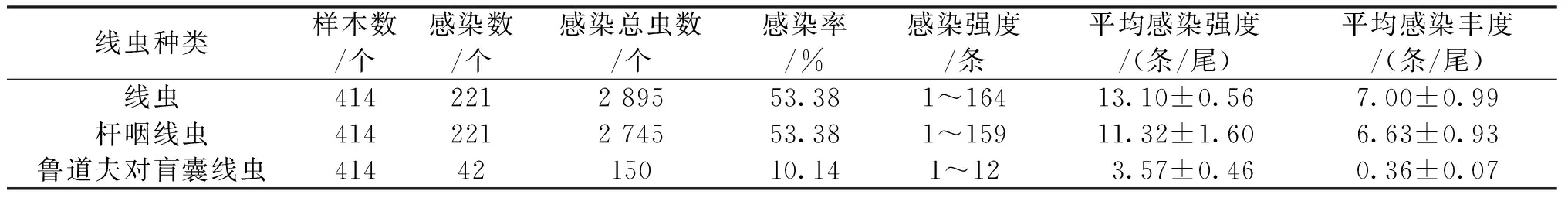
表2 長身高原鰍腸內寄生線蟲的感染情況
2.2 不同體長宿主中桿咽線蟲和魯道夫對盲囊線蟲的感染情況
單因素方差分析比較長身高原鰍各體長組中線蟲感染強度差異,桿咽線蟲感染強度差異性極顯著(=0<0.01),魯道夫對盲囊線蟲感染強度差異性不顯著(=0.302>0.05)。由表3可知桿咽線蟲感染強度和感染豐度均隨著宿主體長的增加而增加,在>14 cm組取得最大值;感染率則呈現波動變化,在>14 cm取得最大值。由表3中的宿主體長值和桿咽線蟲平均感染豐度作非曲線擬合得:=7266 69×10,=4696 17,=0987 16,相關系數在0.01檢驗水平顯著,因而桿咽線蟲在長身高原鰍中的種群平均豐度()與宿主體長()的關系可表示為:=7266 69×104696 17。

表3 桿咽線蟲在不同體長組長身高原鰍的感染情況
由表4可知魯道夫對盲囊線蟲的感染豐度和感染率表現出相同的變化趨勢,均隨著宿主體長的增加而增加,在>14 cm組取得最大值;感染強度則呈現波動變化,在12 cm<≤14 cm組取得最大值。由表4中的宿主體長值和魯道夫對盲囊線蟲平均感染豐度作非曲線擬合得:=4320 85×10,=4669 67,=0965 04,相關系數在005檢驗水平顯著,因而對盲囊線蟲在長身高原鰍中的種群平均豐度()與宿主體長()的關系可表示為:=4320 85×104669 67。

表4 魯道夫對盲囊線蟲在不同體長組長身高原鰍的感染情況
2.3 不同體長宿主中桿咽線蟲和魯道夫對盲囊線蟲的頻率分布
桿咽線蟲和魯道夫對盲囊線蟲在不同體長長身高原鰍中的頻率分布見表5、表6。不同體長宿主均感染桿咽線蟲,未感染桿咽線蟲的比例在>10 cm中隨著宿主體長的增加而減少;而感染20條以上桿咽線蟲的比例隨著宿主體長增加而增大;感染17~20條桿咽線蟲的宿主所占比例最小。在長身高原鰍不同體長組中,未感染魯道夫對盲囊線蟲的宿主占比最高;感染l~4條魯道夫對盲囊線蟲的宿主所占比例次之;未發現感染13條線蟲以上的宿主;感染1~4和5~8條的線蟲隨著宿主體長的增加而頻率增加。
