灰黃青霉D-756實時熒光定量PCR體系的建立及驗證
2022-03-13 09:12:10嚴莉洪石燁祺吳君暢瑛施碧紅
福建農(nóng)業(yè)科技 2022年1期
嚴莉洪 石燁祺 吳君 暢瑛 施碧紅


摘 要:灰黃青霉D-756是1株灰黃霉素高產(chǎn)突變菌株,從轉(zhuǎn)錄水平揭示其灰黃霉素合成關(guān)鍵基因的表達情況,有助于了解其基因功能進而對菌株進行定向遺傳改造。利用Primer3和Primer-BLAST設(shè)計引物,經(jīng)qPCR驗證,引物的擴增產(chǎn)物單一且擴增效率在90%~110%范圍內(nèi),通過geNorm軟件對TUBB、HISH3、RPS24和28S rRNA 4個候選內(nèi)參基因的表達穩(wěn)定性進行評價,選擇評價結(jié)果中較為穩(wěn)定的候選基因TUBB和RPS24作為內(nèi)參基因,構(gòu)建了一個適用于灰黃青霉D-756的qPCR體系。并用該體系檢測灰黃青霉D-756中灰黃霉素合成關(guān)鍵基因gsfA在不同培養(yǎng)時間下的相對表達情況,為下一步研究灰黃霉素合成基因簇中各基因功能提供一個有力的工具。
關(guān)鍵詞:灰黃青霉;灰黃霉素;熒光定量PCR;內(nèi)參基因;聚酮合酶
中圖分類號:Q 933; Q 78? 文獻標志碼:A? 文章編號:0253-2301(2022)01-0006-06
DOI: 10.13651/j.cnki.fjnykj.2022.01.002
Establishment and Validation of the Real-time Fluorescence Quantitative PCR Systemfor Penicillium Griseofulvum D-756
YAN Li-hong, SHI Ye-qi, WU Jun, CHANG Ying, SHI Bi-hong*
(College of Life Science, Fujian Normal University, Fuzhou, Fujian 350108, China)
Abstract: Penicillium griseofulvum D-756 was a mutant strain with high yield of griseofulvin. Revealing the expression of key genes in the biosynthesis of griseofulvin at the transcriptional level would be helpful to understand its gene function and to carry out the directional genetic modification of the strain. The primers were designed by using Primer3 and Primer BLAST. After the qPCR verification, the amplified products of the primers were single and the amplification efficiency was in the range of 90%-110%. Then, the expression stability of the four candidate reference genes TUBB, HISH3, RPS24 and 28S rRNA was evaluated by using the geNorm software. The candidate genes TUBB and RPS24, which were relatively stable in the evaluation results, were selected as the reference genes to construct a qPCR system suitable for Penicillium griseofulvum D-756. Last, this system was used to detect the relative expression of gsfA, the key gene for the biosynthesis of griseofulvin in Penicillium griseofulvum D-756, at different culture times, thus to provide a powerful tool for the further study of each gene function in the biosynthetic gene cluster of griseofulvin.
