東海原甲藻對羊棲菜合子生長及光合活性的化感作用研究
2022-03-15 07:45:42王彩霞秦文莉陳斌斌馬增嶺
水生生物學報 2022年2期
王彩霞 賈 陽 秦文莉 陳斌斌 王 敏 馬增嶺
(1. 溫州大學浙江省亞熱帶水環境與海洋生物資源保護重點實驗室, 溫州 325035; 2. 溫州大學城鎮水污染生態治理技術國家地方聯合工程研究中心, 溫州 325035)
近年來赤潮的發生機理、生態危害及控制策略成為自然科學的研究熱點, 赤潮暴發經常會引起養殖魚類和貝類的大量死亡, 進而導致近岸水生生態系統的失衡[1,2]。一些能夠引發赤潮的藻種會產生毒素并通過食物鏈的富集作用, 對人類健康產生威脅。過去幾十年, 全球已報告了數千起因食用赤潮發生海域海產品引起的中毒事件, 且呈增加態勢[3]。
中國海洋環境質量公報的統計結果顯示, 我國東海海域每年赤潮暴發次數最多、面積最大, 而東海原甲藻 (Prorocentrum donghaiense) 是最主要的赤潮原因種。東海原甲藻赤潮影響面積大(>1000—10000 km2), 持續時間長(>30d), 從1990年開始幾乎每年春季都有暴發[4]。研究顯示, 東海原甲藻赤潮會引起褶皺臂尾輪蟲 (Brachionus plicatilis)和蒙古裸腹溞(Moina mongolica) 存活率下降[5], 降低中華哲水蚤(Calanus sinicus)的產卵量和孵化率, 進而導致浮游動物種群數量的下降[6]。
大型海藻場主要位于潮間帶下區和潮下帶數米淺水區硬相海底, 而人工栽培的大型海藻則主要掛養在近岸水域。近年來近岸海域赤潮頻發正在引起大型海藻場的快速退化及養殖品系的損傷: 如2002年5月末在韓國南部海域發生的赤潮異彎藻(Heterosigma akashiwo)赤潮, 其消退時產生的大量黏液物質(Mucilaginous material)在大葉藻(Zostera marina)構成的海藻場上形成了0.3—1.0 m厚的黏液層, 從而使得該區域海藻場消失殆盡[7,8]; 自20世紀80年代以來經常在日本瀨戶內海Harima-Nada區域發生的浮動彎角藻 (Eucampia zodiacus)赤潮常常引起該地區養殖的紫菜葉狀體漂白, 嚴重影響紫菜質量, 并造成數千萬美元的損失[9,10]; 而發生在南非西海岸、由高濃度非產毒的海洋原甲藻(P. micans)、新角藻(Neoceratium furca)及少量產毒的鏈狀亞歷山大藻(Alexandrium catenella)和漸尖鰭藻(Dinophysis acuminate)形成的“黑潮”, 在消退過程中因赤潮藻細胞腐敗形成的近岸低氧、高硫化氫環境引起了潮間帶95%的生物個體死亡, 并導致當地由紫菜和石莼形成的海藻場全部消失[11]。而海藻場一旦消失, 要靠自然恢復就變得非常困難, 即使環境條件改善也要經歷數年時間[11,12]。
浙江溫州海域是我國經濟海藻羊棲菜的主產區, 其產量約占國內羊棲菜總產量的90%, 產品遠銷日本、韓國; 與此同時, 該海域東海原甲藻赤潮頻發, 羊棲菜養殖戶深受其擾。羊棲菜富含多糖、膳食纖維素、B族維生素、褐藻酸、甘露醇及人體必需的礦物質和微量元素, 具有重要的食用和藥用價值, 在日本被稱為“長壽菜”。其具有性繁殖和無性繁殖2種生活史階段: 孢子體成熟時進行有性繁殖; 之后枝葉即爛去, 而假根繼續保存, 待環境適合生長后, 自假根處再分化出新的幼孢子體[13]。每年的4月中旬至5月中旬羊棲菜孢子體開始成熟, 進入生產采收期; 而采收時也要預留一部分長勢良好、豐產特征明顯的成熟孢子體作為種苗, 用于采收后至6月中旬的人工育苗。