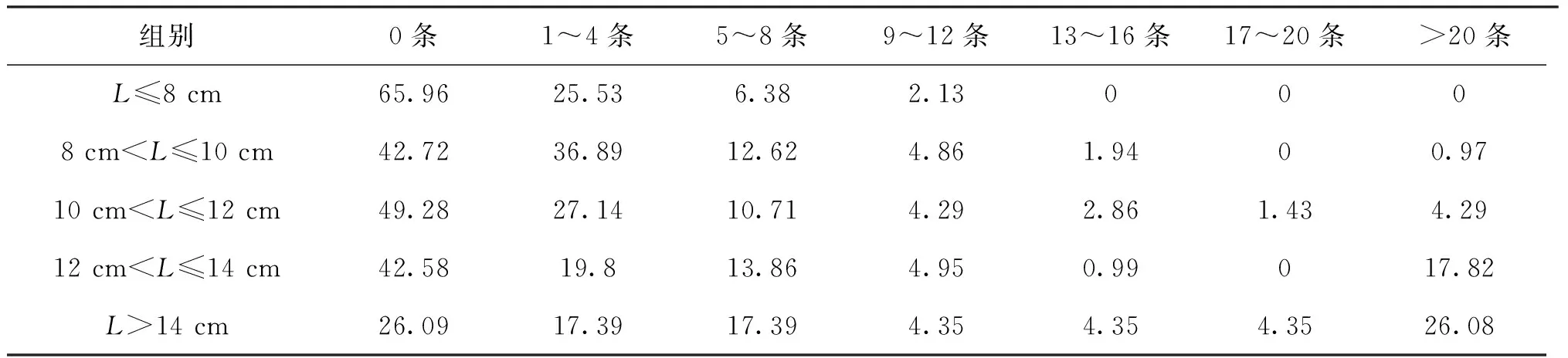
表5 桿咽線蟲在不同體長組宿主的頻率變化
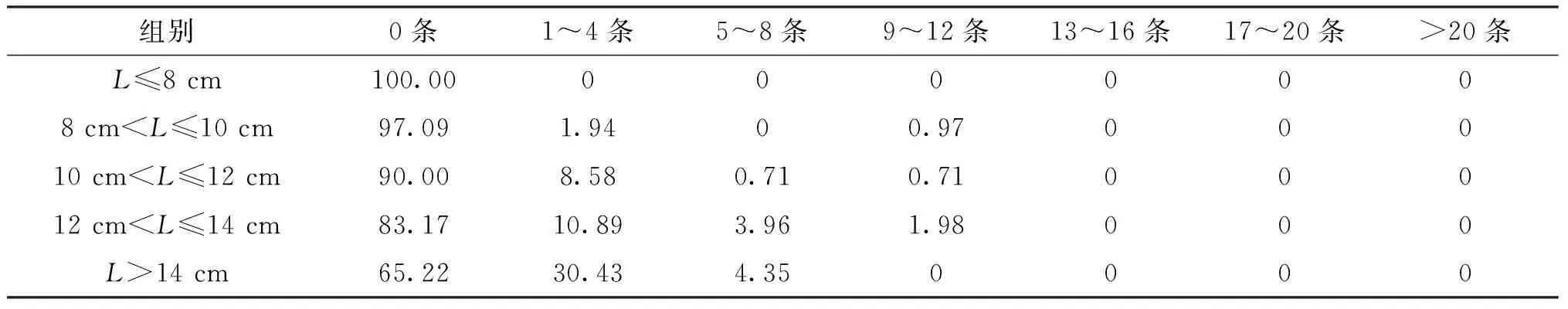
表6 魯道夫對盲囊線蟲在不同體長組宿主的頻率變化
2.4 不同體長宿主中桿咽線蟲和魯道夫對盲囊線蟲的種群空間分布
由表7,表8可知,在各體長段宿主中,桿咽線蟲和魯道夫對盲囊線蟲的>1,>0,表明桿咽線蟲和魯道夫對盲囊線蟲在各體長組長身高原鰍中呈聚集分布。

表7 桿咽線蟲在長身高原鰍種群中的聚集強度參數
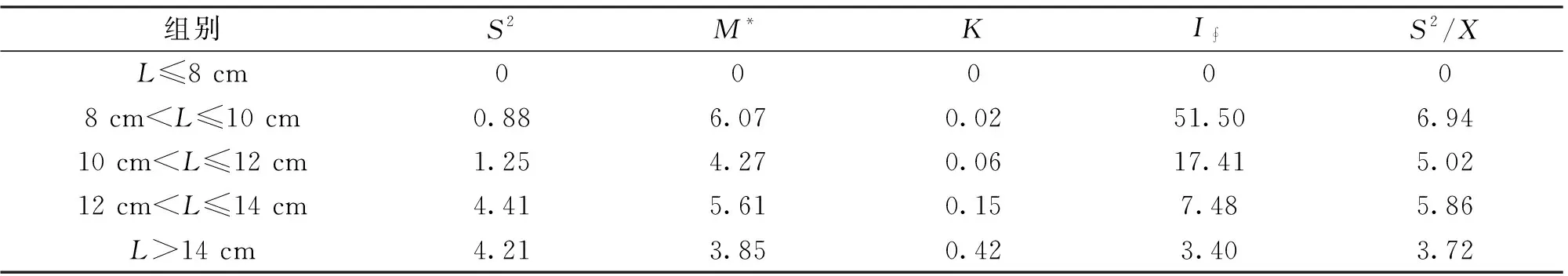
表8 魯道夫對盲囊線蟲在長身高原鰍種群中的聚集強度參數
桿咽線蟲在長身高原鰍各體長的宿主群中,、隨著宿主體長的增加而增大,表明桿咽線蟲在宿主分布的聚集強度隨著宿主體長的增加而增大。魯道夫對盲囊線蟲在大于12 cm的宿主群中,、隨著宿主體長的增加而減小,這說明魯道夫對盲囊線蟲在宿主分布的聚集強度隨著宿主體長的增加而減少。
2.5 桿咽線蟲和魯道夫對盲囊線蟲的種群空間格局與平均豐度關系
由表7中的和表3中的值,經回歸分析得:=23825 5,=1336 4,=0997 8,相關關系在001檢驗水平顯著,方差與平均豐度的關系可表示為=23.825 5。由Taylor判斷法則,說明桿咽線蟲種群在長身高原鰍體內呈聚集分布,并且聚集度有密度制約性。對于呈聚集分布的格局,可以用lawo法則進行判斷。由表 3中和表7的值,作回歸分析得:=10656 3,=3975 3,=0916 6,相關關系在005檢驗水平顯著,可表示為=10656 3+3975 3。>0,>1,說明桿咽線蟲在長身高原鰍的宿主種群中為集群數的鄰接分布,由于這種分布具有復雜性,尚未有合適的數學模型進行描述。