Key words: Penicillium griseofulvum; Griseofulvin; Fluorescent quantitative PCR; Internal reference gene; Polyketide synthase
灰黃霉素是一種聚酮類抗真菌化合物,主要由青霉屬等真菌產(chǎn)生,于1939年由Oxford等分離得到。灰黃霉素能強烈抑制真菌細胞有絲分裂,干擾真菌DNA合成,抑制絲狀真菌的生長。它對多種皮膚真菌,如小孢子癬菌屬、毛發(fā)癬菌屬和表皮癬菌屬均表現(xiàn)出良好的抗菌活性,并且對人低毒[1],2017年被世界衛(wèi)生組織納入基本藥物清單,用于口服治療皮膚癬菌病[2]。在農(nóng)業(yè)上也被用作防治作物真菌性病害,如葉斑病、霜霉病、水稻稻瘟病等[3]。近年來,隨著對灰黃霉素抗菌機制的逐步深入研究發(fā)現(xiàn),它對丙型肝炎病毒具有一定的抑制作用[4],且具有潛在的抗癌作用[5]。
20世紀70年代吳松剛等[6]以展青霉Penicillium patulum 4541(系灰黃青霉Penicillium griseofulvum的異名)作為出發(fā)菌,利用UV+LiCl等復合誘變處理10多輪獲得了灰黃青霉D-756突變株。灰黃青霉D-756菌株具有耐氯且高產(chǎn)灰黃霉素的優(yōu)良生產(chǎn)特性,發(fā)酵液中檢測灰黃霉素含量是其出發(fā)菌株4541的100多倍,但迄今尚未有對其灰黃霉素高產(chǎn)遺傳背景的報道。隨著測序技術(shù)和實時熒光定量PCR技術(shù)的迅速發(fā)展,通過分析基因在轉(zhuǎn)錄水平的表達量來研究基因功能逐漸成為一種重要的研究手段。
熒光定量PCR技術(shù)(qPCR)利用實時熒光信號監(jiān)測目的基因的擴增,根據(jù)Cq值與起始模板量的線性數(shù)量關(guān)系計算起始模板的表達量,是一種高靈敏度、高通量、強特異性、操作簡便的基因定量技術(shù)[7]。但正因為其靈敏度高,1個試驗參數(shù)的變化就會引起試驗結(jié)果的改變,從而得到不同的試驗結(jié)論。為了增加試驗的可靠性及可重復性,人們制定了一套統(tǒng)一的標準來描述該試驗-MIQE(The Minimum Information for Publication of Quantitative Real-Time PCR Experiments guidelines)[8]。利用相對定量方法分析目的基因表達水平的一個關(guān)鍵步驟是利用內(nèi)參基因?qū)δ康幕駽q值進行正確的校正和歸一化處理,但目前僅使用單一內(nèi)參基因作為參照已經(jīng)很少被接受了,因為幾乎很難找到某單個基因可以在不同的培養(yǎng)條件或細胞組織中穩(wěn)定表達[9]。在絲狀真菌中常用的內(nèi)參基因有TUBB、GAPDH、18S rRNA、28S rRNA、Calmodulin、TefA/Tef1、Actin、HISH3、RPS24等[10-14]。考慮候選內(nèi)參基因時應選擇具有不同功能的內(nèi)參基因,以降低內(nèi)參基因之間的共調(diào)控作用。