羊棲菜有性繁殖過程中具有精卵同步釋放、同步受精, 合子固著、幼苗發育快及植株營養生長快速等特點[14—17]。在羊棲菜有性繁殖時, 卵子和精子分別由雌雄生殖托同步釋放,在精卵結合后, 合子24h后進入幼胚生長發育階段;幼胚經自然海區人工放養1.5個月左右, 相繼生長發育成具有三葉體形態特征的幼孢子體(即幼苗), 至8月份幼苗長至10 cm以上時開始夾苗掛養。而每年的4—8月份正是我國東海近海赤潮高發期。其在成熟期、有性生殖期及幼苗期均可能與有害赤潮發生遭遇戰, 這種重要生物學階段與赤潮高發期的高度重合使其成為研究有害赤潮對大型海藻抑制效應及其機制的理想材料。
因為大型海藻人工養殖能夠改善水體富營養化水平并降低赤潮發生風險, 因此其生態效應及經濟價值備受關注。然而, 抑制作用也是相互的[18],赤潮作為密集海水養殖區內常見的生態事件[19], 有關大型海藻的生理活動如何應答甲藻赤潮脅迫卻很少受到關注, 尤其缺乏赤潮發生過程中, 大型海藻合子及幼苗在生長發育、光合特性和產物累積等方面對不同赤潮生物如何應答的研究。在實踐中也觀察到東海原甲藻赤潮侵襲羊棲菜養殖筏的現象[20]。其次已有研究表明, 米氏凱倫藻會顯著抑制羊棲菜孢子體氣囊及生殖托的光合活性[18], 并且其對羊棲菜合子的生長及光合作用的抑制作用具有濃度依賴性[20]。我們前期的研究發現東海原甲藻也會顯著抑制羊棲菜合子的生長和光合作用[21],為了進一步探究其化感機理, 本文研究了濃度為1.00×105cells/mL東海原甲藻活細胞懸浮液(LC)、細胞破碎液(RC)和無細胞濾液(FC)對羊棲菜合子生長及光合特性影響, 并利用快速葉綠素熒光動力學曲線參數分析(JIP-test)解析了東海原甲藻化感作用對羊棲菜合子光系統的損傷位點。以期為準確、客觀地評價東海原甲藻赤潮頻發對羊棲菜栽培的危害提供理論依據和科學數據。
1 材料與方法
1.1 材料和培養條件
東海原甲藻由自然資源部第二海洋研究所的陸斗定教授提供, 用滅菌的F/2培養基[22]進行擴增培養, 培養溫度為22℃, 光強為100 μmol photons/(m2·s), 光暗比為 12h∶12h。培養液每天人工搖動2—3次。取培養至對數生長期的細胞懸浮液用于實驗。羊棲菜雌雄孢子體于2019年5月中旬從洞頭水產科學技術研究所的羊棲菜培育基地獲得, 這一時期羊棲菜的大部分生殖托已經成熟并且開始釋放生殖細胞。雌雄植株用過濾的自然海水清洗3次,然后在充氣(0.5 L/min)條件下培養, 其他培養條件與上同。培養 2—3d后, 取雌雄生殖托按照5∶1的數量比例在同一個塑料水槽中培養, 雌雄托分別釋放生殖細胞并在水體內完成受精; 用 280 目尼龍網收集合子, 沖洗干凈后重新懸浮于裝有過濾海水的燒杯中; 最后將合子均勻接種到6孔細菌培養板中, 培養板海水深度約為0.5 cm, 在培養箱中放置2d, 待合子通過絲狀假根附著于細菌培養板底部后進行共培養實驗。
1.2 東海原甲藻活細胞懸浮液(LC)、細胞破碎液(RC)和無細胞濾液(FC)的制備
