青藏高原不同牧草人工草地對土壤線蟲群落的影響
2022-03-07 09:06:54吳鵬飛
生態學報 2022年3期
關鍵詞:植物
魏 雪,李 雨,吳鵬飛
西南民族大學青藏高原研究院, 成都 610041
青藏高原作為“世界第三極”,是生物多樣性熱點區域以及重要的生態安全屏障區[1]。其天然草地面積約占全國草地面積的三分之一,是我國重要的草牧業生產基地[2]。然而,青藏高原地處生態脆弱區,近幾十年來受氣候變化和過度放牧等的共同影響[3],高寒草地生態系統退化嚴重,導致畜牧生產和生物避難所等重要生態系統服務功能下降[4]。近年來,國家采取了施肥、補種、禁牧圍封和建植人工草地等措施對退化草地進行恢復,其中建植人工草地最為有效[5]。建植人工草地也是解決草畜矛盾的主要途徑[6]。因此人工草地,尤其是單播人工草地在該區得到快速發展[7]。
土壤線蟲數量巨大,占全部多細胞生物數量的80%[8];其物種豐富、分布廣泛,在有機質分解和養分循環方面發揮關鍵作用[9—10],是草地生態系統的重要組成部分。土壤線蟲食性多樣,主要有植食性、食真菌、食細菌、捕食性和雜食性等,在土壤食物網中占有重要地位[11]。其次,線蟲身體透明,內部構造清晰,易于鑒定。同時,線蟲世代周期較短,遷移能力弱,對環境變化敏感,因此被廣泛用作反映生態系統受干擾程度和土壤健康水平的指示生物[12]。
前期研究表明,青藏高原天然草地向人工草地轉變,顯著改變了植物群落組成[7]和土壤碳、氮儲量[5,13]。植物作為土壤食物網的主要生產者和資源提供者,極大地影響著土壤線蟲群落及其生態功能[14—15]。已有研究表明不同植物下的植食性線蟲(針屬Paratylenchus)密度差異顯著[16],甚至某些植食性線蟲只有在特定植物存在時數量才會增加[17—19]。不同物種植物的根系分泌物[20]、根際土壤理化性質[21]和微生物群落[22]存在差異,可間接影響食微線蟲[23—24]。植物功能群也會對土壤線蟲群落產生影響,已有研究表明豆科植物對食細菌線蟲有利,而雜類草對食真菌線蟲有利[25—26]。Ye等[27]研究發現,與禾本科相比,豆科單作和禾本科-豆科混作增加了植食性、食細菌和食真菌線蟲的密度。此外,植物群落多樣性也可影響土壤線蟲群落多樣性。有研究表明線蟲的多樣性隨著植物多樣性的增加而增加[28],但也有研究發現某些植食性線蟲在單一植物栽培中的數量比多種植物混合栽培中更多[24,27]。前期研究發現青藏高原天然草地轉變為燕麥人工草地后改變了線蟲群落結構,增加了線蟲密度以及食細菌和食真菌線蟲的比例[29],然而不同人工草地土壤線蟲群落組成及多樣性的差異尚不明確。
在青藏高原地區選取廣泛建植的6種牧草單播人工草地,通過調查不同人工草地和天然草地的土壤線蟲群落,研究天然草地轉變為人工草地對土壤線蟲群落的影響以及不同人工草地土壤線蟲群落多樣性差異。研究結果不僅有助于揭示建植人工草地對高寒草甸地下食物網結構和功能的影響,還可促進生態環境的保護以及民族地區畜牧業的可持續發展。
1 材料與方法
1.1 研究區概況
