肌肽聯合超高壓對黑魚肉脂質氧化及肌原纖維蛋白的影響
2022-03-06 07:07:00孫良格徐佳美呂經秀張新笑徐為民李鵬鵬王道營
食品工業科技 2022年4期
孫良格,徐佳美,2,呂經秀,2,張新笑,徐為民,2,李鵬鵬,2, ,王道營,2
(1.江蘇省農業科學院農產品加工所,江蘇南京 210014;2.江蘇大學食品與生物工程學院, 江蘇鎮江212013)
黑魚(Ophiocephalus argusCantor)是我國常見的淡水經濟魚類,黑魚肉營養價值豐富,富含人體所需的鈣、鐵等多種微量元素,具有很好的藥用價值,可促進傷口愈合[1]。隨著人們生活水平的提高,特別是隨著我國生鮮超市的發展、銷售冷鏈的形成,消費者對冷鮮魚肉的需求日益增大,并對其鮮度、安全品質等的要求也越來越高。黑魚肉富含不飽和脂肪酸,但在冷藏期間極易發生氧化。脂肪氧化不僅會導致肉品風味改變、褪色,也是淡水魚腥味產生的主要原因[2]。此外,脂質氧化產物也可誘導蛋白質氧化反應,増加蛋白質間的交聯,從而導致肉的持水力下降[3]。因此,控制冷藏期間黑魚肉的脂質氧化對提高冷鮮魚肉品質具有重要意義。
肌肽是一種由β-丙氨酸和L-組氨酸兩種氨基酸組成的二肽,天然存在于動物肌肉中。肌肽通過螯合金屬離子、清除自由基和供氫、供電子能力等方式來抑制氧化[4]。邢子鑫等[5]研究發現肌肽將肉的冷藏期由9 d延長至15 d,對脂類氧化抑制效果顯著。超高壓技術是將密封于彈性容器內的食品置于以水或其他液體作為傳壓介質的壓力系統中,經100 MPa以上壓力處理,在常溫下達到殺菌、鈍酶改善食品品質等目的[6]。食品加工中超高壓技術主要應用于果蔬、果汁及食品包裝材料的研究,近年來開始被用于肉品品質提高的研究。肉制品中的內源酶在超高壓作用下構象改變,酶活性降低甚至失活[7]。用超高壓處理冷藏肉類產品時可以延緩肉品冷藏期間的脂肪氧化速率,保持冷藏肉類產品的風味等[8]。Bajovic等[9]研究表明,超高壓處理冷藏牛肉制品時加入天然抗氧化劑能夠延緩脂肪氧化。然而,肌肽聯合超高壓處理對黑魚肉的冷藏品質的研究尚未有報道。
本研究采用肌肽聯合超高壓處理分析其對黑魚肉冷藏期間的脂質氧化、蛋白氧化的影響,并進一步采用掃描電鏡、SDS-PAGE和圓二色譜等研究超高壓聯合肌肽預處理對黑魚肉肌原纖維蛋白的影響,以明確冷藏期間黑魚肉品質變化的機理,為黑魚魚肉的保鮮和冷藏提供理論依據。
1 材料與方法
1.1 材料與儀器
新鮮黑魚 質量為400~600 g,南京下馬坊華潤蘇果;肌肽 上海源葉生物公司;亞油酸 Sigma公司;戊二醛、硫代巴比妥酸、吐溫20、β-巰基乙醇、三氯乙酸、檸檬酸、磷酸氫二鉀、磷酸二氫鉀(均為分析純)、BCA蛋白濃度試劑盒、考馬斯亮藍蛋白濃度試劑盒、羰基含量試劑盒 南京建成有限公司;熒光蛋白酶分析試劑盒 G-Biosciences。
超高壓食品處理裝置 包頭科發高壓科技有限責任公司;Biofugestratos高速離心機 德國Heraeus公司;T25高速勻漿機 德國IKA公司;HI-9025酸度計 意大利Hanna;Mini-PROTEANTetra Cell電泳儀 美國Bio-Rad公司;BioTekSynergy2多功能酶標儀 美國BioTek公司;UniCenMR臺式冷凍離心機 英國Herolab公司。
