喀斯特區不同退化程度植被群落植物-凋落物-土壤-微生物生態化學計量特征
2022-03-07 09:07:06俞月鳳何鐵光曾成城宋同清彭晚霞韋彩會蘇利榮
生態學報 2022年3期
關鍵詞:植物
俞月鳳,何鐵光,*,曾成城,宋同清,彭晚霞,韋彩會,蘇利榮,張 野,范 適
1 廣西壯族自治區農業科學院農業資源與環境研究所, 南寧 530007 2 中國科學院亞熱帶農業生態研究所, 長沙 410125 3 湖南環境生物職業技術學院園林學院, 衡陽 421005
生態化學計量學理論認為有機體能夠通過控制自身的養分元素平衡,使其元素組成比值和外界環境的元素供應保持一種相對穩定的狀態[1],這種內穩性是生態化學計量學研究的前提[2]。有機體C、N、P 元素含量存在一個相對穩定的比值,其中任何一種元素的較大變化都將使這一比值發生變化;因此,通過有機體C∶N∶P化學計量特征的變化,該理論可以用來判斷有機體生長、發育或者繁殖的限制元素種類[3]。生態化學計量學是生態學研究的前沿領域之一,研究主要集中于生態過程中C、N、P元素的計量關系及其生態學意義。當前,國內外學者對陸地生態系統各組分(植物、凋落物、土壤和微生物)C∶N∶P生態化學計量學特征進行了大量研究,尤其在限制性養分元素判別方面形成了植物 N∶P 養分限制理論[4],但有關退化生態系統的生態化學計量特征及養分限制的研究較少,主要集中在退化草地、山地、濕地及竹林地等生態系統[5—8],且主要關注某一組分(土壤、植物、凋落物或者微生物)或土壤-植物-凋落物的生態化學計量學特征,將“植物-凋落物-土壤-微生物”連續體作為系統研究退化生態系統內部C、N、P 計量學及其相互關系的研究尚未見報道。在植物-土壤系統中,土壤養分循環與植物營養過程與微生物的分解作用密切關聯,微生物生物量C∶N∶P決定了其對養分的礦化、固存作用,進而影響著土壤肥力[9];土壤有機質及植物凋落物反過來也會對微生物的分解速率產生影響,從而導致土壤養分有效性的改變[10],而植物養分化學計量比的改變則可以反映出養分有效性的變化;養分有效性的變化則可以通過植物的改變反映出來[11]。此外,植物養分元素的內穩態可反映各養分元素的平衡狀態, 是植物對環境變化的響應策略[12]。植物生態化學計量內穩性研究已經得到學者們的關注,但有關研究還不多[13]。由于不同的生態系統類型、不同植物群落類型之間存在明顯差異,植物生態化學計量內穩性在不同生態系統和群落中的特征亟待闡明。因此,利用生態化學計量學的思路來研究退化生態系統中“植物-凋落物-土壤-微生物”系統C、N、P元素的動態平衡及植物內穩性特征具有重要的生態學意義。
喀斯特地區環境由于其形成的碳酸鹽巖基質的特殊性而與常態地貌有著極大的差別,過度的人為干擾造成了喀斯特地區植被的退化,水土流失加劇,導致了十分嚴重的石漠化現象,嚴重制約了該區域可持續發展。生態化學計量學的理論和方法可為研究喀斯特區域植被退化過程及其機理提供新思路。近年來,有關喀斯特地區生態化學計量學方面的研究發展較快。劉娜等[14]和劉立斌等[15]探究了喀斯特高原石漠化區次生林植物、凋落物和土壤C、N、P生態化學計量特征;Song等[16]報道了喀斯特地區土壤和微生物碳氮磷化學計量特征對不同演替階段植物群落的響應;胡培雷等[17]研究了喀斯特石漠化地區不同退耕年限下桂牧1號雜交象草植物-土壤-微生物生態化學計量特征。