東海原甲藻細胞懸浮液(Living Cell Suspension, LC)的制備: 取處于指數生長期的東海原甲藻培養液, 在3000×g、 4℃下離心15min后收集細胞,重新懸浮于新鮮的F/2培養基中, 使其細胞濃度達到1.00×105cells/mL。我們前期的研究已經證明此濃度的東海原甲藻會抑制羊棲菜合子的生長及光合作用。東海原甲藻無細胞濾液(Cell-free Supernatant, FC)的制備: 取濃度為1.00×105cells/mL的東海原甲藻細胞培養液, 經0.45 μm醋酸纖維素薄膜過濾得到東海原甲藻的無細胞濾液。東海原甲藻細胞破碎液(Ruptured Cell Suspension, RC)的制備:取濃度為1.00×105cells/mL的東海原甲藻細胞培養液50 mL, 在3000×g、 4℃下離心 15min, 然后將收集的藻細胞在50 mL F/2培養基中重懸, 用超聲細胞破碎儀(JX-650)在冰浴條件下進行破碎。所有的活細胞懸浮液, 細胞破碎液及無細胞濾液均為現配現用。
1.3 共培養實驗
研究一種微藻對其共存生物的化感效應, 主要通過微藻細胞懸浮液、細胞濾液、細胞破碎液或微藻中某些次生代謝物質(化感物質)與其他生物共培養[23]。因此本文采用新鮮F/2培養基培養的羊棲菜合子作為對照組, 采用利用LC、FC及RC培養的羊棲菜合子作為處理組, 在半連續培養(每2天更換1次培養液)條件下進行為期10d的共培養實驗, 在各個共培養組中羊棲菜合子的密度均為35個/cm2。
1.4 羊棲菜合子生長發育的測定
用ECLIPSETs2 倒置顯微鏡觀察各組中羊棲菜合子的形態, 并且每隔1天用數碼顯微相機(NikonDSRi2)對各個組別的羊棲菜合子在同一位位置進行拍照記錄。然后用NIS-Elements D5.10 軟件統計羊棲菜合子的長度和寬度。由于合子的形態近似于圓柱體, 故其大小根據圓柱體體積公式計算。合子的相對生長速率(RGR)用如下公式計算[24], 其中V1和V2分別為t1和t2天時合子的體積:
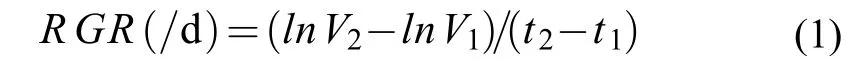
1.5 羊棲菜合子葉綠素a含量的測定
在不同條件下培養10d后, 將各處理組的羊棲菜合子從6孔細菌培養板上刮下, 用0.45 μm醋酸纖維素薄膜過濾后的自然海水沖洗, 至鏡檢未發現東海原甲藻細胞, 用差量法稱取一定量的羊棲菜合子,各個組別的羊棲菜合子均用5 mL的無水甲醇在4℃條件下過夜提取, 然后用離心機在 5000×g離心10min, 用紫外可見分光光度計(UV530; Beckman Coulter, USA)測定上清液的吸光值。葉綠素a濃度的計算方式如下[25]:
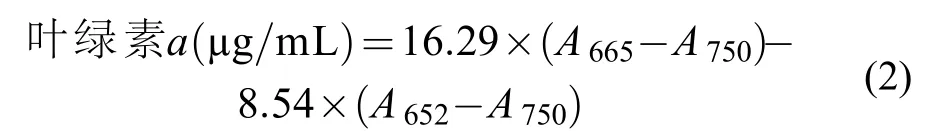
式中,A652、A665和A750分別表示上清液在波長為652、665和750 nm 處的吸光度值。
1.6 羊棲菜合子光合活性的測定
羊棲菜合子的最大光化學效率(Fv/Fm)和相對電子傳遞速率(rETR)采用多激發波長葉綠素熒光儀(Heinz Walz GmbH, Effeltrich, Germany)測定。測定前將羊棲菜合子重懸于3 mL的過濾海水中, 暗適應15min。而后在梯度光照(PAR)水平下測定rETR-I曲線, 每個光強梯度間隔20s。rETR-I曲線各光合參數的計算參照根據 Eilers和Peeters[26]的公式:
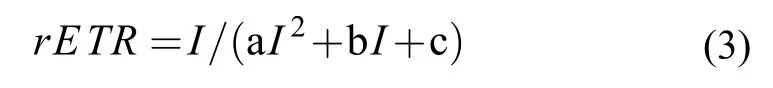
式中, a、b、c為校準參數。表觀光合作用效率(α)、最大相對電子傳遞速率(rETRmax)和半飽和光強(Ik)用參數a、b和 c表示為:
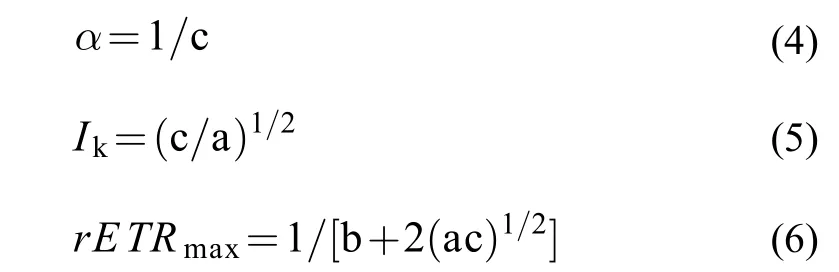
1.7 快速葉綠素熒光誘導動力學曲線分析(JIPtest)