研究區位于青藏高原東緣阿壩藏族羌族自治州紅原縣境內(31°50′—33°22′N,101°51′—103°23′E),平均海拔3500 m。該地是典型的高原大陸性氣候,霜凍期極長,無絕對無霜期,春秋短促,冬季寒冷,雨熱同季。日照時間長,年均達2389 h,年均蒸發量為1232 mm。年均氣溫0.9 ℃;最熱月為7月,月均氣溫為10.9 ℃;最冷月為1月,月均氣溫為-10.3 ℃。降水量年際間波動大,年均降水量約為680 mm,80%集中在生長季階段。植被類型主要為亞高山草甸,蓋度為70%—90%,優勢種為高山嵩草(KobresiapygmaeaC. B. Clarke)和四川嵩草(K.setschwanensisHand.-Mazz.),常見類群有高山紫菀(AsteralpinusL.)、蕨麻(PotentillaanserinaL.)、草玉梅(AnemonerivularisBuch.-Ham.)、垂穗披堿草(ElymusnutansGriseb.)、老芒麥(E.sibiricusL.)和紫羊茅(FestucarubraL.)等為主。
1.2 樣地設置
本研究在西南民族大學青藏高原生態保護與畜牧業高科技研究示范基地(32°48′N,102°33′E)2013年建植的種質資源圃內進行。2016年7月,選取垂穗披堿草(E.nutans,EN)、老芒麥(E.sibiricus,ES)、早熟禾(PoaannuaL.,PA)、羊茅(F.ovinaL.,FO)等多年生禾本科牧草和連續播種的一年生禾本科牧草燕麥(AvenasativaL.,AS)以及多年生豆科牧草紫花苜蓿(MedicagosativaL.,MS)6種單播人工草地作為研究對象[30]。每種牧草人工草地2塊樣地,面積為2 m × 4 m[30]。同時,在資源圃外圍兩側另選2塊2 m × 4 m的天然草地(Natural grassland,NG)作為對照[30]。在每塊樣地內按對角線法設置3個50 cm × 50 cm的樣方。本研究共7種處理,每種處理6個重復。
天然草地的植物優勢種為垂穗披堿草、平車前(PlantagodepressaWilld.)、早熟禾和珠芽蓼(PolygonumviviparumL.),蓋度為75%—90%。各人工草地的施肥、除雜和收割等田間管理措施相同。人工草地均采取春季苗期施肥,施肥標準為含氮量35%的硝酸銨(NH4NO3)33 g/m2和含P2O514%的過磷酸鈣磷(Ca(H2PO4)2·H2O)40 g/m2;每年播種期、苗期對人工草地進行人工除雜。每年9月對人工草地和天然草地進行牧草刈割,留茬4 cm左右。
1.3 土壤樣品采集和線蟲分離及鑒定
在每個樣方內用土鉆采集3個0—15 cm的土樣作為一個復合樣,混合均勻帶回室內。稱取50 g鮮土,采用濕漏斗(Baermann)法分離土壤線蟲,連續分離48 h。分離到的線蟲保存于4%的福爾馬林溶液中。根據《中國土壤動物檢索圖鑒》[31]和《De Nematoden van Nederland》[32]等,在正置顯微鏡(Olympus CX31)下對土壤線蟲進行計數和鑒定。每個樣品隨機選取100條線蟲進行鑒定,不足100條的全部鑒定,一般鑒定到屬。根據土壤含水量,將土壤線蟲密度換算成每100 g干土中的個數。并根據食性將線蟲分為植食性、食真菌、食細菌和雜食-捕食性等4個營養類群[33—34]。