1.2 實驗方法
1.2.1 原料處理 將新鮮黑魚宰殺、去頭、去內臟,取背部肌肉洗凈切碎,分成4組:對照組(Control)、肌肽組(CAR)、超高壓組(UHP)、超高壓+肌肽組(UHP+CAR)。每組20 g魚肉分裝于真空袋中,對照組和超高壓組分別加入8 mL PBS緩沖液后真空包裝,肌肽組和超高壓+肌肽組分別加入8 mL肌肽溶液(PBS緩沖液配制)后真空包裝,超高壓+肌肽組和超高壓組再進行超高壓(壓力300 MPa,保壓時間900 s)處理。各組處理后放4 ℃冷藏,每隔3 d隨機取樣測定丙二醛含量、羰基含量和TCA-溶解肽含量,同時進行SDS-PAGE、SEM和CD分析。
1.2.2 肌肽濃度的選擇 將濃度為0、10、15、20、25、30 mmol/L[5]的肌肽溶液分別加入裝好魚肉的真空袋中放置4 ℃冷藏,3 d后測定丙二醛含量,確定最優濃度。
1.2.3 丙二醛含量的測定 參照國標GB5009.181-2016《食品安全國家標準食品中丙二醛的測定》。稱取5 g樣品(精確至0.01g)放入100 mL帶塞錐形瓶中,準確加入50 mL三氯乙酸混合物,充分搖勻,加塞密封,置于恒溫振蕩器上,50 ℃振搖30 min,取出,冷卻至室溫,用雙層定量慢濾紙過濾,棄去初濾液,繼續濾液備用。分別準確吸取上述濾液和標準系列溶液各5 mL至25 mL帶塞比色管中,另取5 mL三氯乙酸混合物作為樣品空白,加入5 mL硫代巴比妥酸(TBA)水溶液,置于90 ℃水浴中反應30 min,取出,冷卻至室溫。用樣品空白調零點,在1 cm光程532 nm處測量樣品溶液和標準系列溶液的吸光度值,以標準系列溶液的質量濃度為橫坐標,繪制標準曲線。吸光度值作為縱坐標。
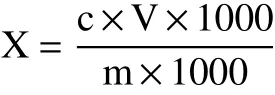
式中,X為樣品中丙二醛的含量,mg/kg;c為標準系列曲線得到的樣品溶液中丙二醛的濃度,μg/mL;V為試樣溶液的定容體積,mL;m為最終試樣溶液所代表的試樣質量,g;1000為換算系數;計算結果表示為在重復性條件下獲得的兩個獨立測定結果,結果保留兩位有效數字。
1.2.4 脂肪氧合酶及總蛋白酶活的測定 魚肉內源蛋白粗酶提取參照Gate等[10]方法,脂肪氧合酶活測定:在離心管中稱取2 g樣品,加入20 mL 0.05 mol/L pH7.0磷酸鹽緩沖液(含1 mmol/Lβ-巰基乙醇和2 mmol/L EDTA),在冰中使用高速勻漿器12000×g水浴勻漿4×10 s,10000×g離心15 min,取上清,四層紗布過濾,得酶粗提液,分裝0.1 mL小瓶,保存在-20 ℃。
底物溶液制備:將140 mg(0.05 mmol)亞油酸溶于5 mL(含180 μL吐溫20)脫氧重蒸餾水中調勻;滴加1 mol/L氫氧化鈉充分混勻至體系為澄清透明液體,直至亞油酸完全溶解,最后用脫氧重蒸餾水稀釋至50 mL,分裝于-20 ℃保存。
將0.1 mL酶原液、0.2 mL 亞油酸底物溶液和2.9 mL 0.05 mol/L 檸檬酸鹽緩沖液(pH5.5)在234 nm穩定混合1 min,讀取增加的吸光度值。脂肪氧化酶的相對活性單位(U)表示為1 min內酶活性單位增加0.001。空白為0.2 mL亞油酸底物溶液與2.9 mL檸檬酸緩沖液混合作為空白。