魯道夫對盲囊線蟲由表8中的和表4中值,經回歸分析得:=4.318 6,=0.702 2,=0.942 3,相關關系在0.05檢驗水平顯著,方差與平均豐度的關系可表示為S=4.318 6M。由Taylor判斷法則,魯道夫對盲囊線蟲種群在長身高原鰍體內為均勻分布,但是平均豐度小于1時,Taylor法則并不適用,而且由負二項參數、方均比等參數和頻率分布結果均判斷為聚集分布。
3 討論
3.1 長身高原鰍寄生線蟲的感染情況
本研究中桿咽線蟲感染率、感染豐度比姚衛健等調查新疆伊犁河斯氏高原鰍感染峨邊桿咽線蟲高,且發現幼蟲較多。兩種高原鰍的桿咽線蟲感染豐度、感染率的差異主要與宿主的食性有關,桿咽線蟲依靠中間宿主(蜉蝣動物和橈足類)進行傳播,長身高原鰍以動物性食料為主,而斯氏高原鰍則以植物性食料為主,這也就意味著長身高原鰍可以獲得更多通過食物鏈傳播的桿咽線蟲。另一方面,本研究采樣主要集中在5-7月,處于桿咽線蟲幼蟲的感染期。CASPETA對桿咽線蟲的發生和成熟的季節動態進行了長期觀察,研究結果表明,根據當地的生態條件,這些線蟲可以全年或僅在一定時期內產卵,在溫帶地區大多在春季和夏季。此外,幼蟲較多也可能與葉爾羌河水溫有關,葉爾羌河流域年均氣溫6 ℃,水溫較低,會導致線蟲在中間宿主、終末宿主中發育較慢,成熟期推遲,這與其他學者所觀察到的結果一致。
本研究中魯道夫對盲囊線蟲感染率、感染豐度比楊廷寶等調查青海湖裸鯉寄生對盲囊線蟲結果低,這主要與第一中間宿主的鋸緣真劍水蚤以及終末宿主葉爾羌河流域食魚鳥類的數量較少有關,導致對盲囊線蟲感染少量長身高原鰍。
長身高原鰍寄生桿咽線蟲和魯道夫對盲囊線蟲感染程度的差異主要受宿主食性分化和對蟲種易感性、免疫力以及第一中間宿主種群數量等影響。另一方面,兩種線蟲占據同一生態位,寄生蟲競爭能力的種間差異起著重要的作用,而且這種競爭能力的差異會隨著季節而變化。