最近本課題組完成了灰黃青霉D-756(P.griseofulvum D-756)的基因組測序,并對灰黃青霉D-756中灰黃霉素合成基因簇進行了分析,發(fā)現(xiàn)該基因簇由9個gsf基因構(gòu)成,包括7個高度保守的gsf基因,即gsfA~gsfF、gsfI,以及2個較不保守的gsf基因(gsfJ、gsfR1)[15]。其中g(shù)sfA編碼聚酮合酶,負責起始灰黃霉素骨架的合成,是灰黃霉素合成的關(guān)鍵基因。Cacho等[16]通過基因敲除的手段證實了gsfA基因的缺失會阻斷灰黃霉素的合成。因此,本研究通過設(shè)計熒光定量PCR的引物、篩選穩(wěn)定表達的內(nèi)參基因,構(gòu)建一套適用于灰黃青霉D-756的實時熒光定量PCR體系,并初步探究gsfA基因在不同培養(yǎng)時間下相對表達量的變化,為進一步研究灰黃霉素基因簇各基因功能提供參考依據(jù)。
1 料與方法
1.1 試驗材料
灰黃青霉D-756菌株。
1.2 試驗方法
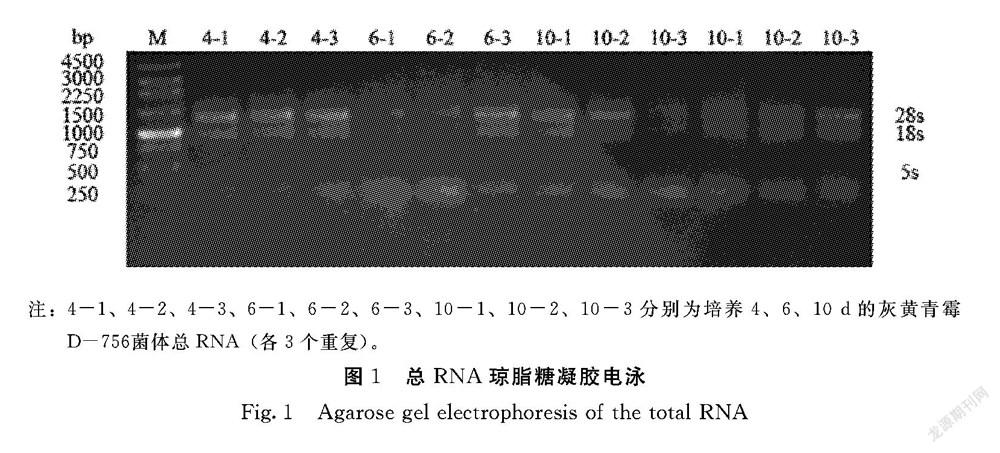
1.2.1 總RNA提取 分別收集適量培養(yǎng)4、6和10 d的灰黃青霉D-756菌絲樣本(每個培養(yǎng)時間設(shè)置3個生物學重復,記為4-1、4-2、4-3;6-1、6-2、6-3;10-1、10-2、10-3),經(jīng)液氮研磨,使用試劑盒提取總RNA(RNA快速提取試劑盒RN40,北京艾德萊),最后進行RNA濃度測定和瓊脂糖凝膠電泳檢測。
1.2.2 RNA反轉(zhuǎn)錄 按照反轉(zhuǎn)錄試劑盒(Takara PrimeScriptTM RT reagent Kit with gDNA Eraser)說明書的操作步驟對提取的總RNA進行反轉(zhuǎn)錄。首先在gDNA Eraser的作用下42℃ 2 min去除RNA樣本中可能含有的基因組DNA,然后將處理過的樣本直接進行15 min的反轉(zhuǎn)錄反應合成cDNA。
1.2.3 引物設(shè)計及合成 選擇4個絲狀真菌中較為常用的候選內(nèi)參基因,分別為TUBB、HISH3、RPS24和28S rRNA,連同目的基因gsfA
,以灰黃青霉D-756中各基因的序列作為模板設(shè)計引物,經(jīng)NCBI-ORF Finder找到序列中最保守且不含內(nèi)含子的區(qū)域,利用Primer-BLAST和Primer3(在線版)設(shè)計qPCR引物(表1),引物由福州鉑尚測序公司進行合成。
1.2.4 引物熔解曲線及標準曲線 將上述設(shè)計好的引物按照熒光定量PCR試劑盒(Takara TB Green Premix Ex TaqTM)說明書中的兩步擴增法以10 μL反應體系進行qPCR,檢測引物的特異性和擴增效率,擴增程序:95℃預變性30 s;95℃ 變性5 s,60℃延伸 30 s,共40個循環(huán);熔解曲線(1個循環(huán)):95℃ 10 s,65℃ 1 min,97℃ 1 s;冷卻(1個循環(huán)):37℃ 30 s。