總蛋白酶活測定:內源蛋白粗酶用G-Biosciences熒光蛋白酶分析試劑盒檢測酶活。
1.2.5 羰基含量的測定 采用南京建成羰基試劑盒檢測樣品。準確稱取0.1 g肉樣加入0.9 mL的試劑一,冰水浴下機械勻漿,2500 r/min,離心10 min,取上清450 μL,加入50 μL試劑二,混勻,室溫放置10 min后,11000 r/min離心10 min,取上清進行蛋白羰基的測定,同時測定部分上清用雙縮脲法測定勻漿的總蛋白濃度。上清液分別取0.1 mL加入測定管和對照管,測定管加入0.4 mL試劑三,對照管加入0.4 mL試劑四,分別渦旋1 min,37 ℃準確避光反應30 min。反應后同時加入0.5 mL試劑五,渦旋1 min,離心(4 ℃,12000 r/min,10 min)去上清取沉淀。兩管分別加入無水乙酸乙酯混合應用液1 mL,離心(4 ℃,12000 r/min,10 min),去上清,留沉淀。兩管此過程重復3次后,加入1.25 mL試劑六,混勻后,37 ℃準確水浴15 min。渦旋混勻后。將全部沉淀溶解,以12000 r/min離心15 min,取上清液在370 nm處(紫外),0.5 cm光徑石英比色皿,試劑六調零,測定各管OD值。

1.2.6 總三氯乙酸(total trichloroacetic acid,TCA)-溶解肽含量的測定 將樣品去除肌膜、結締組織后切碎,稱量2.00 g肉糜,與18 mL預冷的5%三氯乙酸溶液進行混合,冰水浴10000 r/min勻漿40 s,在12000 r/min離心5 min后取上清液,采用BCA法測定小肽含量。
1.2.7 十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(SDSPAGE)分析 根據Shi等[11]方法首先提取肌原纖維蛋白后,再進行SDS-PAGE測定。取2.0 g碎肉組織,添加4倍(w/v)體積的MP分離緩沖液A(0.1 mol/L NaCl,2.0 mmol/L MgCl2,1.0 mmol/L EGTA,6.1 mmol/L Na2HPO4,3.9 mmol/L NaH2PO4·H2O,pH7.0,4 ℃)冰浴勻漿60 s(10000 r/min,30 s/次,2次),冷凍離心15 min(2000×g,4 ℃)后取沉淀,重復該步驟2次。將沉淀與8倍體積緩沖液B(0.1 mol/L NaCl,1.0 mmol/L NaN3,pH6.0)混合,冰浴10000 r/min勻漿60 s后,離心后(同上)取沉淀,該步驟重復2次后過4層紗布,離心得到提純的MP沉淀。沉淀溶于適量PB緩沖液(0.6 mol/L KCl,10 mmol/L K2HPO4,pH6.0)中,待用。
SDS-PAGE測定:100 μL MP溶液(調節蛋白濃度為1.0 mg/mL)中加入25 μL的上樣緩沖液后于95 ℃氣浴5 min滅酶,離心(12000×g,5min)后取上清用于電泳測定(12%分離膠與5%濃縮膠)。每孔等體積上樣5 μL,120 V恒壓約50 min。采用寬分子量蛋白(20~250 kDa,Thermo Scientific?)作為Marker參照。考馬斯亮藍染色液染色凝膠塊15 min,于搖床上均勻脫色,全自動凝膠成像分析儀對脫色凝膠塊成像分析。