這些研究表明喀斯特不同植被類型群落生態化學計量比存在差異,或受N 或受P 素制約,但對群落生態化學計量特征與喀斯特植被退化的關系及退化機理方面的研究尚顯不足,研究喀斯特植被退化過程中植物、凋落物、土壤和微生物系統C、N、P生態化學計量特征及植物內穩性特征的變化,有助于理解喀斯特生態系統退化機理。本文以桂西北喀斯特地區為研究區域,通過建立不同退化程度植被群落樣地,并進行土壤和植被調查,研究植物-凋落物-土壤-微生物系統生態化學計量變化特征、相互關系及植物內穩性特征,旨在揭示:(1)喀斯特生態系統退化是否會引起植被、凋落物、土壤、微生物系統C、N、P含量變化,從而改變其化學計量特征及養分限制格局與內穩性?(2)這種變化與喀斯特生態系統退化的關系如何?通過上述問題的探討,以期為喀斯特退化生態系統植被恢復與重建提供科學依據。
1 研究區概況及研究方法
1.1 研究區概況
研究區選擇在廣西大化瑤族自治縣(107°18′45″—108°03′45″E,23°32′30″—24°22′30″N),位于云貴高原向廣西丘陵過渡地帶的斜坡上,屬于廣西最典型的“九山半水半分田”的石山區貧困縣。全縣總面積2716 km2,耕地總面積162 km2,境內峰叢密布,喀斯特面積共2059 km2,占全縣總面積的73%,喀斯特地貌較為典型。成土母質以碳酸鹽巖為主,地帶性紅壤只是分布在土山,石山、半石山普遍為石灰土。研究區氣候溫和,屬中亞熱帶季風氣候,雨熱同季。年平均氣溫18.2—21.3 ℃。1月份氣溫最低,平均12.2 ℃,極端最低氣溫零下3 ℃,7月份氣溫最高,平均28.2 ℃,極端最高氣溫39.7 ℃。年日照時數1217—1587 h,年積溫6300 ℃。生長期、無霜期年平均分別達300 d、335 d。多年平均相對濕度74%—80%。年平均降雨量1249—1673 mm,蒸發量1213—1645 mm。桂西北喀斯特地區是西南喀斯特地區代表性區域,其生態環境極其脆弱,該區域一直以來是喀斯特地區生態恢復與重建的重點和難點。自20世紀80年代中期之后,該區域開始實施以植被恢復為主的宏觀生態工程,植被開始得到自然恢復,因此不同退化程度的植被群落共存。
1.2 研究方法
1.2.1野外調查及取樣
本研究在對研究區環境及植被特征全面調查的基礎上,結合相關資料,采用空間序列代替時間序列的方法,將植被群落分為5個退化程度(表1):未退化(Ⅰ)、潛在退化(Ⅱ)、輕度退化(Ⅲ)、中度退化(Ⅳ)、重度退化(Ⅴ)。經過全面踏查,2016年7月選擇符合5個退化程度的典型植被群落設置樣地,每個退化程度設置9個20 m × 20 m樣地,共計樣地45個,樣地設置及樣地基本情況團隊已報道過[18—19]。在樣地內按“S”型隨機采取5—8個表層土壤樣品(0—15 cm),混合后代表該樣地表層土樣;在每個樣地內選擇各群落內的優勢種采集植物葉片。各喬木按徑階選取平均標準木采集葉片,同種樹種不同徑階標準木葉片混合,取冠層東南西北4 個方位和上中下各部位完整的成熟葉片,每份樣品200—300 g;灌叢和灌草叢中灌木葉片的采集在地面借助剪刀完成,收集方法同上。采集草本植物葉片時直接用剪刀剪取成熟葉片。在樣方的4個角和中央設置5個1 m×1 m的方格收集凋落物,其中部分為枯落物,如凋落葉片、枝條、草本和木炭等,有些是難以分辨的有機物分解碎片。