以50 ng·μL-1 cDNA為起始核酸濃度,按照10倍比稀釋形成5個濃度梯度的cDNA樣本作為模板,qPCR反應體系TB Green Premix Ex Taq 5 μL,正反引物根據(jù)各引物擴增效率添加,ddH2O補足至10 μL,繪制標準曲線,計算擴增效率,選擇擴增效率在90%~110%的引物用于后續(xù)qPCR試驗。
1.2.5 內(nèi)參基因篩選 利用geNorm軟件分析灰黃青霉D-756的4個候選內(nèi)參基因的Cq值。geNorm是Microsoft Excel中自帶的一種應用程式視覺化的Basic腳本(VBA),它可以根據(jù)樣本Cq值自動計算給定樣本中所有內(nèi)參基因的穩(wěn)定性M值(M值與基因表達的穩(wěn)定性成反比),分步排除掉具有最高M值的基因后,剩下的內(nèi)參基因即是樣本中最穩(wěn)定的內(nèi)參基因[17]。候選內(nèi)參基因超過3個時,軟件可通過計算內(nèi)參基因隨機組合的M值,進一步得出內(nèi)參基因的最佳配對數(shù)目及組合。
1.2.6 gsfA基因熒光定量表達分析 gsfA基因相對定量分析按照TB Green試劑盒說明書中要求進行qPCR,后采用2-△△Cq法計算gsfA基因在不同培養(yǎng)時間的相對表達量[18]。
2 結(jié)果與分析
2.1 總RNA完整性檢測
利用1.5%瓊脂糖凝膠電泳檢測灰黃青霉D-756總RNA的完整性,結(jié)果顯示3條清晰的條帶,分別為28S rRNA、18S rRNA和5S rRNA,其中28S rRNA 條帶比18S rRNA條帶亮,5S rRNA條帶較淡,說明總RNA無降解、完整性較好,可用于下一步實時熒光定量PCR(圖1)。
2.2 引物驗證
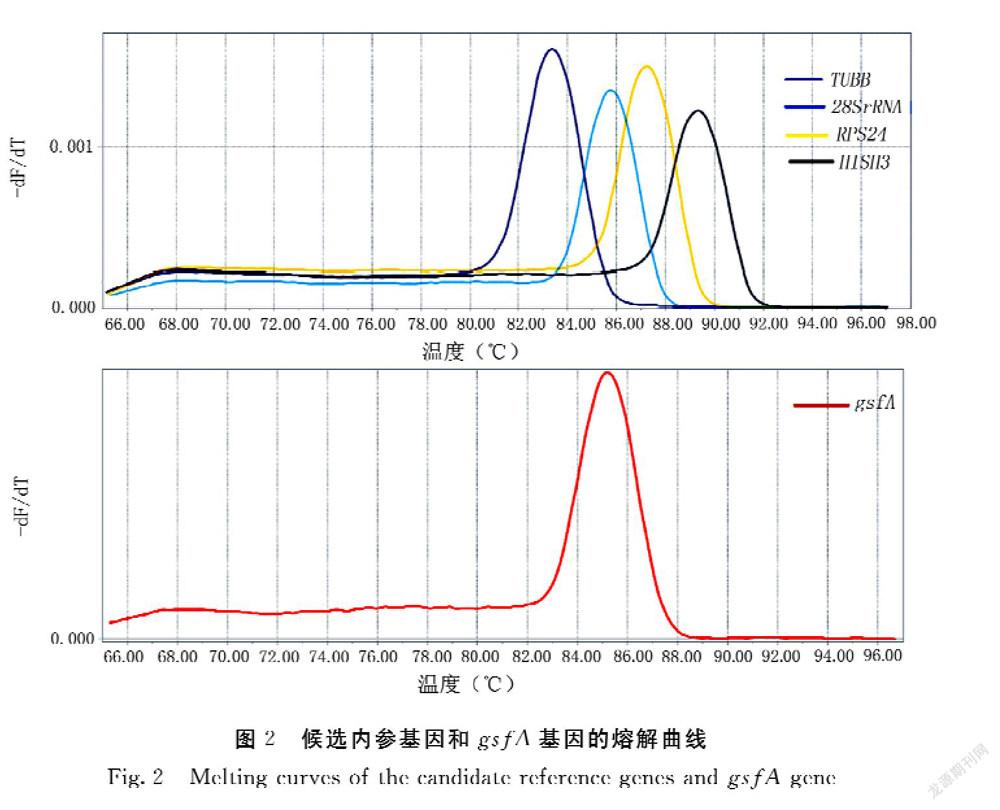
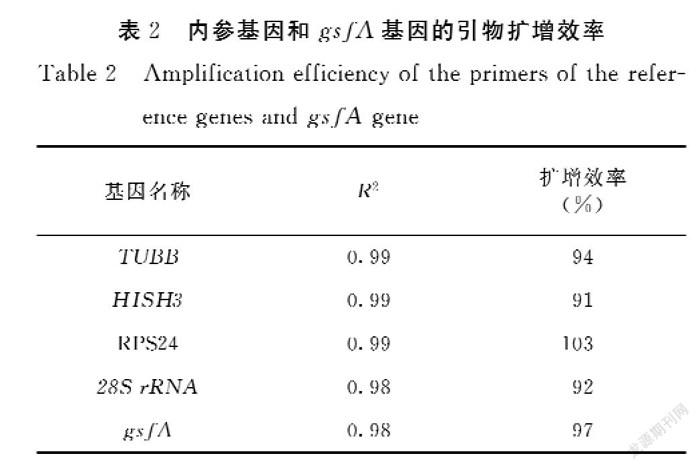
經(jīng)過qPCR擴增獲得引物的熔解曲線,結(jié)果顯示:4個候選內(nèi)參基因和gsfA基因的擴增產(chǎn)物的熔解曲線均只出現(xiàn)單一峰,具有良好的特異性,熔解溫度在80~90℃(圖2)。通過標準曲線進一步確定引物擴增效率,5對引物在0.1~0.3 μmol·L-1濃度范圍內(nèi)的擴增效率為90%~110%,相關(guān)系數(shù)R2為0.998~1,符合引物的要求(表3)。
2.3 內(nèi)參基因穩(wěn)定性分析
一般來說,理想內(nèi)參基因的Cq值在15~30,本試驗利用qPCR對4個候選內(nèi)參基因在培養(yǎng)4、6和10 d時的表達量(Cq值)進行分析,結(jié)果顯示28S rRNA的Cq值低于15,表明其在灰黃青霉D-756中的表達豐度較高,另外3個候選內(nèi)參基因的Cq值均在合理范圍內(nèi)(圖3),可用作下一步內(nèi)參基因穩(wěn)定性分析。