1.2.8 掃描電鏡(SEM)分析 在魚肉表面下1 mm處切取的3 mm×3 mm×1 mm的小樣品塊,在含3%戊二醛固定1 h后,分別置于10%、40%、70%、100% 乙醇中梯度脫水并在室溫下揮干溶劑后噴金,最后使用SEM對樣品肌肉結構進行觀察。
1.2.9 圓二色譜(CD)分析 根據Zou等[12]方法測定肌原纖維蛋白二級結構變化。使用圓二色譜儀分析MP于185~250 nm處的光譜圖(25 °C)。測定前,采用PBS緩沖液(20 mmol/L,pH7.0)將MP濃度調整至200 μg/mL,并置于1.0 mm長的CD石英槽中進行光譜掃描(相應的緩沖液為參比)。掃描速率、反應時間和狹縫寬度分別為100 nm/min、0.50 s和2.0 nm。利用帶旋光儀的蛋白質二級結構估算程序計算相應百分率。
1.3 數據處理
所有實驗均重復3次,結果均以平均值±標準差表示。采用Microsoft Excel 2016軟件對數據進行處理分析,SPSS 22軟件進行顯著性分析(多重比較采用Duncan檢驗),并用Origin 9.1軟件作圖。
2 結果與分析
2.1 肌肽聯合超高壓處理對冷藏期黑魚肉脂質氧化的影響
黑魚魚肉富含不飽和脂質,極易發生氧化。脂質氧化產生的揮發性物質是魚肉冷藏期間腥味產生的主要原因[13]。動物宰后肌肉脂質氧化主要由活性氧和脂肪氧合酶引起。清除活性氧和抑制脂肪氧合酶活性對抑制冷藏期魚肉脂質氧化并提高魚肉品質具有重要意義。肌肽作為新型食品添加劑,具有清除活性氧、抑制氧化的作用。如圖1所示,肌肽能顯著抑制冷藏魚肉的脂質氧化(P<0.05),肌肽濃度在25 mmol/L時,對魚肉脂質氧化抑制作用最強,所以選定25 mmol/L肌肽溶液完成后續儲藏實驗。

圖1 添加不同濃度肌肽溶液的魚肉4 ℃冷藏3 d后MDA含量Fig.1 MDA content of snakehead fish with different concentrations of carnosine solution added after 3 days of refrigerated storage at 4 ℃
超高壓技術是一種非熱加工鈍酶技術,在此條件下酶分子結構中的氫鍵、疏水離子作用等發生變化,最終導致酶的分子構象改變而失活。Gavan等[14]研究發現,隨著壓力和保壓時間的增加,超高壓鈍酶效果增強。王幫國等[15]研究發現,隨著壓力和保壓時間增加,脂肪氧合酶分子結構發生顯著變化,酶活不斷降低。如圖2A所示,超高壓(壓力300 MPa,保壓時間900 s)作用下脂肪氧合酶的酶活降低了64.46%。
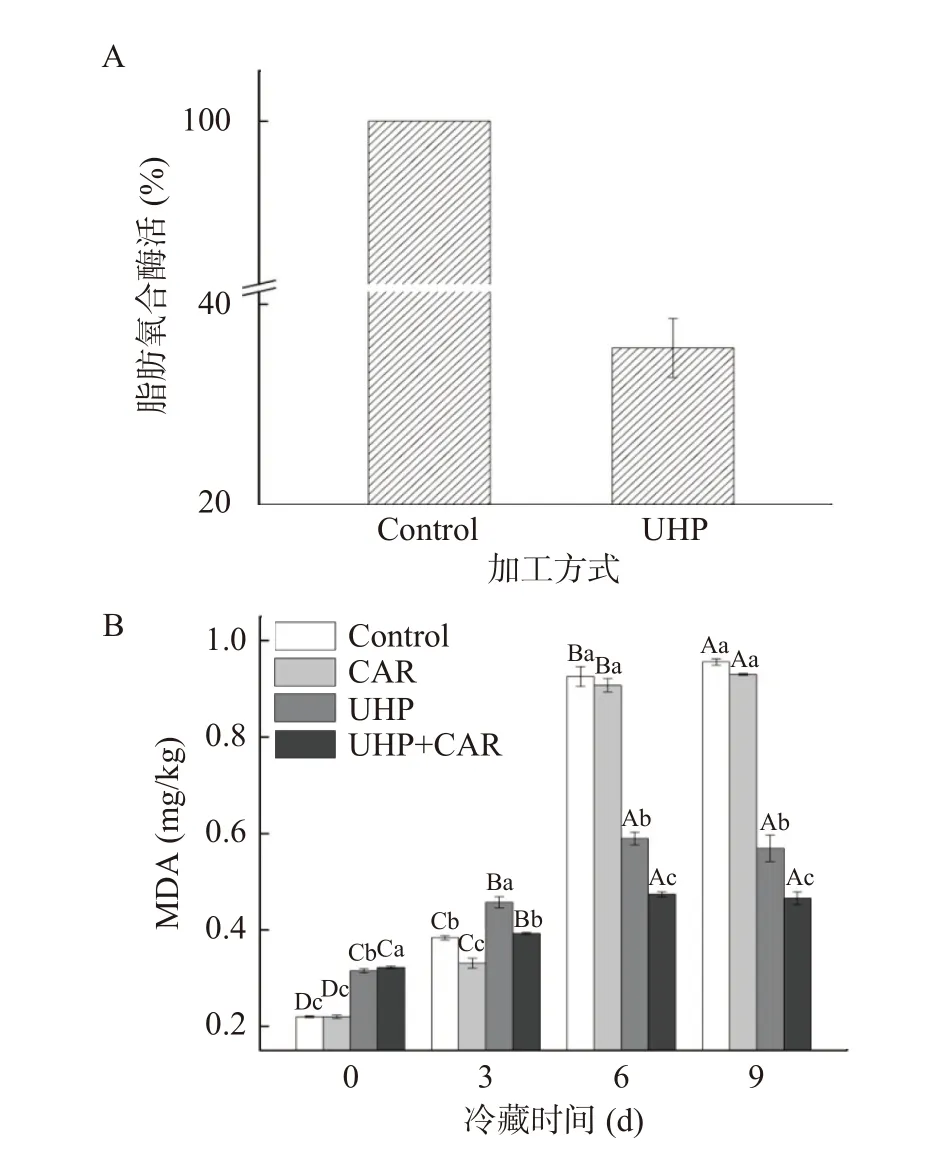
圖2 魚肉超高壓對脂肪氧合酶活影響及肌肽聯合超高壓處理對冷藏期黑魚脂質氧化的影響Fig.2 Effects of lipoxygenase activity after UHP treatment and lipid oxidation with carnosine combined with ultrahigh pressure treatment of snackhead during cold storage