1.2.2樣品處理及測定
每個土壤樣品充分混勻后按四分法分成兩份,一份現場密封后入冰柜保存(4 ℃),并盡快分析土壤微生物生物量C、N、P;一份帶回實驗室風干,先過1 mm篩,得到土壤樣品再過0.149 mm篩,用于測定土壤有機質、全氮、全磷。收集到的葉片和凋落物帶回實驗室后,放入烘箱105℃ 殺青2 h后80℃烘干至恒量,然后粉碎、過0.149 mm篩、裝信封待測定。有機碳(SOC)含量采用重鉻酸鉀氧化-外加熱法測定,全氮(TN)含量采用半微量凱氏法-流動注射儀法測定,全磷(TP)含量采用NaOH熔融-鉬銻抗顯色-紫外分光光度法測定。植物全C含量采用重鉻酸鉀氧化-外加熱法測定,N含量采用H2SO4-H2O2消煮、流動注射儀法測定,P含量采用H2SO4-H2O2消煮、鉬銻抗比色法測定。土壤微生物量碳采用CHCl3熏蒸法-K2SO4提取-總有機碳自動分析儀法,微生物量氮采用CHCl3熏蒸-K2SO4提取-氮自動分析儀法,微生物量磷采用CHCl3熏-NaHCO3提取-Pi 測定-外加Pi 校正法-紫外分光光度法測定;微生物碳、氮轉換系數為0.45,微生物磷轉換系數為0.40[20]。
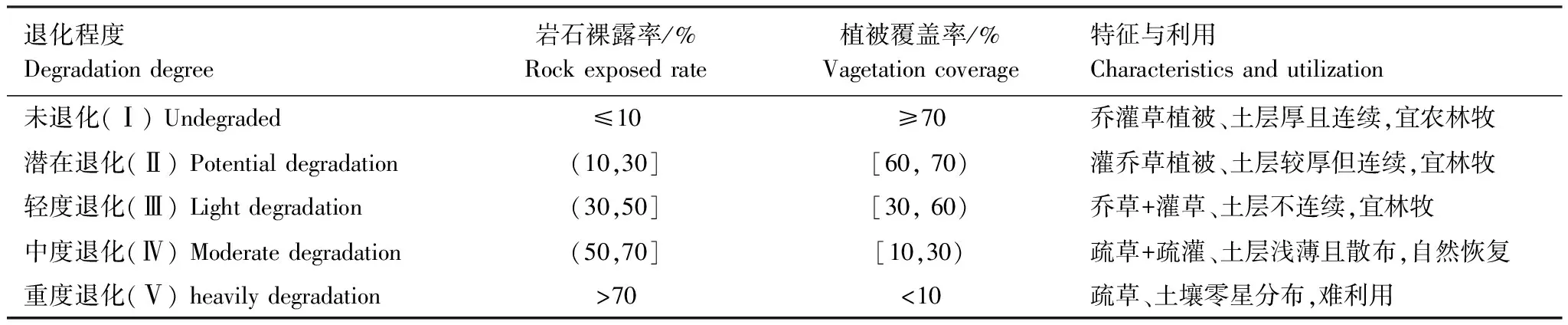
表1 植被群落退化等級劃分標準
1.3 數據處理
化學計量比根據各優勢種在群落內的重要值,利用加權平均計算。采用Excel 2010 和SPSS 18. 0 軟件對數據進行統計分析。用單因素方差分析(ANOVA) 檢驗不同退化階段土壤、植物、凋落物和微生物C、N、P 濃度及其化學計量比之間的差異性,然后用最小顯著差數法(LSD) 檢驗差異顯著性。
植物內穩態反映植物隨著環境變化保持自身化學組成穩定的能力[21],內穩性的高低用內穩性指數來表示,通過內穩性模型計算得出,該模型公式為:
y=cx1/H
其中,x為土壤N或P含量(g/kg)或N∶P值,y為植物體器官對應的全氮(N)或全磷(P)含量(g/kg)或N∶P值,c為常數,H為植物內穩性指數。
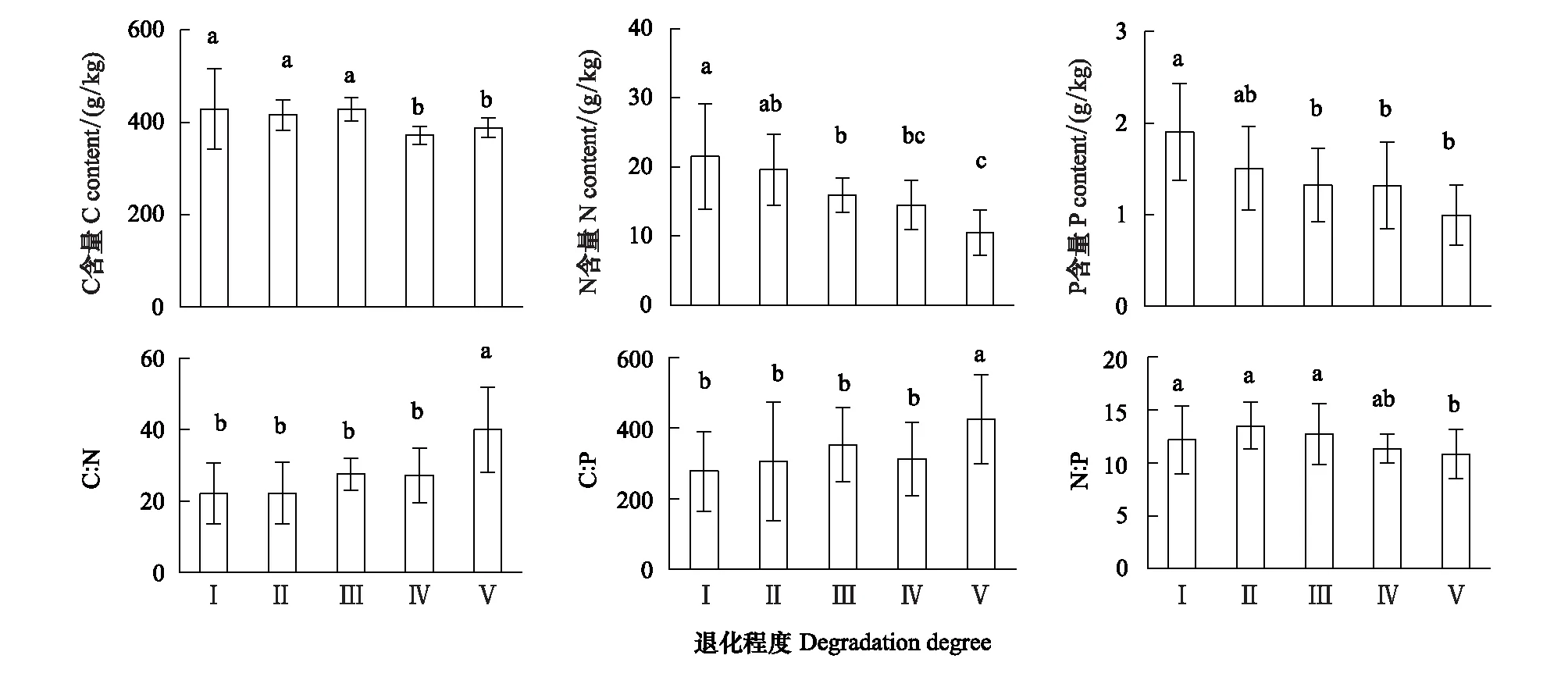
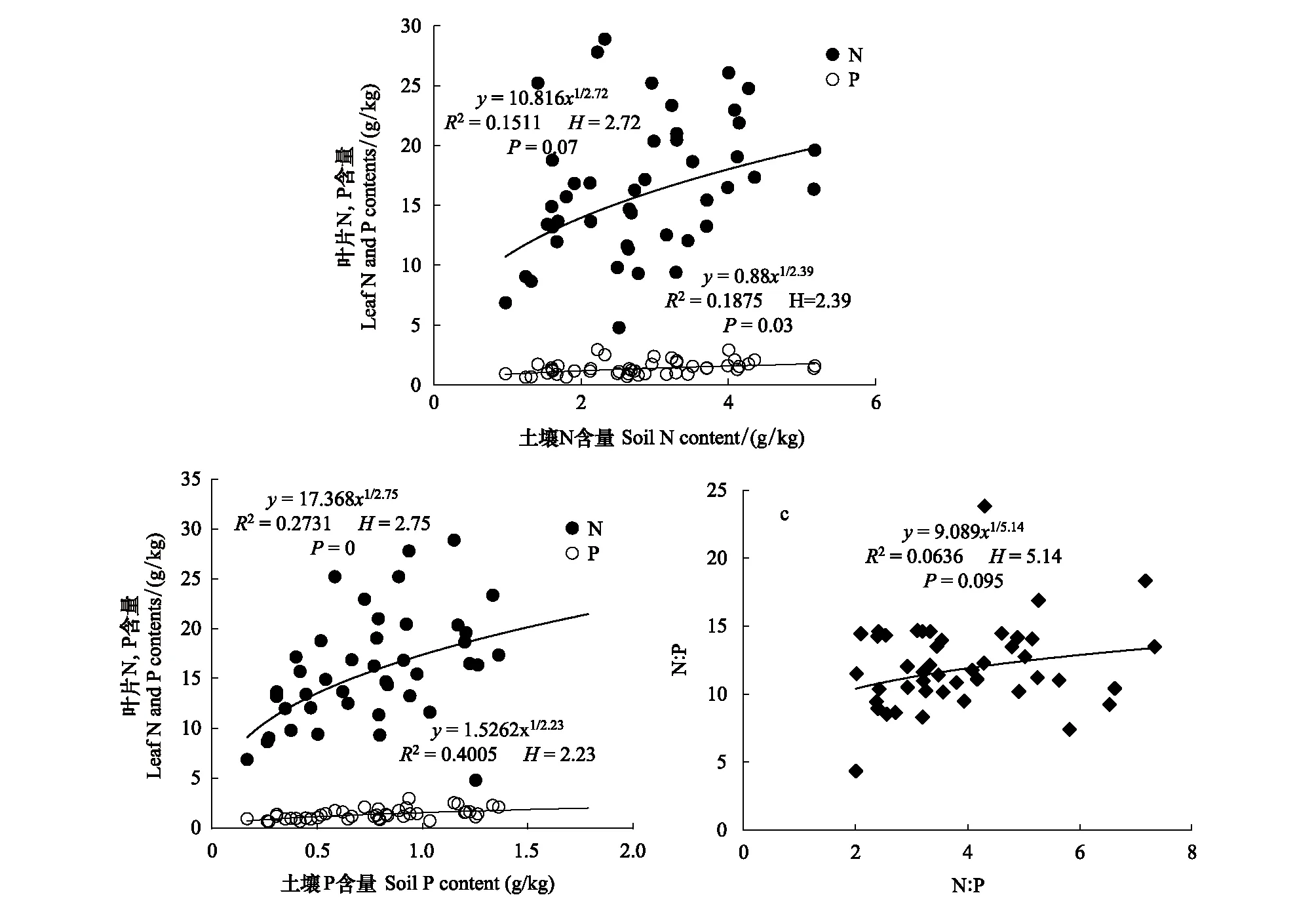
Persson[22]認為植物內穩性特征可用內穩性指數H進行界定。回歸分析顯著性檢驗采用α=0.1 進行單側檢驗,若回歸關系不顯著 (P>0.1),被定義為“絕對穩態”;當回歸分析顯著時(P≤0.1),按內穩性指數可將植物劃分為4種類型:H>4為穩態型;2 由圖1 可知,隨退化程度加劇,葉片C、N、P含量呈降低變化趨勢,葉片C∶N和C∶P逐漸升高,N∶P則呈先升高后降低的趨勢。未退化、潛在退化與輕度退化群落葉片的C含量及中度退化、重度退化群落葉片的C含量間均無顯著差異,但前三者葉片C含量均顯著高于后二者。未退化群落葉片N含量最高,與潛在退化群落無顯著差異;潛在、輕度與中度退化群落葉片N含量間無顯著差異;重度退化群落葉片N含量最低,與中度退化群落無顯著差異,但顯著低于輕度退化群落。未退化群落葉片P含量顯著高于輕度、中度和重度退化群落,潛在退化群落葉片P含量則與其他群落間均無顯著性差異。重度退化群落葉片C∶N和C∶P顯著高于其他群落,其N∶P則最低;其他四個群落葉片的C∶N、C∶P和N∶P之間無顯著性差異。 