1.4 土壤理化性質分析
土壤含水量(WC)采用烘干法測定;土壤有機碳(SOC)含量采用硫酸-重鉻酸鉀氧化外加熱法進行測定;全氮(TN)含量采用半微量開式定氮法測定;有效氮(AN)含量采用堿解擴散法測定;全磷(TP)含量采用高氯酸-硫酸-鉬銻抗比色法測定;有效磷(AP)含量采用碳酸氫鈉浸提-鉬銻抗比色法測定;全鉀(TK)含量采用氫氟酸-高氯酸消煮火焰光度計法測定;有效鉀(AK)含量采用中性乙酸銨提取-火焰光度計法測定;土壤pH采用電位法測定[35]。
1.5 數據分析
根據線蟲各類群的密度占總密度的比例來劃分數量等級,10%以上的為優勢類群,1%—10%的為常見類群,1%以下的為稀有類群。
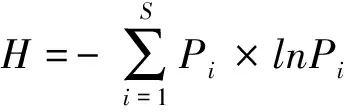
自由生活線蟲成熟度指數MI(Maturity index)[36]:

植物寄生線蟲成熟度指數PPI(Plant parasite index)[36]:
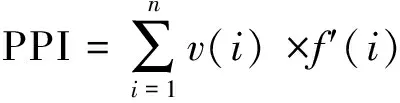
式中v(i)為第i類線蟲的c-p值,f(i)為第i類占總數的比例,f′(i)為第i類植物寄生線蟲占總數的比例。
基礎指數BI(Basal index)[11]:
通道指數CI(Channel index)[11]:
富集指數EI(Enrichment index)[11]:
結構指數SI(Structure index)[11]:
式中,b=∑kb×nb,e=∑ke×ne,s=∑ks×ns。b為基礎組分,包含Ba2(食細菌線蟲中的c-p2類群)和Fu2(食真菌線蟲中的c-p2類群);e為富集組分,包含Ba1(食細菌線蟲中的c-p1類群)和Fu2;s為結構組分,包含Ba3-Ba5、Fu3-Fu5、Om3-Om5和P2-P5(c-p值范圍為3—5的食細菌、食真菌和雜食性線蟲以及c-p值范圍為2—5的捕食性線蟲)。kb、ke和ks為各類群所對應的權重值(值為0.8—5.0之間),nb、ne和ns為各類群的相對密度。BI指示土壤食物網的抵抗力,值越大,相對抵抗力越大;CI指示土壤食物網中占優勢的分解通道,值大于50表明以真菌分解通道為主,值小于50表明以細菌分解通道為主;EI指示外界養分投入狀況,值越大,外界養分投入越多;SI指示食物鏈的長度及食物網的連通性,值越大,食物鏈越長,連通性越高[11,37]。EI與SI聯合使用,以SI為橫坐標,EI為縱坐標,可以將線蟲區系劃分為A、B、C和D 4個象限,分別代表了食物網不同的特征[11]。