MDA為脂質氧化過程產生的丙二醛,用于檢測魚肉在冷藏期間發生脂質氧化程度。如圖2B所示,對照組MDA值隨著冷藏時間的延長不斷升高,到第9 d的MDA值由初始的0.22 mg/kg上升到0.956 mg/kg。CAR組MDA值在冷藏期間一直低于對照組,在第3 d時顯著低于對照組(P<0.05)。肌肽通過清除活性氧、螯合金屬離子來抑制脂質氧化。Gopalakrishnan等[16]通過添加不同濃度的肌肽于豬肉中,也發現肌肽能顯著抑制豬肉的脂質氧化。在冷藏后期CAR組值顯著增加(P<0.05),可能是微生物的大量繁殖加速了脂質氧化[17]。與對照組和CAR組相比,UHP和UHP+CAR兩組冷藏期間MDA值上升速率緩慢,到第9 d分別比對照組MDA值低0.387、0.490 mg/kg。超高壓鈍化脂肪氧合酶活性,減少了冷藏期間魚肉的脂質氧化。此外,超高壓處理還能抑制微生物的活性,在一定程度上減緩了微生物活動對脂質氧化的作用[17]。在冷藏第9 d,UHP+CAR組較UHP組低0.103 mg/kg,說明肌肽聯合超高壓能協同抑制冷藏魚肉的脂質氧化。Marcos等[18]的研究表明,抗氧化劑聯合超高壓處理肉制品,比只添加抗氧化劑的肉制品保鮮效果好,該結論與本文結果一致。經過超高壓處理的兩組樣品的MDA值高于另外兩組,可能是由于高壓促使部分復合物釋放出游離金屬離子,導致部分脂質發生氧化[19]。
2.2 肌肽聯合超高壓處理對黑魚肉冷藏期間蛋白質氧化的影響
蛋白氧化影響肉的滋味、嫩度、色澤、多汁性等感官品質,還影響肌肉的組織結構和功能特性。朱文慧等[20]研究發現魷魚蛋白氧化過后肌纖維間隙增大、結構疏松,肌原纖維小片化指數增加,保水性降低。動物宰后肌肉蛋白質的氧化多由活性氧直接引發或由脂質氧化產物誘導引起的蛋白質肽鏈骨架重置和氨基酸側鏈共價修飾間接引發[21],其中氨基酸側鏈被氧化形成的羰基常用于檢測蛋白質氧化的程度。如圖3所示,各組樣品的羰基含量在冷藏期間均呈上升趨勢。CAR組羰基含量一直低于對照組,在冷藏期間第3 d CAR組羰基抑制率為22.08%,達到最高。UHP+CAR組在冷藏期第3 d羰基含量較UHP組減少10.85%,差異顯著。以上說明肌肽在冷藏期前3 d發揮抑制蛋白氧化作用最強。肌肽可以清除活性氧減少氨基酸側鏈的氧化,也可以與活性醛反應,減少蛋白質氧化的誘導物,抑制冷藏期間的脂質氧化,從而降低蛋白質的羰基化。此外,肌肽還能結合蛋白質前體上的羰基,形成蛋白質-羰基-肌肽復合物,防止蛋白形成損傷性交聯[22]。UHP組和UHP+CAR組初始羰基含量高于對照組與CAR組,可能是超高壓在處理魚肉過程中釋放的自由基引起的蛋白氧化。UHP組和UHP+CAR組的羰基值在冷藏期間增速較緩,UHP+CAR組第9 d羰基含量較對照組減少21.70%。結合2.1中結果,說明抑制脂質氧化能減少蛋白質氧化的誘導劑,有利于抑制蛋白質的氧化。此外,有研究表明肉品貯藏過程中微生物增長能促進肌肉蛋白氧化,超高壓處理能在一定程度上滅活腐敗微生物[6],也有助于抑制蛋白質的氧化。
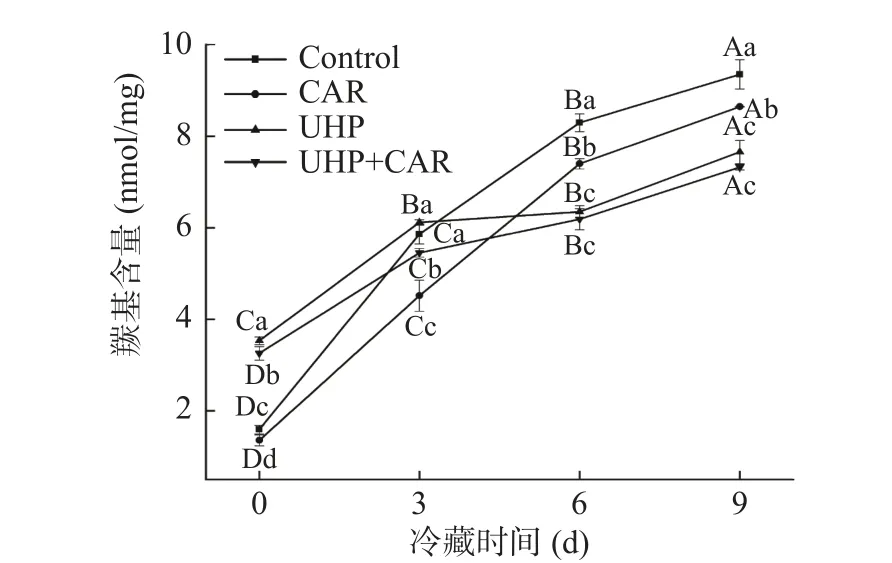