經(jīng)geNorm軟件分析4個候選內(nèi)參基因在培養(yǎng)4、6和10 d時的表達量(Cq值),獲得穩(wěn)定性M值(表3),以穩(wěn)定性M值=1.5為界限,該數(shù)值越接近0說明該內(nèi)參基因的表達越穩(wěn)定。結(jié)果顯示在不同培養(yǎng)時間里4個內(nèi)參基因的穩(wěn)定性M值均小于1.5,表明候選內(nèi)參基因的表達都較穩(wěn)定。由表3可知,在不同培養(yǎng)時間下,M值最小的均為RPS24,說明經(jīng)geNorm軟件分析
RPS24的穩(wěn)定性最好。在培養(yǎng)4 d時,HISH3的M值最大;在培養(yǎng)6、10 d時,28S rRNA的M值最大,說明HISH3和28S rRNA的穩(wěn)定性較差。利用geNorm軟件進一步分析候選內(nèi)參基因的最佳配對數(shù)目及組合,結(jié)果表明選擇RPS24與TUBB
兩個基因作為內(nèi)參基因組合可獲得最佳的穩(wěn)定性,M值為0.011。因此,本試驗從4個候選內(nèi)參基因中篩選得到2個內(nèi)參基因,即RPS24與TUBB組合,后續(xù)以二者Cq值的幾何平均值作為內(nèi)參基因的Cq值計算灰黃青霉D-756中g(shù)sfA基因的相對表達量。
2.4 gsfA基因表達量分析
根據(jù)geNorm軟件分析得出的結(jié)果,選擇RPS24與TUBB組合作為內(nèi)參基因,檢測灰黃青霉D-756中g(shù)sfA基因在搖瓶培養(yǎng)4、6和10 d的相對表達量。單因素方差分析結(jié)果表明,總體上gsfA基因在不同培養(yǎng)時間(4、6、10 d)的相對表達量無顯著差異(F=3.688073,DF=2,P=0.090),其中第4 d與第6 d的相對表達量無顯著差異(T=-1.388, DF=4,P=0.237),第6 d與第10 d的相對表達量差異較大(T=2.357,DF=4,P=0.078)。從趨勢上看,gsfA在第4 d到第6 d的表達水平逐漸升高,并在第6 d時達到最高,平均相對表達量為306.60,而在第6 d到第10 d的表達水平逐漸下降,10 d時平均相對表達量為35.53。
3 結(jié)論與討論
本試驗選擇了4個候選內(nèi)參基因(TUBB、HISH3、RPS24和28S rRNA),除28S rRNA的Cq值小于15之外,其余3個候選內(nèi)參基因的Cq值都在合理范圍。利用geNorm軟件對4個候選內(nèi)參基因在灰黃青霉D-756不同培養(yǎng)時間(4、6和10 d)下的表達量進行分析,結(jié)果表明單個基因表達最穩(wěn)定的是RPS24,TUBB次之,28S rRNA和HISH3的穩(wěn)定性在4個候選內(nèi)參基因中較差。而將RPS24與TUBB組合作為內(nèi)參基因時,它們的穩(wěn)定性更佳,因此最終選擇RPS24與TUBB組合作為灰黃青霉D-756的內(nèi)參基因用于目的基因表達量的標準歸一化。