羊棲菜合子暗適應15min后, 通過多激發波長葉綠素熒光儀(Heinz Walz GmbH, Effeltrich, Germany)測其快速葉綠素熒光動力學曲線(OJIPcurves)。快速葉綠素熒光誘導動力學曲線有O、J、I和P等相, 在該熒光曲線上O點是熒光值最小的點, P點是熒光值最大的點[27], 這些點反應了光合生物進行光合作用時PSⅡ中光量子在傳遞過程中的狀態, 該傳遞過程可以簡單地描述為質體醌QA的還原過程, 過程如下: 首先O點是快速光曲線熒光最低點, 該點熒光值為Fo, 在該點時PSⅡ電子傳遞受體則處于氧化態; 熒光曲線上升至J點和I點, 該過程是光化學反應階段, 此時質體醌QA接受電子, 被還原為和Q2A-, 電子也傳遞到次級醌受體QB; 當QB從接受電子, 形成Q2B-時, QA完全進入還原狀態,此時PSⅡ反應中心完全關閉, 熒光曲線也上升到P點, 即熒光值達到最高(Fm)[28]。另外為了方便比較不同樣品的快速光曲線及相關熒光參數, 原始OJIP熒光曲線可通過下列公式標準化為相對可變熒光:
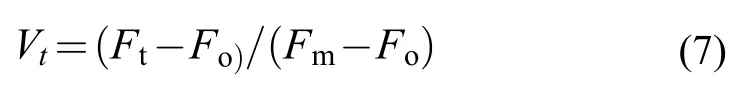
快速熒光曲線包含大量關于PSII反應中心原初光化學反應的信息, 通過對曲線熒光參數的分析,可以反映環境脅迫對羊棲菜合子產生的影響。本研究主要通過以下參數對合子的OJIP曲線進行分析:VJ是在J點的相對可變熒光, 代表照光2ms時活性反應中心關閉的程度;Mo是OJIP曲線的初始斜率, 代表QA被還原的最大速率,VJ和Mo分別通過以下公式計算:
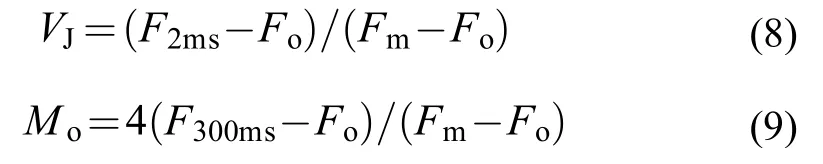
式中,F2ms和F300ms代表熒光曲線在2和300ms 時的熒光值。用以下幾個參數來代表量子產額或能量分配比率: 最大光量子產率(φpo=Fv/Fm);ψo代表了在反應中心捕獲的激子中用來推動電子傳遞到電子傳遞鏈中超過 QA的其他電子受體的激子占用來推動 QA還原激子的比率, 即照光2ms 時有活性的反應中心的開放程度;φEo, 代表反應中心吸收的光能用于電子傳遞的量子產額, 即反應中心吸收的光能將電子傳遞到電子傳遞鏈中超過 QA的其他電子受體的概率。以上參數的計算方式分別如下:
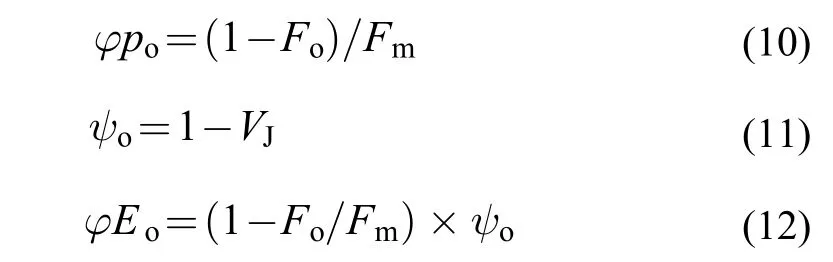
以下幾個比活性參數表示羊棲菜合子單位PSⅡ反應中心的活性: 單位反應中心吸收的光能(ABS/RC), 單位反應中心捕獲的用于還原QA的能量(TRo/RC), 單位反應中心捕獲的用于電子傳遞的能量(ETo/RC), 單位反應中心熱耗散的能量(DIo/RC)表達式分別如下:
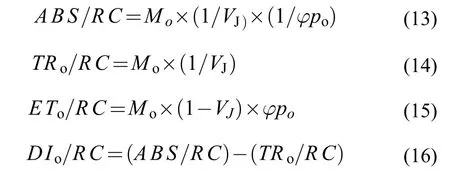
性能指數(Performance Index,PI)能夠較好地反映從激子到最后還原PSⅠ和最終電子受體這個過程的能量變化, 是應激情況下光合生物總體活力的指標[29]。PI由3個參數構成: PSⅡ反應中心的密度(RC/ABS), 光子捕獲性能(PTR), 被捕光子激發電子傳遞的性能(PET), 表達式分別如下[30]:
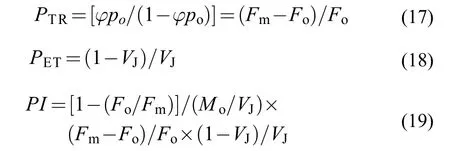
1.8 統計分析
所有的測定結果表示為平均值±標準誤(n≥3),數據采用 Excel 和 SPSS 進行整理和統計分析, 使用 Origin9.0(OriginLab, USA)作圖。用單因素方差分析(One-way ANOVA, Duncan)進行不同處理間的差異性分析, 差異顯著水平為P=0.05。
2 結果
2.1 東海原甲藻對羊棲菜合子發育的影響