采用car軟件包的qqplot函數對數據進行正態性檢驗,以及用Bartlett檢驗進行方差齊性檢驗。對于不符合正態分布的數據采用car包的powerTransform函數進行Box-Cox正態變換。為研究不同牧草人工草地對土壤線蟲群落密度、類群數、各營養類群密度和生態指數以及土壤理化性質等參數的影響,對符合假設檢驗的參數采用單因素方差分析(One-way ANOVA)進行分析,不符合的則直接進行非參數Kruskal-Wallis檢驗。方差分析用aov函數進行,對差異顯著的數據采用multcomp包的glht函數進行多重比較。Kruskal-Wallis檢驗用kruskal.test函數進行,對差異顯著的數據采用FSA包的dunnTest函數進行多重比較。
采用去趨勢對應分析(Detrended correspondence analysis,DCA)對樣方及群落進行排序,第一排序軸長度小于3,因此選用冗余分析(Rundancy analysis,RDA)對土壤線蟲屬的分布與環境因子間的關系進行排序。去趨勢對應分析和冗余分析分別采用vegan包的decorana和rda函數進行。此外,采用全子集多元回歸分析(Multiple regression analyses)探索土壤線蟲群落與環境因子間的關系,采用lm函數以及leaps包的regsubsets函數進行。以上數據分析均在R 3.6.1(R Development Core Team)中進行。
2 結果
2.1 土壤線蟲群落組成
本研究分離到的土壤線蟲隸屬于2綱8目32科58屬(附表1)。土壤線蟲平均密度為1754個/100 g干土;其中紫花苜蓿樣地的密度最低,為949個/100 g干土;燕麥樣地最高,為3267個/100 g干土(附表1)。在科水平上,墊刃科(Tylenchidae)和奎希婭科(Qudsianematidae)為優勢類群,分別占總密度的15.73%和14.41%,紐帶科(Hoplolaimidae)和頭葉科(Cephalobidae)等16科為常見類群,共占總密度的64.53%;諾帝科(Nordiidae)和細齒科(Leptonchidae)等14科為稀有類群,共占總密度的5.32%(附表1)。在屬的水平上,沒有發現優勢類群(附表1)。
不同草地的土壤線蟲群落優勢類群存在一定差異(附表1)。在屬水平上,紫花苜蓿樣地的優勢類群為絲尾墊刃屬(Filenchus)(占11.86%);天然草地為小矛線屬(Microdorylaimus)(占10.55%)和真滑刃屬(Aphelenchus)(占10.15%);而其他人工草地無優勢屬(附表1)。
2.2 土壤線蟲密度和多樣性
線蟲密度、類群數和Shannon多樣性指數均在燕麥樣地最高,Simpson優勢度指數在燕麥樣地最低(圖1)。分析結果表明,燕麥樣地的線蟲密度與老芒麥樣地無顯著差異,但顯著高于其他樣地;與天然草地相比,燕麥樣地的線蟲密度增加了93.47%(圖1)。燕麥樣地的線蟲類群數和Shannon多樣性指數顯著高于天然草地,其Simpson優勢度指數顯著低于垂穗披堿草、紫花苜蓿樣地和天然草地;此外,羊茅樣地的線蟲Simpson優勢度指數顯著低于天然草地NG(圖1)。
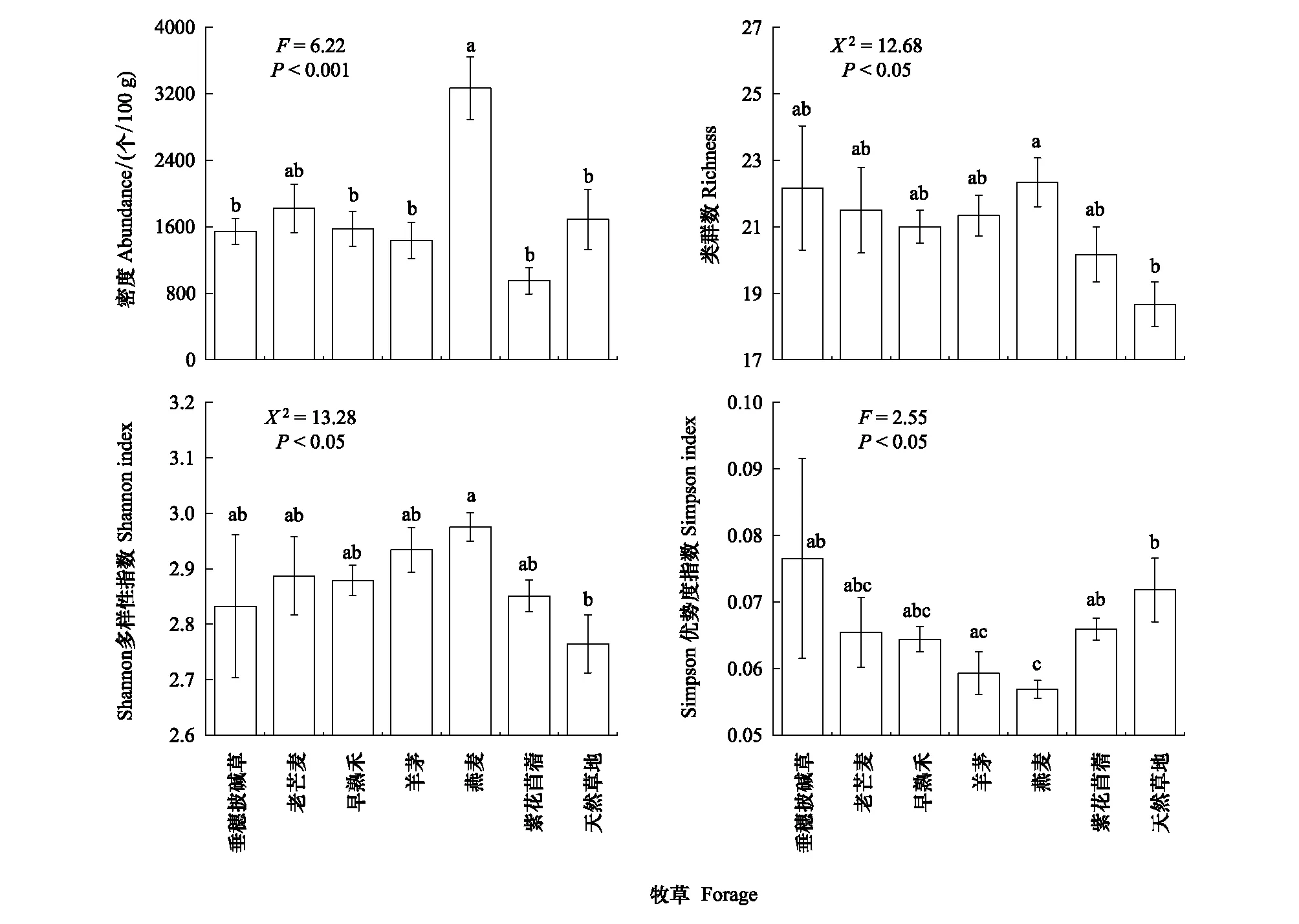