圖1 不同退化程度群落植物葉片C、N、P 含量及化學計量比Fig.1 Leaf C、N、P contents and stoichiometric ratio in vegetation communities along degradation不同小寫字母代表不同退化程度之間差異顯著(P<0.05);Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ分別表示未退化、潛在退化、輕度退化、中度退化、重度退化 圖2 喀斯特不同退化程度群落凋落物C、N、P 含量及化學計量比Fig.2 Litter C、N、P contents and stoichiometric ratio in vegetation communities along degradation in karst area 隨著退化程度的加劇,凋落物C含量呈先降低后升高再降低的趨勢,N、P含量表現為先升高后降低的趨勢(圖2)。中度退化群落凋落物C含量顯著高于未退化、潛在退化及重度退化群落,且未退化、潛在退化及重度退化群落凋落物C含量并無顯著差異,但均顯著高于輕度退化群落。未退化、潛在退化和輕度退化群落凋落物N含量無顯著差異,且均顯著高于中度和重度退化群落凋落物N含量,且后兩者之間并無顯著差異。潛在退化群落凋落物P含量顯著高于無退化、中度退化和重度退化群落,而輕度退化群落則與其他群落均無顯著性差異。隨退化程度加劇,凋落物C∶N和C∶P呈先降低后升高的趨勢,中度退化和重度退化程度群落凋落物C∶N和C∶P并無顯著差異,均顯著高于未退化、潛在退化和輕度退化群落,且后三者間亦無顯著差異。隨退化程度加劇,凋落物N∶P呈下降趨勢,未退化群落僅略高于潛退化和輕度退化群落,但顯著高于中度和重度退化群落,且潛退化和輕度退化群落及中度和重度退化群落間并無顯著差異。 隨退化程度的加劇,土壤C、N、P 含量均呈先升高后降低的趨勢(圖3)。潛在退化群落的土壤C含量顯著高于中度和重度退化群落,與未退化和輕度退化群落無顯著差異,且后四者差異不顯著。潛在退化群落土壤N、P含量顯著高于重度退化群落,且后者與其他三個群落之間N、P含量并無顯著差異。隨退化程度加劇,土壤C∶N、C∶P均呈先降低后升高變化趨勢;未退化和重度退化群落土壤C∶N差異不顯著,但均顯著高于其他三種退化程度群落,且后三者間亦無顯著差異;重度退化群落土壤C∶P顯著高于未退化群落,而未退化群落土壤C∶P亦顯著高于潛在退化、輕度退化和中度退化群落,但后三者間土壤C∶P無顯著差異。隨退化程度加劇,土壤N∶P總體呈升高變化趨勢,但不同退化程度群落之間土壤N∶P并無顯著差異。 圖3 喀斯特不同退化程度群落土壤C、N、P 含量及化學計量比Fig.3 Soil C、N、P contents and stoichiometric ratio in vegetation communities along degradation in karst area 由圖4分析可知,隨退化程度加劇,土壤微生物量C呈降低變化趨勢,而微生物量N、P則呈先升高而后降低變化趨勢。未退化群落土壤微生物量C顯著高于潛在退化和輕度退化群落、中度退化和重度退化群落,且前二者微生物量C亦顯著高于后二者,但前二者間及后二者間差異均不顯著。潛在退化群落土壤微生物量N、P與未退化和輕度退化群落差異均不顯著,但均顯著高于中度和重度退化群落,而后二者微生物量N差異不顯著,微生物量P差異顯著。