受精后, 羊棲菜合子會進行持續的減數分裂,經1—2h相繼發育至64倍體或128倍體, 進入幼胚生長發育階段, 并在24h分生出透明的絲狀假根, 將合子固著于基質上。圖1展示了合子(幼胚)在10d中形態的變化, 在不同共培養方式下, 合子均由最初的近橢圓形沿長軸逐漸伸長并發育成柱狀體。在培養6d后, 各處理組的合子形態保持正常, LC、RC和 FC處理組的合子在培養 6d后體積開始明顯小于對照組(圖1)。此外, 在10d培養結束時各組合子大小依次為: 對照組>RC組>LC組>FC組, 各組合子的平均體積分別為: 31.83×10-3、22.55×10-3、19.94×10-3和18.17×10-3mm3(圖2A), 對應的相對生長速率分別為0.20 、0.13、0.12和0.12/d(圖2B)。與對照組相比, LC、RC和FC處理組合子的體積分別下降了37.37%、29.15%和 42.93% (圖2A), 相對生長速率分別下降了 41.23%、34.63%和42.02%(圖2B)。從合子體積和相對生長速率的變化可以看出, LC、RC和FC處理對合子的生長皆有顯著的(P<0.05)抑制作用(圖2), 其中FC及LC處理對合子抑制程度接近且均大于RC組。
2.2 東海原甲藻對羊棲菜合子色素含量的影響
在培養10d后, 在對照組、LC、RC和FC處理組中合子的葉綠素a的含量分別為 579.60、501.82、562.91和262.15 μg/g 鮮重(圖3), 在3種處理方式下培養的合子的色素含量均顯著(P<0.05)低于對照組(圖3)。與對照組相比, 在LC、RC和FC處理組合子的葉綠素a含量分別降低了13.42%、2.88%和54.77%, 對羊棲菜合子葉綠素a的抑制強度依次為FC>LC>RC。
2.3 東海原甲藻對羊棲菜合子光合活性的影響
圖4展示了不同處理方式下培養10d后羊棲菜合子的快速光曲線(RLCs, 圖4A), 及由該曲線得到的表觀光合作用效率(α, 圖4B)和最大相對電子傳遞速率(rETRmax, 圖4C)。對照組、LC處理組、RC處理組和FC處理組羊棲菜合子的α分別為0.22、0.20、0.23和 0.20 electrons/photon; 對應的最大相對電子傳遞速率分別為139.44、107.18、131.30和63.36 μmol electrons/(m2·s)。與對照組相比,LC和FC處理下合子的表觀光合作用效率分別降低了9.13%和7.10%, 而RC處理下則提高了5.88%;LC、RC和FC處理下合子的最大相對電子傳遞速率分別降低了23.14%、5.84%和54.57%; 即各處理組對羊棲菜合子最大相對電子傳遞速率和表觀光合作用效率的抑制強度依次為FC>LC>RC。
2.4 東海原甲藻對羊棲菜合子快速熒光動力學曲線(OJIP)的影響
東海原甲藻處理10d后羊棲菜合子的初始快速熒光動力學曲線以對數形式展示在圖5A中, 從該圖可以看出3種處理下羊棲菜合子的整體熒光值均小于對照組。為了方便各組間進行比較, 將初始熒光曲線進行標準化, 得到相對可變熒光曲線(圖5B),展示從50μs到1s之間各組熒光的變化情況。從圖5B中可以看出各處理組中各點的相對可變熒光(Vt)均大于對照組, 說明處理組中合子的電子傳遞受阻。
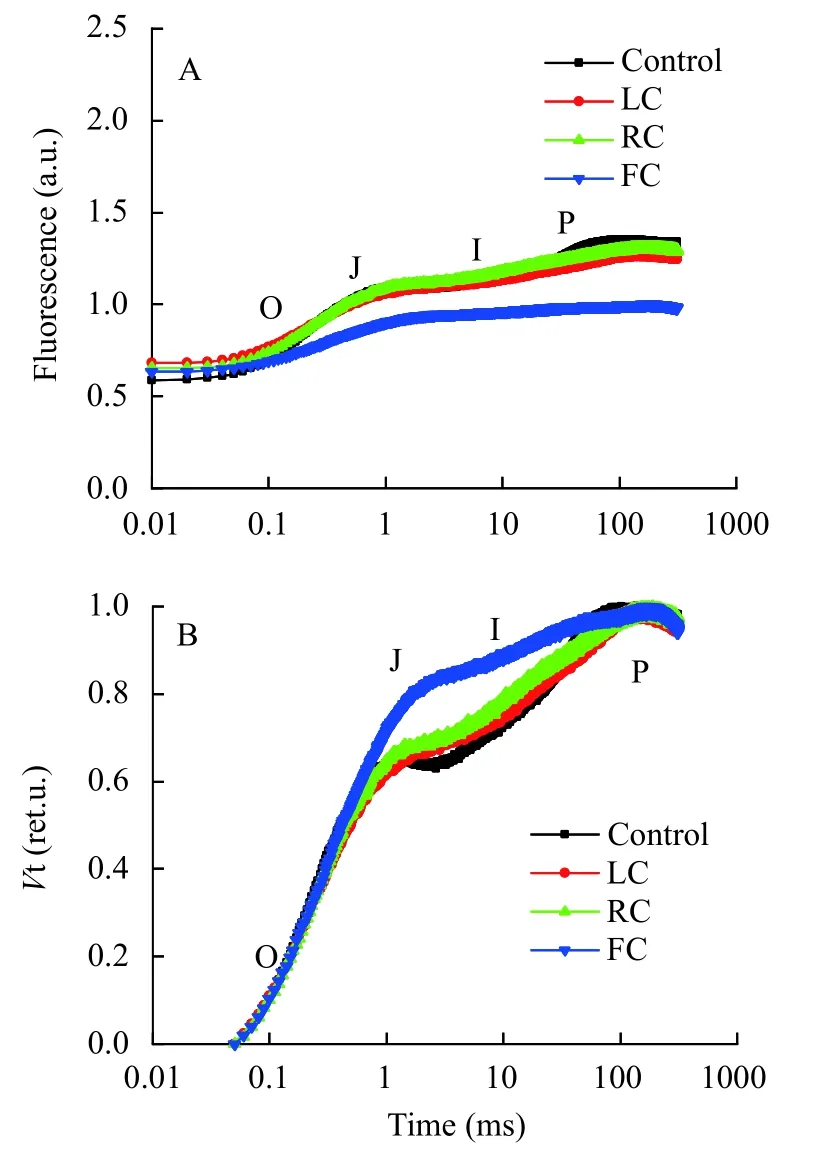
圖5 東海原甲藻處理10d后羊棲菜合子的初始快速熒光動力學曲線(OJIP; A)和標準化的快速熒光動力學曲線(B)Fig. 5 Effects of P. donghaiense treatments on the original (A)and normalized OJIP curves (B) of S. fusiforme zygotes after 10 days treatments with P. donghaiense