圖3 肌肽聯合超高壓處理對冷藏期黑魚羰基含量的影響Fig.3 Effects of CUH treatment on carbonyl content of snakehead fish during refrigerated storage
2.3 肌肽聯合超高壓處理對黑魚肉冷藏期間蛋白質降解的影響
肌原纖維蛋白包括肌球蛋白、肌動蛋白、原肌球蛋白、肌原蛋白以及多種微量調節蛋白,其中肌球蛋白重鏈(MHC)和肌動蛋白(Actin)是最主要的蛋白條帶。如圖4所示,超高壓處理之后,MHC和Actin的條帶強度明顯減弱。馬海建[23]在對草魚魚糜中的肌動球蛋白進行超高壓處理后,發現MHC帶和Actin帶變化不大。UHP和UHP+CAR組在超高壓處理后,35 kDa附近(肌鈣蛋白T、原肌球蛋白)的條帶變淺/消失。結合前文結果及文獻報道,推測超高壓處理后魚肉部分蛋白發生了氧化和變性,導致部分蛋白成為不可溶蛋白[24]。冷藏后期,對照組與CAR組樣品在9 d時15 kDa附近出現了新的條帶,可能是在內源蛋白酶和微生物分泌的蛋白酶作用下,肌原纖維蛋白發生降解。UHP和UHP+CAR組較對照組和CAR組的肌原纖維蛋白在冷藏期未出現明顯變化,可能由于超高壓作用抑制了蛋白酶活性。葛黎紅[25]研究表明,內源蛋白酶和微生物是造成魚肉蛋白降解和質構劣化的主要因素,而超高壓作用既能降低內源蛋白酶活,又可以有效滅活微生物,從而減少肌原纖維蛋白的降解。

圖4 冷藏期間魚肉肌原纖維蛋白SDS-PAGE的變化Fig.4 Changes of SDS-PAGE of myofibrillary protein in snakehead fish during refrigerated storage
TCA-溶解肽是常用的蛋白水解參數,能體現SDS-PAGE難以分析的低分子肽的變化。如圖5A所示,對照組及CAR組的溶解肽含量隨著冷藏時間的延長逐漸升高,9 d后CAR組溶解肽含量減少了12.20%。UHP和UHP+CAR組第0 d小肽含量高于對照組,可能是超高壓作用使魚肉部分蛋白質發生降解。UHP+CAR組冷藏期間溶解肽含量的變化平穩且保持在較低水平,9 d后含量為0.15 mg/mL,分別比對照組、CAR組、UHP組低了63.41%、53.33%、6.25%。組織蛋白酶B、H、L等魚肉冷藏期間最活躍的溶酶體蛋白酶,對肌肉組織降解具有重要作用[26]。微生物分泌的蛋白酶也能夠加速肌肉蛋白的水解作用。由圖5B所示,高壓過后的總蛋白酶活降低了25.75%,所以9 d后UHP+CAR組能夠保持較低水平的溶解肽含量。此外,脂質氧化生成的自由基、某些羰基化合物具有很高的反應活性,能夠促進蛋白質降解[27]。馬海建[23]也研究發現在超高壓300 MPa及以上壓力條件下,組織蛋白酶B活性顯著下降,同時微生物活性降低。肌肽能延緩自由基及羰基化合物的生成,抑制蛋白氧化引起的蛋白質降解。因此超高壓和肌肽能協同作用抑制冷藏魚肉的蛋白質降解。
圖5 肌肽聯合超高壓處理對冷藏期黑魚TCA-溶解肽的影響及黑魚肉超高壓后總蛋白酶活情況Fig.5 Effects of TCA-soluble peptides with carnosine combined with ultra-high pressure treatment and the total protease activity after UHP treatment of fish meat during cold storage