隨退化程度加劇,土壤微生物量C∶N總體呈降低變化趨勢,土壤微生物量C∶P和N∶P則呈先降低后升高變化趨勢。不同退化程度群落土壤微生物量C∶N無顯著性差異;重度退化群落土壤微生物量C∶P與未退化群落無顯著差異,但顯著高于其他三種群落,且后四者之間土壤微生物量C∶P差異并不顯著;重度退化群落土壤微生物量N∶P顯著高于其他群落,其他四種群落之間則差異不顯著。 圖4 喀斯特不同退化程度群落土壤微生物生物量C、N、P 含量及化學計量比Fig.4 Soil microbial biomass C、N、P contents and stoichiometric ratio in vegetation communities along degradation in karst area 如圖5所示,植物葉片C與凋落物N、微生物量N呈極顯著正相關,與凋落物C∶N呈極顯著負相關,與微生物量C顯著正相關;植物葉片N與土壤N、微生物量N呈顯著正相關,與土壤C∶P、N∶P呈顯著負相關;植物葉片P與凋落物C、土壤N、微生物量C呈顯著正相關,與土壤P、微生物量C∶N呈極顯著正相關,與土壤C∶P、N∶P呈極顯著負相關;植物葉片C∶N與土壤C、N、微生物量C呈顯著負相關;植物葉片C∶P與凋落物C、土壤N、P、微生物量C∶N呈顯著或極顯著負相關,與土壤C∶N、C∶P、N∶P呈顯著或極顯著正相關;植物葉片N∶P與凋落物N、N∶P呈顯著正相關,與土壤P呈顯著負相關。土壤C與凋落物N、N∶P、微生物量C、N呈顯著或極顯著正相關;土壤N與凋落物N∶P、微生物量P呈顯著正相關。凋落物N與微生物量C、N呈極顯著正相關;凋落物P與微生物量C、凋落物N∶P與微生物量N顯著相關;凋落物C∶N與微生物C、N呈顯著或極顯著負相關。 圖5 群落退化過程中植物葉片、凋落物、土壤、微生物量C、N、P含量與化學計量比的關系Fig.5 Relationships of leaf, litter, soil and microbial biomass C, N, P contents and stoichiometry during the degradation process 喀斯特植被退化過程中,隨著土壤N、P含量及N∶P的變化,葉片N、P含量及N∶P利用穩態性模型模擬結果均顯著(P<0.1),呈不同的內穩性特征(圖6)。在土壤N、P含量變化下,植物葉片N元素的內穩性指數H分別為2.72和2.75,平均值為2.73,屬于弱穩態型;植物葉片P元素的內穩性指數H則分別為2.39和2.23,平均值為2.31,屬于弱穩態型。植物葉片N∶P內穩態指數H為5.14,屬于穩態型。綜合分析表明,植物葉片N元素的內穩性略高于P元素,但明顯低于N∶P,也即,相對于N、P元素內穩定性,元素計量比N∶P具有更高的內穩性。 圖6 土壤N、P含量、N∶P 與植物葉片N、P含量、N∶P的關系Fig.6 Relationship between soil N, P contents and leaf N and P contents 葉片是植物對環境變化做出反應的最敏感的指示部位[23]。植物葉片C 含量越高表明其比葉重越大,具有較強的防御能力,葉片N、P 含量越高,表明其資源競爭能力越強[24—25]。