2.5 JIP-test分析結果
在圖6展示的是與PSⅡ受體側量子產率相關的部分熒光參數。與對照組相比, FC處理組的VJ值提高了29.49%; 各處理組的Mo值沒有顯著差異;LC及FC處理組的φPo均顯著低于對照組, 分別降低了17.24%和35.81%; 與對照組相比, FC處理組的φo值顯著低于對照組, 降低了48.57%; FC處理組的φEo顯著低于對照組, 降低了64.89%。
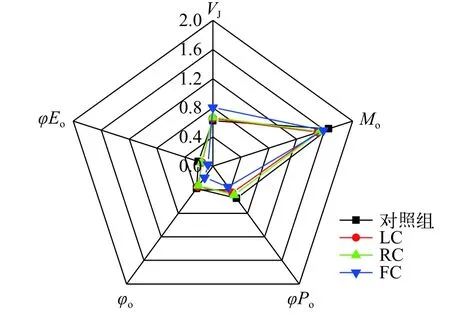
圖6 由東海原甲藻處理10d后羊棲菜合子的快速光曲線得到的與能量分配相關參數 (VJ, Mo, φPo, ψo, φEo)分析Fig. 6 Analysis of selected JIP-test parameters (VJ, Mo, φPo, ψo,φEo) derived from the chlorophyll fluorescence transients of S.fusiforme zygotes after 10 days treatments with P. donghaiense
圖7展示了東海原甲藻處理10d后羊棲菜合子的幾組比光合活性參數。參數ABS/RC表示反應中心的表面積, FC處理組的ABS/RC值與對照組相比顯著(P<0.05)提高, 增加了16.02%。TRo/RC值反映了反應中心的活性, 該值在LC、RC和FC處理組中均顯著(P<0.05)降低, 與對照組相比分別降低了13.86%、14.15%和26.20%。與反應中心電子傳遞相關的參數ETo/RC值在FC處理組中顯著(P<0.05)上升, 與對照組相比提高了71.36%。與熱耗散相關的DIo/RC值在各處理組中也呈現出上升趨勢, 其中在FC處理組中顯著(P<0.05)提高, 增加了66.89%(圖7)。
圖8展示了東海原甲藻處理10d后羊棲菜合子光合性能指數(PI)及其各參數的變化情況。從圖8A可以看出, 不同處理下羊棲菜合子的PI值均低于對照組(P<0.05), 其中FC處理組降低了76.89%。此外,與對照組相比, 與光合作用光反應階段相關的PTR值在LC、RC和FC處理組中均顯著降低(P<0.05),分別降低了31.08%、19.56%和54.84%。PET及RC/ABS值在各組中沒有顯著差異(P>0.05)。從圖8B中還可以看出, 對于LC、RC和FC處理組中PI值的降低, 暗反應部分(PET值)的貢獻率最大。
3 討論
當前研究結果表明, 東海原甲藻的活細胞懸浮液, 細胞破碎液及無細胞濾液均會對羊棲菜合子的生長、色素合成及光合活性產生抑制作用, 但是不同處理方式對羊棲菜合子的抑制程度有差異。化感作用在生物相互作用之間的重要性已廣為人知,已有報道很多微藻會通過化感作用抑制其他共存藻類的生長甚至導致細胞裂解死亡[31]。在本研究中, 東海原甲藻活細胞懸浮液處理組及無細胞濾液處理組中羊棲菜合子的生長(圖1和圖2) 及光合活性(圖4—8)均被顯著抑制, 細胞破碎液處理組中羊棲菜合子葉綠素a的含量(圖3)與對照組相比變化不顯著。這表明東海原甲藻細胞分泌到培養液中的化感物質能夠對羊棲菜合子的生長及光合作用產生抑制作用。Poulin等[32]的研究表明短凱倫藻(Karenia brevis)分泌到共培養液中的化感物質能夠破壞競爭對象細胞膜的完整性及光合作用過程進而對競爭者的生長產生抑制作用。此外, Shang等[20]的研究結果表明羊棲菜合子的生長和光合活性可以被米氏凱倫藻(K. mikimotoi)分泌的化感活性物質所抑制, 并且該抑制作用與米氏凱倫藻細胞濃度呈正相關。