圖1 不同人工草地和天然草地的土壤線蟲群落密度和多樣性(平均值±標準誤)Fig.1 The abundance, taxonomic richness, Shannon index and Pielou index of the soil nematode communities in the artificial grasslands of the different forage species and the natural grasslands (Mean±SE) 不同小寫字母表示不同樣地間差異顯著(P<0.05)
2.3 土壤線蟲營養類群結構
燕麥樣地的各營養類群密度均為最高,其植食性和雜食-捕食性線蟲密度顯著高于天然草地,分別增加了179.42%和113.66%;其食真菌線蟲密度顯著高于垂穗披堿草、老芒麥、羊茅和紫花苜蓿樣地,其食細菌線蟲密度顯著高于早熟禾、羊茅和紫花苜蓿樣地(圖2)。此外,老芒麥和早熟禾樣地的雜食-捕食性線蟲密度顯著高于紫花苜蓿樣地(圖2)。

圖2 不同人工草地和天然草地的土壤線蟲營養類群密度(平均值±標準誤)Fig.2 The abundances of the trophic groups of the soil nematode communities in the artificial grasslands of the different forage species and the natural grasslands (Mean±SE)
2.4 土壤線蟲生態指數
禾本科牧草的自由生活線蟲成熟度指數呈增加趨勢,其中早熟禾和羊茅樣地顯著高于天然草地(圖3)。紫花苜蓿樣地的植物寄生線蟲成熟度指數PPI最低,顯著低于天然草地(圖3)。與天然草地相比,垂穗披堿草、老芒麥、早熟禾和羊茅等多年生禾本科牧草樣地的線蟲基礎指數BI顯著下降,但各人工草地的通道指數均無明顯變化(圖3)。此外,垂穗披堿草樣地的線蟲通道指數小于50,表明以細菌分解通道為主;而其他樣地大于50,表明以真菌分解通道為主。
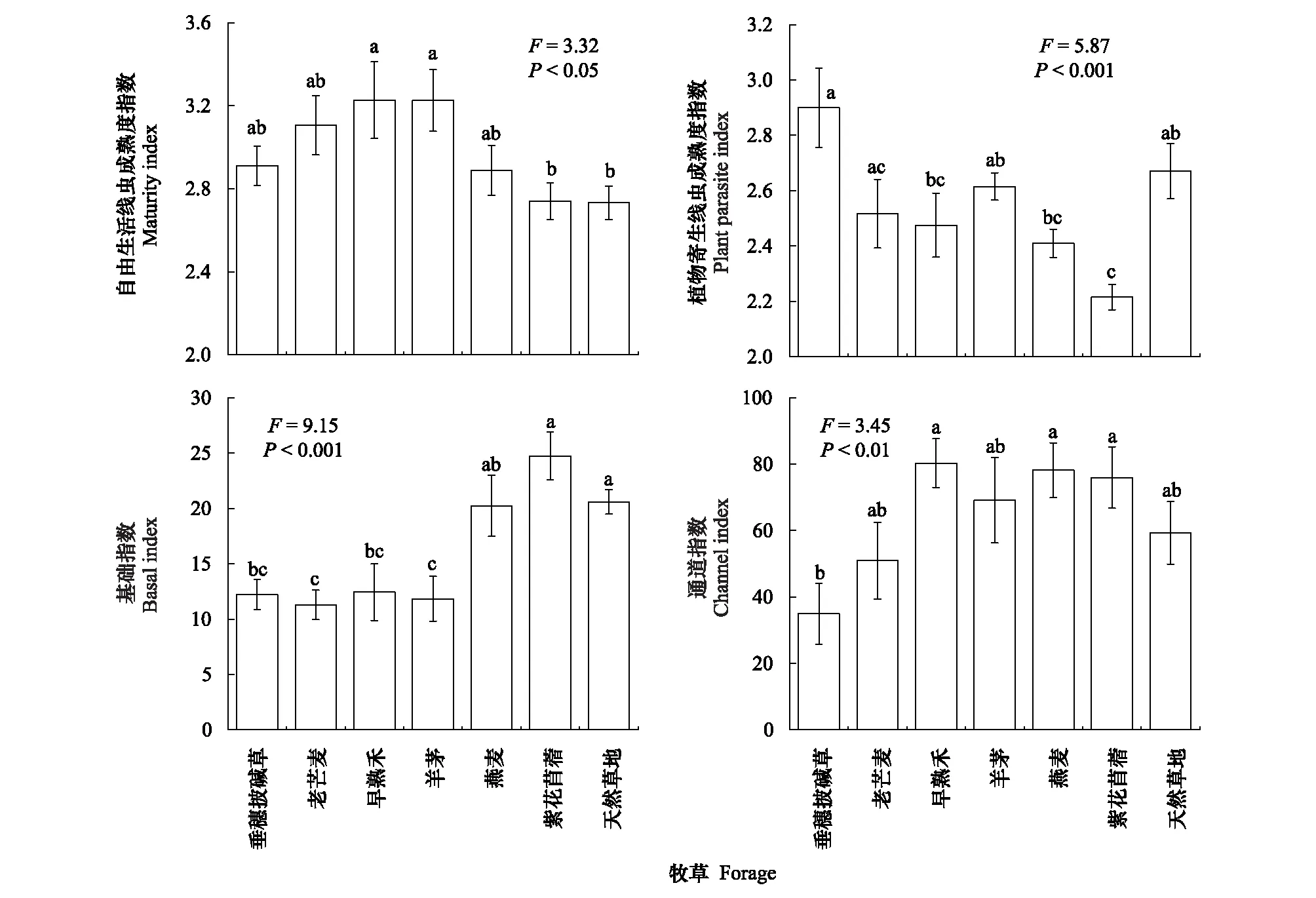
圖3 不同人工草地和天然草地的土壤線蟲生態指數(平均值±標準誤)Fig.3 The MI, PPI, BI and CI of the soil nematodes community in the artificial grasslands of the different forage species and the natural grasslands (Mean±SE)
不同人工草地的線蟲富集指數和結構指數具有顯著差異(圖4),表明植物物種對養分富集狀況和土壤食物網的結構的影響不同。其中,紫花苜蓿樣地的線蟲富集指數和結構指數最低。天然草地、早熟禾和羊茅樣地的線蟲區系處于B和C象限之間,表明土壤養分狀況較好,食物網穩定成熟;與天然草地相比,早熟禾和羊茅樣地結構指數增加,表明食物鏈更長,連通性更高。垂穗披堿草和老芒麥樣地的線蟲區系位于B象限,表明土壤養分狀況好,食物網成熟。燕麥和紫花苜蓿樣地的線蟲區系位于C象限,表明土壤養分富集狀況中等,食物網處于結構化狀態。
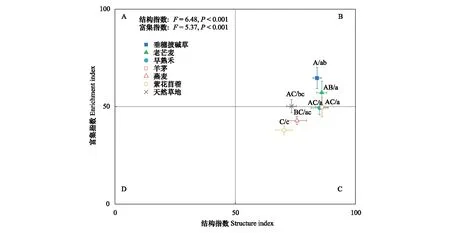