2.4 肌肽聯合超高壓處理對冷藏期黑魚肉及肌原纖維蛋白結構的影響
魚體肌肉組織中內源蛋白酶活性較強,淡水魚在冷藏過程中極易腐敗變質。如圖6所示,對照組及CAR組的肌肉組織線條紋理緊湊清晰,排列有序,而經超高壓處理的UHP和UHP+CAR組樣品表面紋理變得模糊,并出現少量的小分子顆粒,可能是高壓的作用使肌漿蛋白變性凝聚成小分子[28]。冷藏6 d時,對照組纖維表面出現空洞及蛋白網絡松散坍塌,纖維間隙增大,肌內膜與肌纖維分離。冷藏期內源蛋白酶及微生物的作用下蛋白被分解利用,造成質構劣化。CAR組較對照組的表面結構損壞較小,纖維表面出現少量空洞。肌肽能夠抑制魚肉在冷藏期發生的脂質、蛋白氧化及蛋白分解,但冷藏后期微生物大量繁殖分泌蛋白酶加速了蛋白降解。UHP組在冷藏期間肌束排列有序,肌纖維表面結構空隙較少。UHP+CAR組表面肌束清晰,纖維表面未出現明顯空洞及蛋白網絡松散狀態,可能是由于超高壓抑制魚肉中總蛋白酶活和微生物活性。如圖6A所示,在超高壓作用下總蛋白酶活降低25.75%,減少了魚肉中肌原纖維蛋白的降解。上文又驗證了肌肽在魚肉冷藏期間顯著抑制脂質氧化、蛋白氧化和微生物分解蛋白速率,有利于延緩魚肉在冷藏期間腐敗速率。

圖6 不同處理下組織超微結構電鏡圖Fig.6 Ultrastructural electron microscopy of tissue under different treatments
為進一步研究肌肽聯合超高壓對冷藏魚肉蛋白質品質變化的影響,本研究通過圓二色譜分析了肌肽聯合超高壓對黑魚肌肉蛋白質二級結構的影響。220 nm和209 nm兩個負峰是α-螺旋結構的特征吸收峰,216 nm是β-折疊的吸收峰,220~230 nm是β-轉角吸收峰,198 nm是無規則卷曲的吸收峰[29-30]。如圖7和圖8所示,對照組和CAR組蛋白0 d在209和220 nm附近兩個負峰明顯,冷藏期間負峰逐漸減少。對照組和CAR組蛋白α-螺旋比例在冷藏期間整體呈現先減少后增多的趨勢。在0 d對照組和CAR組MP結構以α-螺旋為主,分別占44.3%和45.6%。對照組和CAR組α-螺旋比例第3 d分別下降了73.36%、67.98%。蛋白質氧化可引起蛋白質的骨架和側鏈的改變,從而導致蛋白質的一級、二級和三級結構發生改變[31]。可能是由于冷藏期間微生物繁殖及脂質氧化加強,導致蛋白氧化,進而引起蛋白結構改變[32]。0與3 d CAR組α-螺旋比例較對照組分別提高2.93%和23.72%。對照組和CAR組α-螺旋比例在9 d出現明顯的上升,同時CAR組較對照組具有更低的無規則卷曲比例和更高的α-螺旋結構比例。說明肌肽在冷藏的前3 d能夠顯著抑制蛋白結構改變。該結論與上文肌肽在冷藏前期能夠顯著抑制蛋白氧化的結論相契合。肌肽通過清除活性氧、螯合金屬離子等方式減少MP結構損傷,有利于蛋白保持有序結構。
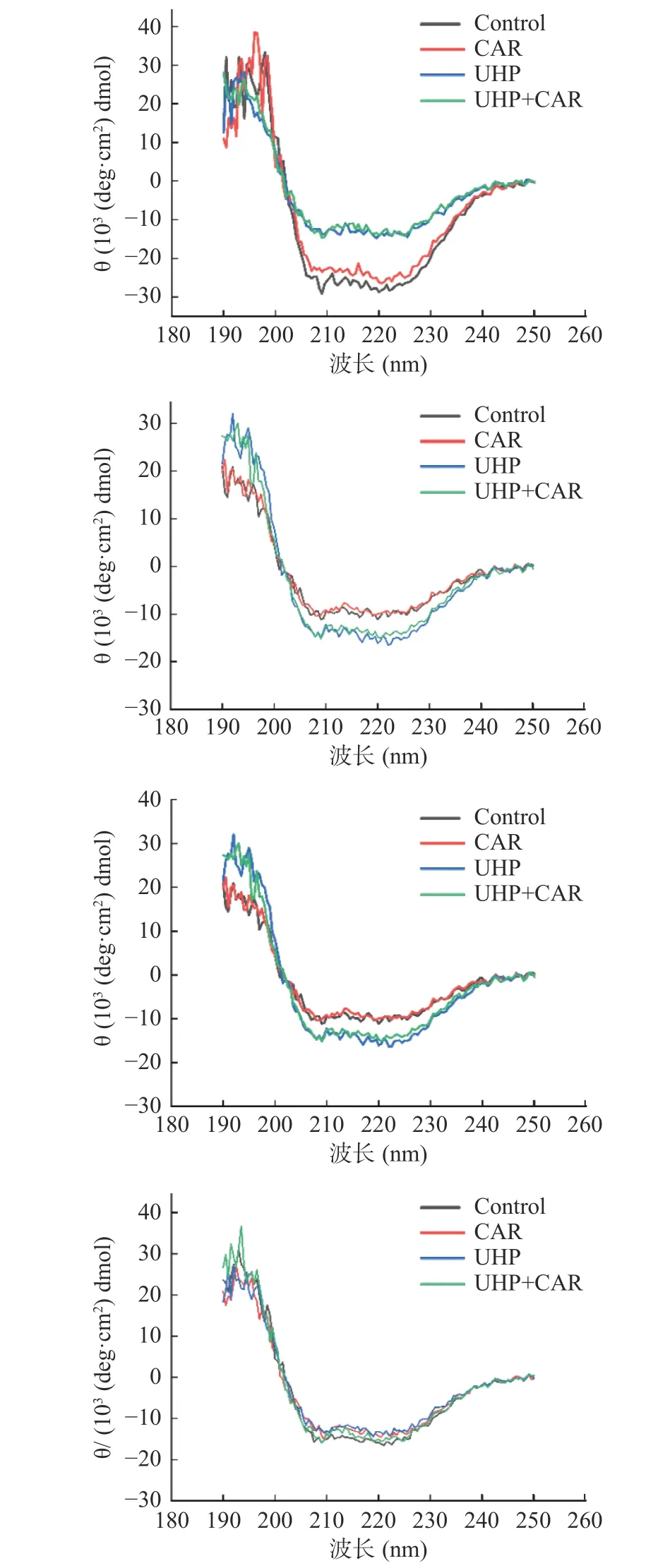