隨群落退化程度的加劇,植物葉片的C、N、P含量呈下降趨勢,表明喀斯特植被群落退化過程中,植物葉片養分含量降低,對環境變化的防御能力和資源競爭能力下降。葉片N、P含量與土壤N、P含量之間存在顯著和極顯著正相關關系,而與土壤C∶P、N∶P則呈顯著和極顯著負相關關系,表明植物的營養元素主要來源于土壤,其含量和土壤中含量密切相關[26];土壤C∶P、N∶P越高,表明土壤可利用性N、P含量越低[27],進而影響其葉片含量。 葉片是植物的主要光合器官,因此葉片的化學計量比能代表植物整株的狀況[28]。葉片C∶N、C∶P比代表著植物吸收N、P元素時所能同化C的能力及固C效率的高低,可反映植物的生長速率和養分利用率,較高的C∶N、C∶P 對應較低的生長速率和較高的養分利用率[29—30],一定程度上也反映了土壤中N和P的供應狀況[26]。植物在營養元素供應缺乏的情況下往往具有較高的養分利用效率,反之,營養元素供應充足, 元素利用效率則較低[31]。重度退化群落植物葉片C∶N、C∶P顯著高于其他群落,表明當群落發生重度退化后,植物同化C的能力和固C速率顯著性下降,植物的生長速率也顯著降低,因而養分利用效率提高,可能原因是一方面群落退化后養分歸還量減少,另一方面植被減少引起水土流失而導致養分流失,最終引起土壤養分供應不足。植物葉片的N∶:P臨界值被認為可以作為判斷環境對植物生長的養分供應狀況的指標[4],當植物N∶ P<14 時,植物生長表現為受N限制;當N∶ P >16 時,表現為受P 限制;當14 凋落物是群落中聯結土壤和植物的重要組成部分,其養分含量受群落優勢植物及凋落物組成的影響[34]。本研究中,隨退化程度的加劇,凋落物C含量呈先降低后升高再降低的趨勢,N、P含量則表現為先增后減的變化趨勢,這可能與不同退化程度群落中優勢種組成不同有關。有關研究表明,當凋落物N<7 g/kg,P<0.5 g/kg時,凋落物N、P養分被葉片完全吸收;而N>10 g/kg、P>0.8 g/kg/時,則表明沒有完全吸收[35—36]。本研究中,中度、重度退化群落凋落物N含量(4.64 g/kg、4.13 g/kg)低于7 g/kg,P含量(0.40 g/kg、0.41g/kg)均低于0.5 g/kg,表明植被群落退化達中度以上,會發生凋落物N、P的完全吸收,這說明群落退化后植物通過提高養分重吸收效率來適應土壤環境的變化。然而,退化群落凋落物N、P含量低的重要的原因可能是秉承了植物的特性[26],即植物的養分含量降低。凋落物N含量與葉片N∶P顯著正相關,說明植物葉片N∶P越高時,植物生長N供應充足,凋落物N含量越高。 凋落物C∶N是預測凋落物分解速率最理想的指標[37],較低的C∶N對應較高的凋落物分解速率[38—39]。本研究中,隨著退化程度的加劇,凋落物C∶N總體上呈增加趨勢,說明喀斯特地區植被群落退化過程中,凋落物的分解速率降低,養分回歸量減少。有研究表明,凋落物N∶P越高,凋落物的分解受到P素的限制越強,尤其是當凋落物的N∶P大于25或其P含量低于0.22 g/kg時[40]。本研究中,不同退化程度植被群落凋落物N∶P均小于25,且隨退化程度加劇而下降,說明喀斯特區植被群落凋落物分解受P的限制小,并在群落退化過程中逐漸減小。凋落物N∶P與葉片N∶P之間顯著正相關,可見凋落物分解與植物生長的養分限制類型有關。 