圖4 不同人工草地和天然草地的線蟲區系分析(平均值±標準誤)Fig.4 Nematode faunal analysis of the different artificial grasslands and the natural grasslands (Mean±SE) 誤差線上不同字母表示不同樣地間的富集指數(大寫)或結構指數(小寫)差異顯著(P<0.05)
2.5 土壤環境因子和土壤線蟲群落的關系
土壤含水量、有機碳、全氮、有效氮、全磷、有效磷、全鉀、有效鉀和pH在樣地間均具有顯著差異(表1)。老芒麥樣地的土壤含水量顯著高于紫花苜蓿樣地。土壤有機碳、有效氮、有效磷、有效鉀均在天然草地最高,全氮、全磷、全鉀均在羊茅樣地最高,土壤pH在垂穗披堿草樣地最高。

表1 不同人工草地和天然草地的土壤理化性質(平均值±標準誤)
冗余分析結果表明,第一排序軸的解釋量具有顯著性(P<0.01)。第一軸與全鉀(r=0.80,P<0.05)顯著正相關,與碳氮比(r=-0.98,P<0.01)顯著負相關;第二軸與全磷(r=0.95,P<0.001)和有效鉀(r=0.78,P<0.001)顯著正相關(圖5)。天然草地與6種人工草地的土壤線蟲群落組成存在差異,尤其是與垂穗披堿草、早熟禾、羊茅和紫花苜蓿樣地明顯不同。此外,紫花苜蓿樣地與其他樣地的土壤線蟲群落組成存在明顯差異。
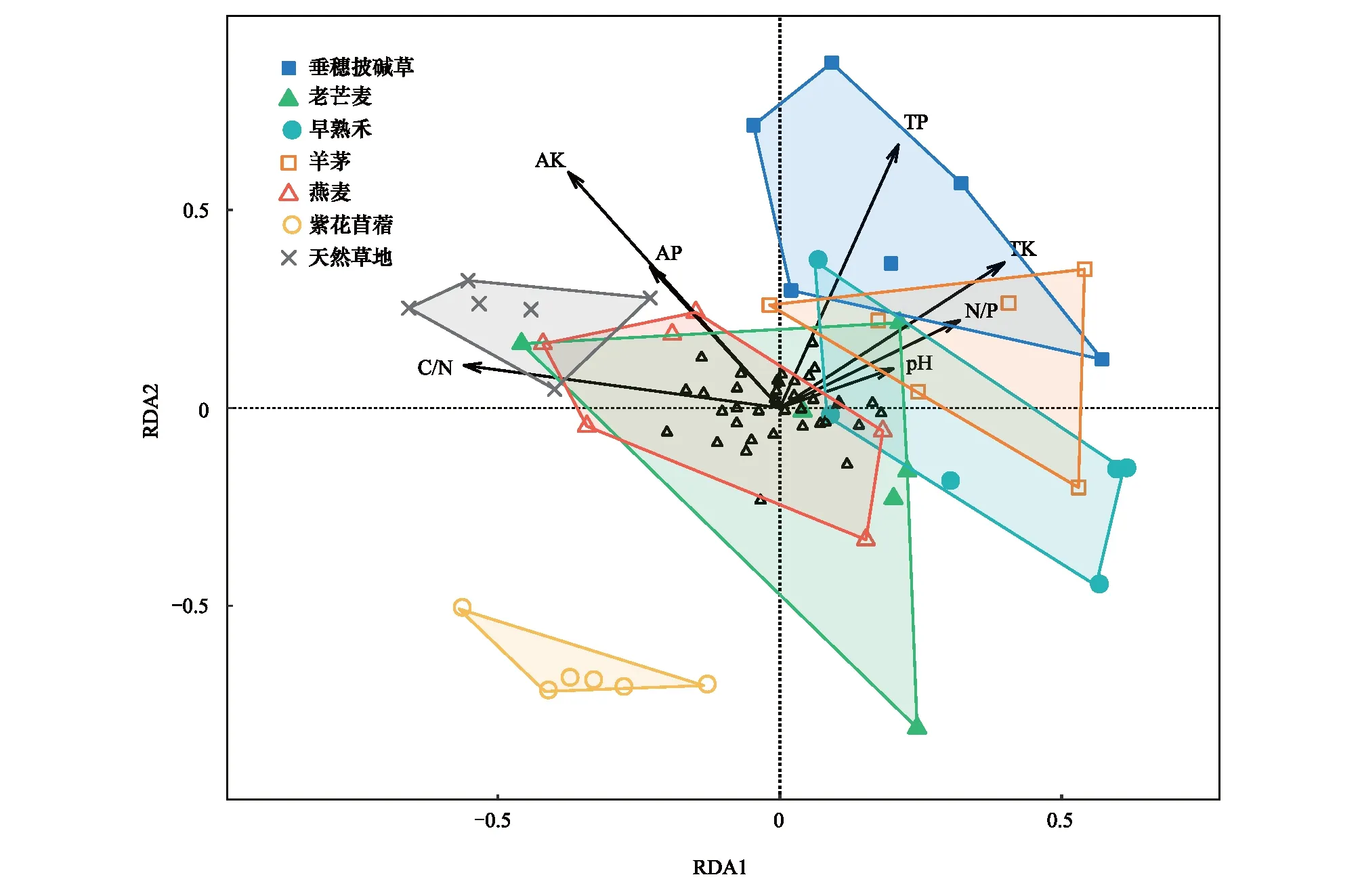
圖5 土壤線蟲群落與土壤環境因子的冗余分析Fig.5 Ordination diagram from the redundancy analysis (RDA) on the relationships between the soil nematode communities and the soil parameters TP:全磷 Total phosphorus;TK:全鉀 Total potassium;AP:有效磷 Available phosphorus;AK:有效鉀 Available potassium;黑色箭頭代表土壤參數,空心三角形代表土壤線蟲的屬
多元回歸分析結果表明土壤線蟲密度和土壤有效鉀含量呈顯著正相關關系,與碳磷比呈顯著負相關關系;線蟲類群數與土壤碳氮比呈顯著負相關關系(表2)。在所觀測的土壤環境因子中,有效鉀是影響土壤線蟲群落最主要的因素。
3 討論
3.1 人工草地與天然草地間土壤線蟲群落差異
本研究發現,天然草地轉變為人工草地后土壤線蟲的群落組成、密度和多樣性發生變化,其中燕麥樣地線蟲群落的密度、類群數和Shannon多樣性指數以及植食性和雜食-捕食性線蟲密度均顯著增加,而其他人工草地無顯著變化(圖1—2)。這與楊銳等[29]的研究結果相似。天然草地轉變為人工草地后,植物的物種、群落多樣性、生物量和蓋度等的變化是影響土壤線蟲群落的重要因素。首先,與天然草地相比,人工草地植物物種單一,多樣性降低。