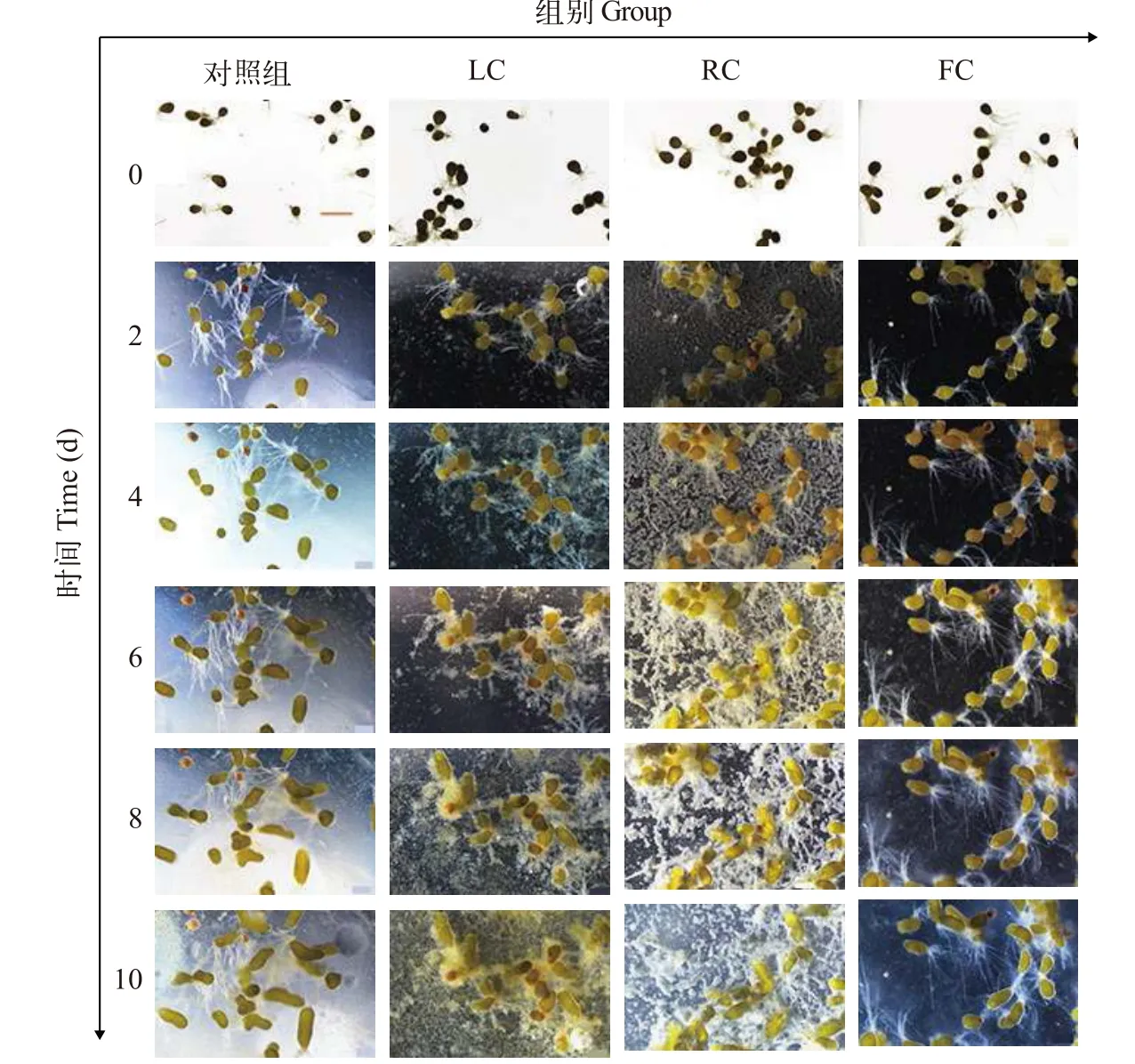
圖1 東海原甲藻對羊棲菜合子形態的影響Fig. 1 Effects of P. donghaiense treatments on morphology of S. fusiforme zygotes
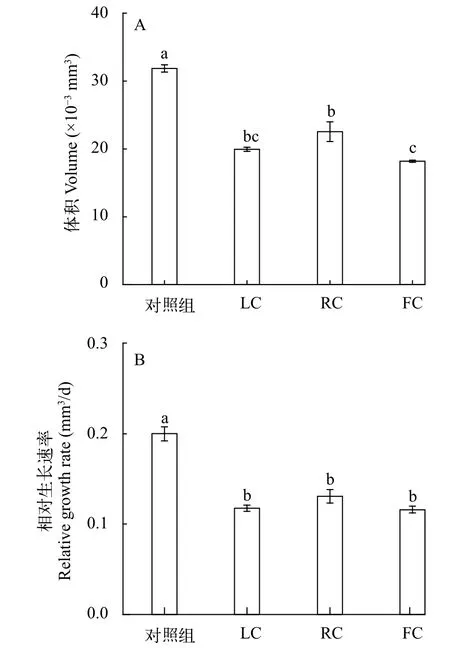
圖2 東海原甲藻處理10d后羊棲菜合子的體積(A)和相對生長速率(B)的變化Fig. 2 Effects of P. donghaiense treatment on volume (A) and relative growth rate (B) of Sargassum fusiforme zygotes after 10 days’ treatments with P. donghaiense
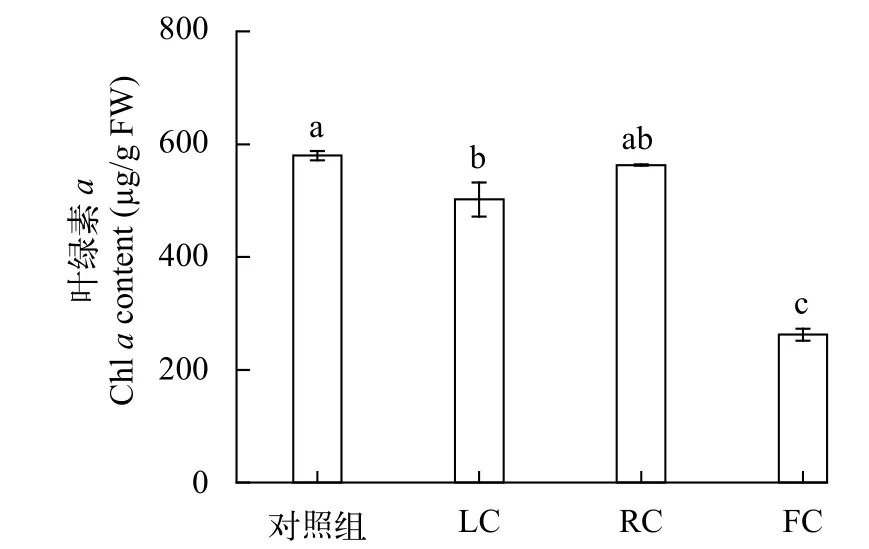
圖3 東海原甲藻處理10d后羊棲菜合子葉綠素a含量的變化Fig. 3 The effect of P. donghaiense treatments on chlorophyll a contents of Sargassum fusiforme zygotes after 10 days treatments with P. donghaiense
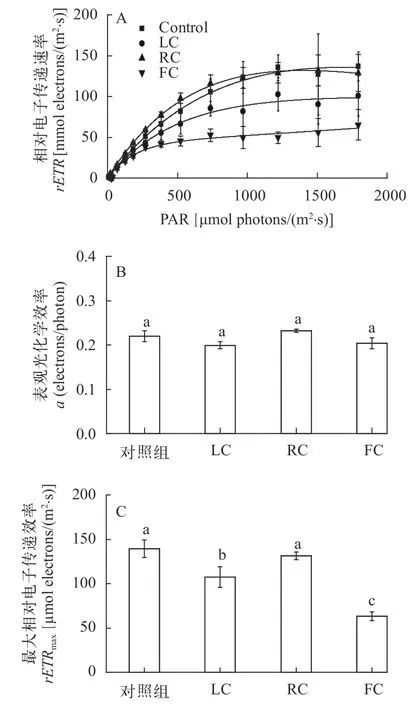
圖4 東海原甲藻處理10d后羊棲菜合子的光合作用(P)-光強(I)響應曲線(A)、表觀光化學效率(B)和最大相對電子傳遞速率(C)Fig. 4 Effects of P. donghaiense treatments on the rETR (P)versus irradiance (I) curves (A), the apparent photosynthetic efficiency (α; B) and the maximal electron transport rate (rETRmax;C) and of Sargassum fusiforme zygotes after 10 days treatments with P. donghaiense
細胞破碎液處理組由于細胞破碎會導致細胞內容物的大量釋放, 本應包含更多的化感物質, 從而導致羊棲菜合子受到更大的抑制, 但是該處理組中羊棲菜合子在生長及光合活性方面受到的抑制程度卻沒有活細胞懸浮液處理組及無細胞濾液處理組大。這應該是由于東海原甲藻細胞破碎使細胞中化感物質釋放的同時, 藻細胞中能夠促進羊棲菜合子生長的大量營養物質也釋放出來, 從而在很大程度上抵消了化感物質的抑制作用[20]。已有研究表明東海原甲藻細胞中包含苯丙氨酸、賴氨酸和組氨酸等豐富的營養物質, 且這些物質可以刺激其他微藻的生長[33]。此外, 大型海藻對營養物質具有很強的同化能力, 在沿海地區常用來降低水體的富營養化[34]。因此, 細胞破碎液處理組中羊棲菜合子在受到化感物質抑制的同時也能夠吸收東海原甲藻破碎而釋放的營養物質用于生長。如Doblin等[35]的研究顯示鏈狀裸甲藻的生長可以被低分子量的有機物質促進, 并提出鏈狀裸甲藻對這些物質的吸收可能是由于有機物質在細胞表面細菌和細胞外肽酶的作用下降解從而被細胞吸收。