植被退化過程中,隨植物群落物種組成的改變,土壤的養分特征也發生變化[41]。土壤C、N與凋落物N、N∶P存在極顯著或顯著正相關關系,可能原因是凋落物N、N∶P能夠影響其分解速率[38],從而影響土壤C、N含量,進一步說明土壤中N含量與凋落物密切相關,而P含量則與土壤中P含量相關性不顯著,可能原因是兩者的來源不同,土壤N主要來源于凋落物歸還,而土壤中的P不僅來自于凋落物的輸入,還與土壤成土母質中的礦物成分密切相關[42]。土壤C與微生物量C、N和土壤N與微生物量P存在極顯著或顯著正相關關系,說明微生物量C、N是土壤C、N動態的敏感指示者[43]。 土壤的 C∶N 可以衡量其 C、N 的營養平衡狀況,并能影響其C、N 養分循環,是反映土壤質量的敏感指標[34,44],一般而言,土壤較低C∶N 表現為高肥力和較快的C、N礦化速率[45]。本研究中,中度退化群落土壤 C∶N 最低,但并不意味著其土壤肥力高,C、N、P不平衡性增加或降低也會引起土壤C∶N、C∶P、N∶P的變化[46],隨著植被退化,土壤C含量的降幅高于N含量的降幅,從而C∶N 減小;而當植被退化到一定程度,達到重度退化,土壤中的N元素顯著下降,因此重度退化群落土壤C∶N顯著升高。較低的C∶P比是磷有效性高的一個指標[27]。隨著退化程度的加劇,土壤C∶P不斷升高,重度退化程度最高,說明重度退化群落P有效性顯著降低。土壤N∶P可以作為養分限制類型的有效預測指標[27],不同退化程度群落土壤N∶P 之間差異不顯著,說明植被群落發生退化,植被群落的養分限制格局沒有改變。 土壤微生物是有機質的分解者,微生物量的任何變化,都會影響土壤養分的循環和有效性,能夠及時反映土壤的質量狀況[47]。本研究中,土壤微生物量隨群落退化而變化的結果表明,植被群落退化后,群落穩定性下降,水熱條件、相對濕度和溫度等波動較大,尤其是重度退化群落,植被層次極不完整、生物多樣性低,群落輸入的有機質較少,因而土壤微生物生物量最低[48]。微生物C、N是衡量土壤微生物活性的重要指標,其數值的高低直接說明土壤肥力的好壞[34]。微生物量C、N與凋落物N含量呈極顯著正相關,與凋落物C∶N則顯著或極顯著負相關,這是因為凋落物C∶N通常用來表示凋落物的質量,其對微生物分解和礦化作用有較大的影響,凋落物N含量高時對應較低的C∶N,其礦化和腐化都較易進行,分解快[25],微生物活性高。 土壤微生物量C∶N比值是否恒定,不同學者的觀點各異。王紹強等[27]認為土壤微生物生物量的C∶N比約為10∶1,本研究中喀斯特地區不同退化程度群落土壤微生物生物量C∶N分布在8.27—11.01之間,平均值為9.32,與之接近。不同退化程度群落土壤微生物量C∶N 差異不顯著,說明土壤微生物具有較高的內穩性。土壤微生物量C∶P 可作為衡量微生物礦化土壤有機質釋放P或從土壤中吸收固持P 潛力的重要指標,比值一般在7—30之間[49]。本研究中重度退化群落土壤微生物量C∶P(19.32)較高,表明微生物與植物競爭土壤有效磷,出現固磷現象;其他群落土壤微生物量C∶P偏低,說明微生物在礦化土壤有機質中釋放P 的潛力較大。 土壤養分含量的變化可以直接影響植物養分的吸收及其化學計量比,而植物則通生態化學計量及養分利用策略調節來適應土壤N、P含量的變化[50-51]。內穩態理論認為生命有機體能夠維持自身體內化學元素組成不隨外部環境的變化而發生劇烈改變,而是將其體內化學元素組成保持在相對狹窄的范圍內[52]。