理論上植物多樣性的降低可通過食物資源的多樣性(凋落物的質量和組成)降低和生境簡化[24,38]降低線蟲群落的多樣性[24],然而很多研究發現線蟲多樣性很少受到植物多樣性的影響,植物物種對線蟲群落的影響更大[26,28,39]。已有研究發現,單一作物處理下的某些植食性線蟲數量比多種作物混種處理下更多[24,27],這與不同物種的植物營養成分[40]、根系分泌物[20]、微生物群落[22]和土壤理化性質[21]不同有關。本研究中不同牧草的營養成分及其根系分泌物和微生物情況尚不清楚,需進一步分析。其次,轉變為人工草地后,植物生物量的變化會改變食物資源的數量,進而影響線蟲增長[41]。燕麥的地上生物量和總生物量較高[42],是植食性線蟲密度和總密度增加的原因之一。其次,垂穗披堿草和老芒麥為疏叢型上繁草,早熟禾為根莖-疏叢型下繁草,羊茅為矮生-密叢型下繁草,燕麥為疏叢型上繁草,紫花苜蓿為直立型上繁草[40],不同植物的分蘗特性和株叢形狀不同會影響植物群落蓋度。燕麥樣地的蓋度高于天然草地[42],已有研究表明蓋度的增加會降低土壤溫度同時增加濕度[43],進而可能影響土壤線蟲群落。
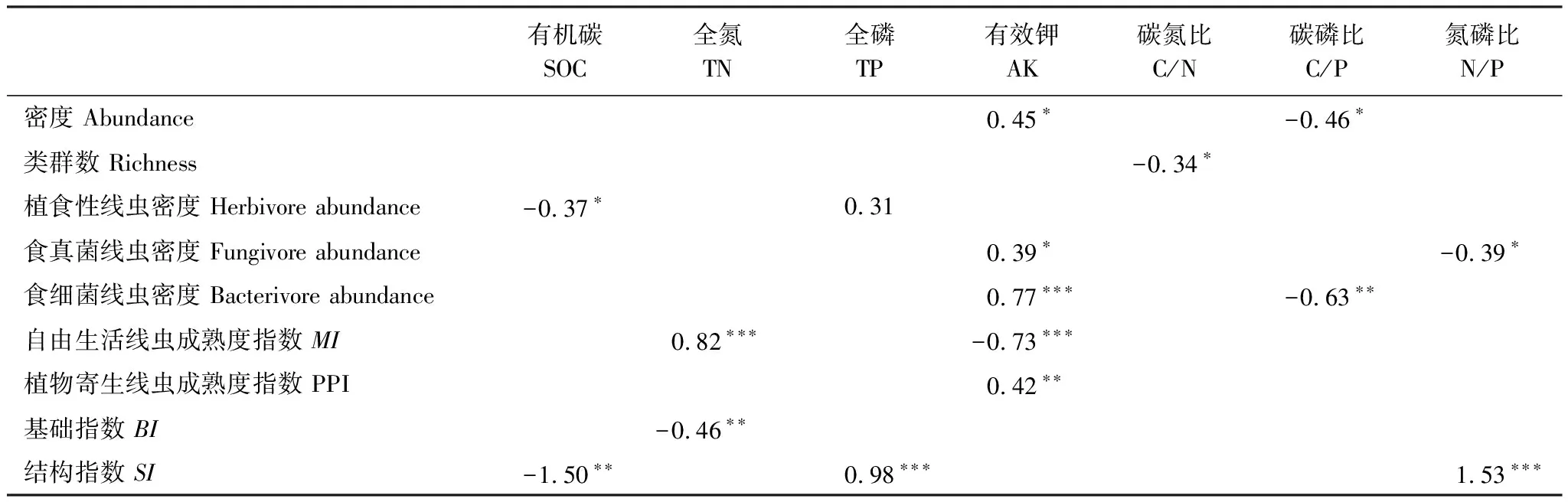
表2 土壤線蟲群落與土壤化學性質的多元回歸分析的回歸系數
此外,天然草地轉變為人工草地后,播種、翻耕、施肥和除雜等田間管理措施以及植物群落的變化會改變土壤理化性質[44],是影響土壤線蟲群落的另一重要因素[27,45,46]。冗余分析結果表明,土壤全鉀、碳氮比、全磷和有效鉀是影響線蟲群落組成的重要因素(圖5),這些養分的變化可以通過植物的生物量和土壤pH間接影響線蟲群落組成[47]。有研究表明青藏高原地區適當使用鉀肥可以促進植物對氮的吸收,提高植物的生物量和質量[48—49]的同時可能間接增加線蟲密度。本研究也發現線蟲密度與有效鉀顯著正相關(表2),焦加國等[50]也有類似發現。線蟲類群數與土壤碳氮比呈顯著負相關(表2),本研究燕麥樣地的碳氮比下降(表1),有利于凋落物降解,資源可用性的增加減輕了種間競爭的強度,從而可供更多樣的線蟲生存[51]。此外,與天然草地和多年生牧草人工草地不同,一年生燕麥每年需要翻耕重新播種,翻耕會降低土壤容重,增加土壤孔隙和通氣性,可能有利于線蟲的運動和生存[52]。
3.2 人工草地間土壤線蟲群落差異
本研究發現,燕麥(一年生禾本科)人工草地的線蟲總密度以及食真菌、食細菌和雜食-捕食性線蟲密度分別顯著高于其他幾種多年生禾本科和豆科紫花苜蓿人工草地(圖1—2),表明同一功能群的不同植物物種對土壤線蟲群落的影響差異很大,很多研究也有類似發現[16,26]。其次,多年生禾本科牧草和豆科紫花苜蓿人工草地間的線蟲群落密度、多樣性以及植食性、食真菌和食細菌線蟲密度均無顯著差異(圖1—2)。上述研究結果表明,與植物功能群相比,植物物種對土壤線蟲群落密度和多樣性的影響更大。然而,其他研究發現豆科固氮植物通過提高土壤肥力和凈初級生產力,導致其土壤總微生物、細菌、真菌和綠藻生物量以及植食性、食細菌和食真菌線蟲密度顯著高于禾本科植物[27,53]。本研究結果與其相反,可能因為隨著紫花苜蓿種植年限的增加,酚酸類和皂苷類等自毒物質的積累[54—55]抑制了線蟲的增長[56—57]。也可能因為施用的無機氮過量,限制了豆科植物共生根瘤菌的形成及相應的共生固氮作用[58—59],從而對線蟲群落產生了抑制作用。此外,豆科和禾本科植物的根系結構不同,豆科牧草為直根系,根系深;而禾本科牧草為須根系,根系淺,分布更廣,單位體積根系生物量和根系密度更高,且比表面積更大[60—61],為土壤線蟲提供了更多的食物資源和避難所,從而促進其增長[62]。