圖7 肌肽聯合超高壓處理對冷藏期肌原纖維蛋白結構的影響Fig.7 Effect of CUH treatment on MP structure during refrigerated storage
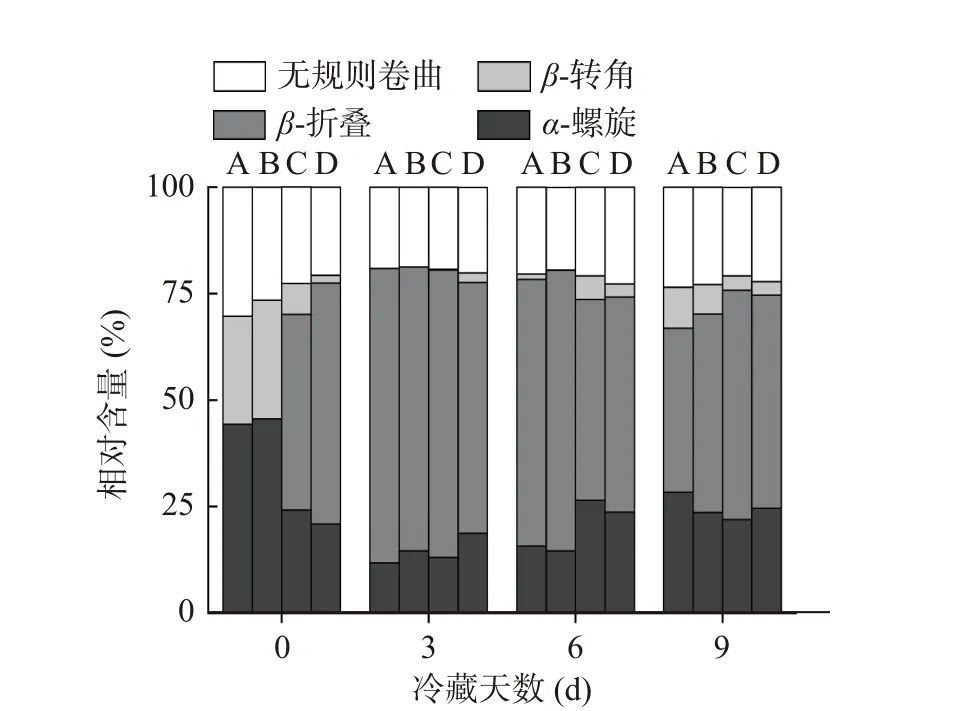
圖8 肌肽聯合超高壓處理對冷藏期肌原纖維蛋白二級結構相對百分含量的影響Fig.8 Effects of CHU treatment on the relative percentage of secondary structure of MP during refrigerated storage
UHP和UHP+CAR組在0 d時220和209 nm兩個負峰較對照組變小。冷藏期間,UHP+CAR和UHP組220 nm和209 nm負峰比較穩定。UHP組在0 d時α-螺旋比例較對照組減少了45.37%,β-折疊比例升高為45.9%。王瑋等[33]發現超高壓處理降低了酶蛋白結構中α-螺旋的比例,與本結果相同。Tan等[34]認為高壓通過影響蛋白質分子間的相互作用鍵(氫鍵、疏水相互作用、靜電力),導致蛋白質變性。UHP+CAR組0 d時較CAR組的β-折疊上升了56.6%,無規則卷曲比例減少21.89%。UHP+CAR組0 d時β-折疊比例較UHP組上升了23.31%,無規卷曲比例減少8.81%。UHP+CAR組與CAR組、UHP組相比具有較高β-折疊和較低的無規則卷曲比例,初始肌原纖維蛋白處于更有序狀態。在冷藏期間UHP+CAR組的α-螺旋比例先減少后增多,第3 d UHP和UHP+CAR組α-螺旋比例下降率明顯低于對照組和CAR組。超高壓抑制內源蛋白酶和微生物活性降低了肉品的氧化,從而減弱了蛋白結構的變化。在第6 dα-螺旋比例出現明顯增高,較CAR組提前3 d。表明超高壓處理作用后魚肉蛋白結構展開,在冷藏第6 d后逐漸呈現有序化,超高壓處理使魚肉體系中形成新的氫鍵形成,蛋白質結構更加有序化。第9 d時的UHP+CAR組較對照組、CAR組和UHP組具有更高的α-螺旋比例和較低的無規則卷曲。超高壓抑制內源蛋白酶和微生物活性及肌肽抑制蛋白的氧化分解的作用下,蛋白的結構性破壞減少,UHP+CAR處理促進肌原纖維蛋白結構的有序化。
3 結論
研究表明,采用不同濃度的肌肽溶液處理魚肉,在肌肽溶液為25 mmol/L時,魚肉的抗脂質氧化能力最強。本論文采用肌肽聯合超高壓處理后的魚肉的脂肪氧合酶及總蛋白酶活顯著降低,脂質氧化程度及TCA-溶解肽含量受到顯著抑制(P<0.05)。冷藏期間魚肉肌原纖維蛋白的羰基含量顯著減少,蛋白氧化程度顯著降低(P<0.05)。SEM表明,肌肽聯合超高壓處理后的黑魚肉在冷藏期間魚肉纖維表面結構空隙較少,未出現蛋白網絡松散狀態。SDS-PAGE和TCA-溶解肽結果表明,超高壓不會引起肌原纖維蛋白大量降解,肌肽聯合超高壓處理顯著降低了魚肉冷藏期間肌原纖維蛋白的降解(P<0.05)。圓二色譜分析發現,肌肽聯合超高壓處理后黑魚肉的肌原纖維蛋白的二級結構發生變化,冷藏期間α-螺旋的比例先減少后增加,說明冷藏后期有新的氫鍵生成,蛋白結構呈現有序化。因此,肌肽聯合超高壓處理既能夠改善魚肉冷藏品質,又能延長魚肉制品冷藏貨架期,同時本文研究結果可為調控魚肉品質提供理論基礎。