3.2 不同體長宿主桿咽線蟲和魯道夫對盲囊線蟲感染情況
本研究結果表明,長身高原鰍寄生魯道夫對盲囊線蟲和桿咽線蟲的平均感染豐度均隨著長身高原鰍體長增加而增大。結合兩種線蟲的頻率分布數據,結果說明,多數宿主未感染或感染較少線蟲,少數宿主感染較多線蟲,且感染線蟲數量隨宿主體長的增加而增大,這一結果與MORAVEC等和王娜等所觀察到的結果一致。導致這種現象的原因有很多,首先是葉爾羌河冬天水溫較低,魚類一般聚集成群生活,這一定程度上增加了線蟲與宿主接觸的機會。其次桿咽線蟲和對盲囊線蟲對魚類危害較小、即使感染強度很高也不會引起宿主死亡。長身高原鰍在每年3-7月為繁殖期,生理抵抗能力下降,攝食量也會有所增加;且隨著體長增加食量也會增加,這些均增加了線蟲感染量;年齡較大的長身高原鰍獲得更多再次感染線蟲機會而使體內累積線蟲數量增加。
桿咽線蟲在不同體長宿主組的感染強度的差異顯著(<0.05),這表明宿主年齡的大小對桿咽線蟲的感染影響較大,主要是隨著宿主年齡的增長,體型增大,食量也會增加有關,這與楊廷寶等的觀點一致。魯道夫對盲囊線蟲在不同體長組的感染強度的差異不顯著(>0.05),這表明宿主的年齡大小對魯道夫線蟲的感染影響較小,這可能由于對盲囊線蟲第一中間宿主以及終末宿主數量較少,導致宿主隨機感染,從而引起長身高原鰍體內聚集對盲囊線蟲程度較輕。
3.3 桿咽線蟲和魯道夫對盲囊線蟲的分布模式
本研究中、等均表明桿咽線蟲和魯道夫對盲囊線蟲在長身高原鰍各體長組中均為聚集分布,這與對盲囊線蟲在青海湖裸鯉和額爾齊斯河的銀鯽中的分布相似。聚集分布是一般寄生蟲在宿主種群的分布模式,即大多數宿主感染少量的寄生蟲,少數宿主感染絕大多數的寄生蟲,使寄生蟲對宿主種群的負面影響降到最小,這是寄生蟲與宿主長期協同進化的結果。據相關報道,聚集分布主要受宿主對寄生蟲的易感性、宿主行為的分化、寄生蟲直接在宿主體內繁殖等因素的影響。