引物設(shè)計對于實時熒光定量PCR方法來說也是十分重要的。本試驗首先利用NCBI-ORF Finder預測序列中最保守的外顯子序列,以此為模板利用Primer-BLAST和Primer3(在線版)設(shè)計qPCR引物。經(jīng)qPCR的熔解曲線僅出現(xiàn)一個單峰,表明利用該方法設(shè)計的引物特異性良好。進一步通過設(shè)置5個濃度梯度的cDNA樣本作為模板以及在0.1~0.3 μmol·L-1范圍內(nèi)調(diào)整引物濃度制作標準曲線檢測引物的擴增效率,找到了不同引物的最佳濃度,使得擴增效率在90%~110%區(qū)間內(nèi),從而為qPCR的定量提供準確性。
最后,將構(gòu)建的qPCR體系用于檢測灰黃青霉D-756的gsfA基因在本試驗條件下培養(yǎng)4、6和10 d的表達情況。根據(jù)CHOOI等[19]闡述的灰黃霉素生物合成途徑,聚酮合酶GsfA起始灰黃霉素的合成,即一個乙酰輔酶A和六個丙二酸單酰輔酶A起始單元在gsfA基因編碼的非還原聚酮合酶作用下生成灰黃霉素骨架-二苯甲酮,之后在灰黃霉素合成基因簇中其他酶的作用下生成終產(chǎn)物灰黃霉素。在灰黃青霉D-756菌株中,gsfA基因的表達情況與其近緣菌株灰黃青霉PG3的結(jié)果相似[14],在整個培養(yǎng)期間(4~10 d),gsfA基因的相對表達量無顯著性差異。培養(yǎng)4~6 d時,D-756菌株gsfA基因的相對表達量保持較高水平(合成骨架需要),培養(yǎng)10 d時,D-756菌株中g(shù)sfA基因的表達水平出現(xiàn)下降,這與gsfA基因在灰黃霉素合成過程中的作用基本一致。本試驗為后續(xù)開展灰黃青霉D-756灰黃霉素生物合成基因簇各基因表達的分析打下了基礎(chǔ)。
參考文獻:
[1]HAMDY A K, SHEHA M M, ABDEL-HAFEZ A A, et al.Design, Synthesis, and Cytotoxicity Evaluation of Novel Griseofulvin Analogues with Improved Water Solubility[J].International Journal of Medicinal Chemistry,2017,2017:7386125.DOI:10.1155/2017/7386125.
[2]WHO Expert Committee on Selection, Use of Essential Medicines, World Health Organization.The Selection and Use of Essential Medicines: Report of the WHO Expert Committee, 2017(including the 20th WHO Model List of Essential Medicines and the 6th Model List of Essential Medicines for Children)[M].Geneva,World Health Organization, 2017.
[3]張傳能,黃銘杰,毛寧.灰黃霉素對水稻稻瘟病菌的防治效果研究[J].中國農(nóng)學通報, 2015, 31(4):190-194.