已有研究發現植食性線蟲的取食作用通過促進根系的碳滲出會增加土壤微生物活性[63],本研究燕麥地較多的植食性線蟲通過促進微生物群落,可能間接增加食細菌和食真菌線蟲的密度。叢枝菌根真菌是食真菌線蟲的食物之一,在草本植物群落中普遍存在[22],燕麥的叢枝菌根真菌孢子密度大于早熟禾、紫羊茅和紫花苜蓿[64],可能是燕麥樣地的食真菌線蟲較多的另一個原因。此外,雜食性線蟲通常以多種食物資源為食,捕食性線蟲以土壤微小型動物為食,燕麥地提供了較多的植食性、食細菌和食真菌線蟲等作為雜食-捕食性線蟲的食物資源,自下而上的調控作用可能會增加燕麥地雜食-捕食性線蟲的密度。
3.3 人工草地對土壤食物網結構和功能的影響
土壤線蟲群落是良好的指示生物,可以指示和反映土壤食物網結構和功能的變化[12]。本研究發現建植人工草地增加了自由生活線蟲成熟度指數(圖3),表明禾本科牧草,尤其是早熟禾和羊茅樣地線蟲群落K對策者比例較高。可能因為禾本科人工草地的土壤有機碳和可利用養分等食物資源降低(表1),導致典型機會主義線蟲(c-p1)類群減少。本研究發現多年生禾本科人工草地降低了線蟲的基礎指數(圖3),表明其土壤食物網的抵抗力降低。線蟲通道指數結果(圖3)表明垂穗披堿草樣地與其他草地不同,以細菌分解通道為主[11]。可能因為該樣地的土壤碳氮比最低(表1),致使食細菌線蟲增加,進而導致分解通道發生轉變[65]。燕麥為一年生牧草,每年進行翻耕處理,其干擾較多年生牧草大,且其線蟲密度高,養分循環快,可能是導致其富集指數和結構指數較低的原因。紫花苜蓿樣地的線蟲富集指數和結構指數最低(圖4),可能是紫花苜蓿產生的酚酸類和皂苷類等次生代謝物引起的自毒效應[54—55]抑制了線蟲的增長[56—57],從而造成了土壤食物網的相對退化。這也是其紫花苜蓿下的植物寄生線蟲成熟度指數PPI最低的原因。同時,可能由于施用的無機氮過量,限制了豆科的固氮作用[58—59],從而可能降低土壤養分。因此,為維持土壤的生態功能,不同植物物種人工草地的施肥策略需進一步研究。
4 結論
綜上所述,青藏高原地區建植燕麥人工草地顯著增加了土壤線蟲群落的總密度和多樣性以及植食性和雜食-捕食性線蟲密度。與植物功能群相比,植物物種對土壤線蟲群落密度和多樣性的影響更大。天然草地轉變為人工草地改變了土壤線蟲的群落組成,其中土壤全鉀、碳氮比、全磷和有效鉀是影響土壤線蟲群落組成的重要因素。建植人工草地是緩解青藏高原草蓄矛盾的重要措施,鑒于當地生態環境脆弱,在大面積建植人工草地前應對地下生態過程的變化深入研究。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