內穩態特征是生態化學計量學研究的基礎,內穩性的強弱反映了生物在長期進化過程中對環境變化的生理和生化調節的適應程度[53]。本研究中植物葉片N、P元素均為弱穩態型,N∶P則為穩態型,表現為HN∶P>HN>HP,表明喀斯特地區植物具有一定的內穩性,可以通過調節自身養分元素的平衡,使元素比值處于相對穩定的狀態。同時在一定程度上證實了限制性養分元素穩定性假說:由于生理和養分平衡的制約,限制性養分元素在植物體內的含量具有相對穩定性, 對環境變化的響應也較為穩定[1],這表明隨著喀斯特地區植被群落退化,在N養分限制的環境中,植物葉片N具有較強的內穩性。 桂西北喀斯特地區植被群落退化過程中,植物葉片、凋落物、土壤、微生物量C、N、P含量及其化學計量比發生明顯變化,植物對環境變化的防御能力和資源競爭能力下降;植物的生長速率顯著降低,而養分利用效率提高;植物生長主要受N限制,且退化越嚴重,N限制作用越明顯;凋落物C∶N總體上呈增加趨勢,說明喀斯特地區植被群落退化過程中,凋落物的分解速率降低,養分回歸量減少;重度退化群落土壤P有效性顯著降低;植被群落發生退化,植被群落的養分限制格局沒有改變;重度退化群落土壤微生物量C∶P偏高,表明微生物與植物競爭土壤有效磷,出現固磷現象,而其他群落土壤微生物在礦化土壤有機質中釋放P 的潛力較大。葉片N、P元素均為弱穩態型,N∶P則為穩態型,表明喀斯特地區植物具有一定的內穩性,可以通過控制自身養分元素的平衡,使其元素組成比值保持相對穩定的狀態。研究結果對深入理解喀斯特植被退化過程中,生態系統各組分(植物葉片、凋落物、土壤、微生物)養分狀況和養分循環具有重要意義。但本研究未考慮不同退化程度植物群落化學計量特征種間差異與種內特征及其指示作用,因此進一步對不同退化群落中相同植物種類間化學計量特征的差異進行研究具有重要意義。2 結果與分析
2.1 植物葉片C、N、P 含量及化學計量比

2.2 凋落物C、N、P 含量及化學計量比
2.3 土壤C、N、P 含量及化學計量比
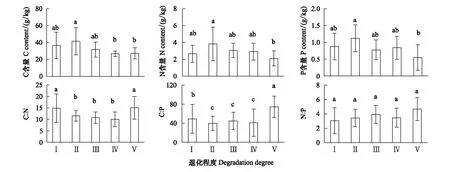
2.4 土壤微生物量C、N、P 含量

2.5 植物-凋落物-土壤-微生物C、N、P及化學計量比的相關性

2.6 葉片N、P含量及N∶P比隨土壤養分變化的內穩態分析
3 討論
3.1 不同退化程度群落植物葉片C、N、P 含量及化學計量特征
3.2 不同退化程度群落凋落物C、N、P 含量及化學計量特征
3.3 不同退化程度群落土壤C、N、P 含量及化學計量特征
3.4 不同退化程度群落土壤微生物生物量C、N、P 含量及化學計量特征
3.5 植物葉片N、P含量及N∶P內穩性特征
4 結論
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22學苑創造·A版(2020年3期)2020-04-24 09:21:39小溪流(畫刊)(2017年11期)2018-01-09 19:15:14少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33紅領巾·萌芽(2017年5期)2017-06-23 10:35:59爆笑show(2016年7期)2017-02-09 09:36:13