[4]JIN H, YAMASHITA A, MAEKAWA S, et al.Griseofulvin, an oral antifungal agent, suppresses hepatitis C virus replication in vitro[J].Hepatol Res,2008, 38(9): 909-918.
[5]REBACZ B, LARSEN T O, CLAUSEN M H, et al.Identification of griseofulvin as an inhibitor of centrosomal clustering in a phenotype-based screen[J].Cancer Research, 2007, 67(13): 6342-6350.
[6]吳松剛,郝家驥,戚曉玉,等.Penicillum patulum 4541耐前體突變株的選育[J].遺傳, 1980, 2(4): 16-18.
[7]BUSTIN S A, BENES V, NOLAN T, et al.Quantitative real-time RT-PCR-a perspective[J].J Molecular Endocrinology, 2005, 34(3): 597-601.
[8]BUSTIN S A, BENES V, GARSON J A, et al.The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments[J].Cliniacl Chemistry, 2009, 55(4): 611-622.
[9]VANDESOMPELE J.Reference Gene Validation Software for Improved Normalization[J].Real-time PCR: current technology and applications, 2009, (4): 47-64.
[10]BARKER B K K, VDISCH M, MAZURIE A, et al.Transcriptomic and proteomic analyses of the Aspergillus fumigatus hypoxia response using an oxygen-controlled fermenter[J].BMC Genomics, 2012, (13): 62.DOI: 10.1186/1471-2164-13-62.
[11]ARCHER M, XU J.Current Practices for Reference Gene Selection in RT-qPCR of Aspergillus: Outlook and Recommendations for the Future[J].Genes (Basel), 2021, 12(7): 960.DOI: 10.3390/genes12070960.
[12]ZAMPIERI D, NORA L C, BASSO V, et al.Validation of reference genes in Penicillium echinulatum to enable gene expression study using real-time quantitative RT-PCR [J].Current Genetics, 2014, 60(3): 231-236.
[13]LEVIN E, RAPHAEL G, MA J, et al.Identification and Functional Analysis of NLP-Encoding Genes from the Postharvest Pathogen Penicillium expansum[J].Microorganisms, 2019, 7(6): 175.DOI: 10.3390/microorganisms7060175.
[14]BANANI H, MARCET-HOUBEN M, BALLESTER A R, et al.Genome sequencing and secondary metabolism of the postharvest pathogen Penicillium griseofulvum[J].BMC Genomics, 2016, (17): 19.DOI: 10.1186/s12864-015-2347-x.
[15]PARISA A, LIHONG Y, YULONG W,et al.Conservation of griseofulvin genes in the gsf gene cluster among fungal genomes[J].G3(Genes Genomes Genetics), 2021.jkab399.DOI:10.1093/g3journal/jkab399.
[16]CACHO R A, CHOOI Y H, ZHOU H, et al.Complexity generation in fungal polyketide biosynthesis: a spirocycle-forming P450 in the concise pathway to the antifungal drug griseofulvin[J].ACS Chemical Biology, 2013, 8(10): 2322-2330.
[17]VANDESOMPELE J, DE PRETER K, PATTYN F, et al.Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes[J].Genome Biology,2002, 3(7): RESEARCH0034.DOI: 10.1186/gb-2002-3-7-research0034.
[18]LIVAK K J, SCHMITTGEN T D.Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta DeltaC(T)) Method[J].Methods, 2001, 25(4):402-408.
[19]CHOOI Y H, CACHO R, TANG Y.Identification of the viridicatumtoxin and griseofulvin gene clusters from Penicillium aethiopicum[J].Chemical Biology, 2010, 17(5):483-494.
(責任編輯:柯文輝)
收稿日期:2021-12-10
作者簡介:嚴莉洪,女,1995年生,碩士研究生,主要從事真菌分子遺傳學研究。
通信作者:施碧紅,女,1968年生,博士,教授,主要從事微生物生化與分子生物學等研究(E-mail:shibh@fjnu.edu.cn)。
基金項目:國家自然科學基金(91435106);2022年福建師范大學生命科學學院“溪源江學者”科研創(chuàng)新項目(22FSSK015)。
3044501908206