已有研究針對石莼(Ulva lactuca)[36]及強壯前溝藻(Amphidinium carterae)[37]的光合特性進行JIPtest分析, 結果表明利用JIP-test分析能夠很好地反映PSII反應中心的活性狀態[38]。從對羊棲菜合子葉綠素a快速熒光動力學曲線的JIP-test分析結果來看, 東海原甲藻活細胞懸浮液、細胞破碎液及無細胞濾液處理組羊棲菜合子的最大光化學產量(Fm)值均顯著降低, 表明東海原甲藻分泌的化感物質能夠抑制羊棲菜合子的最大量子產率(圖6)。從抑制效果來看, 無細胞濾液處理組及活細胞懸浮液處理組的抑制程度大于細胞破碎液處理組, 這與各處理組對羊棲菜合子生長及色素含量的抑制效果一致。此外, Shang等[20]的研究顯示羊棲菜合子的Fm可以被米氏凱倫藻分泌的化感物質所抑制, 而米氏凱倫藻和東海原甲藻都是在我國東海長江口廣泛分布的有害赤潮種。
在某些脅迫條件下, PSⅡ反應中心會部分甚至全部失活, 從而導致單個活性反應中心吸收的光能增加, 但大部分被捕獲光能不能用來推動電子傳遞,而是轉化為熱能耗散掉[28,39]。在本研究中, 無細胞濾液處理組、活細胞懸浮液處理組及細胞破碎液處理組中羊棲菜合子的ABS/RC及DIo/RC參數比對照組顯著升高, 而參數TRo/RC顯著降低(圖7B), 該結果表明東海原甲藻處理導致了羊棲菜合子PSⅡ部分反應中心失活, 并且用于電子傳遞將QA還原為Q-A的能量減少, 從而使大部分能量轉化為熱能耗散掉。這證明了東海原甲藻所分泌的化感物質可以通過使羊棲菜合子活性反應中心失活從而對其光合作用產生抑制作用。
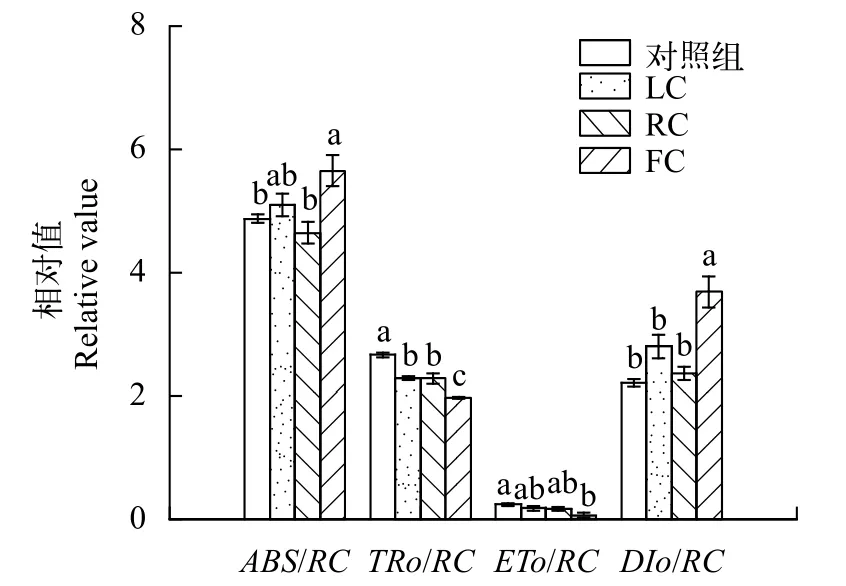
圖7 東海原甲藻處理10d后羊棲菜合子每PSⅡ反應中心(/RC)的能量通量變化Fig. 7 Fast chlorophyll transients energy flux ratios per RC of S.fusiforme zygotes after 10 days treatments with P. donghaiense
光合性能參數PI包含的3個參數(PTR、PET和RC/ABS), 可以反映光合生物光合機構的狀態[40]。其中捕光性能參數PTR的降低表示RCs產生了持續性損傷進而導致RCs的失活[41]。在本研究中, 與對照組相比, 各處理組的PI值、PET值及PTR均出現了降低(圖8A), 其中無細胞濾液處理組的PI值顯著降低(P<0.05), 而PET對PI降低的貢獻最大(圖8B), 該結果表明東海原甲藻抑制了羊棲菜合子的光合活性, 而主要的抑制作用發生在PSⅡ的暗反應階段。此外,PTR值的降低也表明東海原甲藻對羊棲菜合子PSⅡ活性反應中心具有抑制作用。
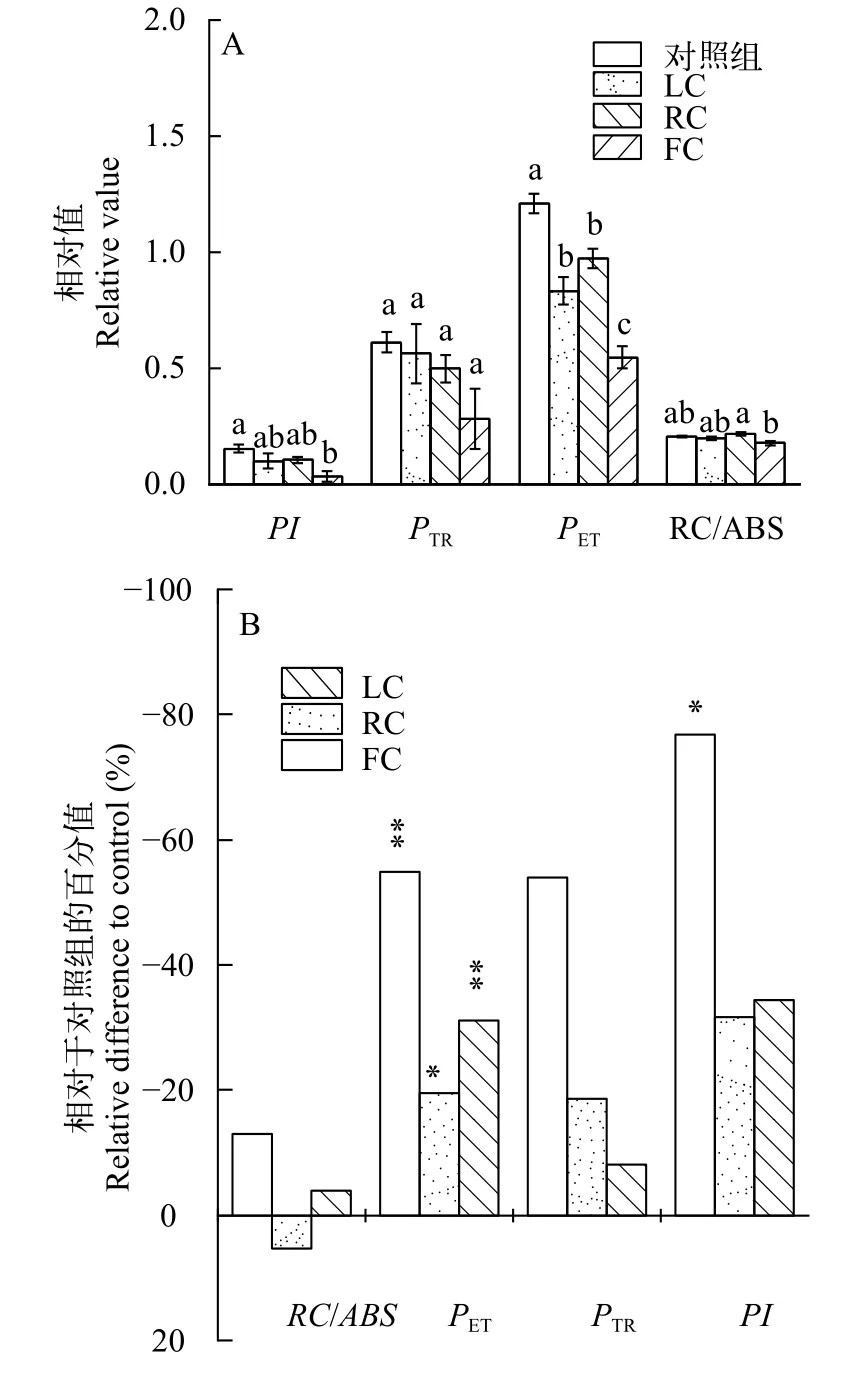
圖8 東海原甲藻處理10d后羊棲菜合子光合性能指數(PI)及其參數(PTR、PET 和RC/ABS)變化(A)及各性能指數相對于對照組降低的比率(B)Fig. 8 Changes in PI and its components of Sargassum fusiforme zygotes after 10 days treatments with Prorocentrum donghaiense(A), and the ratios relative to the mono-cultured ones (B)
4 結論
細胞密度為1.00×105cells/mL下的東海原甲藻活細胞懸浮液、細胞破碎液和無細胞濾液均能顯著抑制羊棲菜合子的生長、光合作用及葉綠素a的合成, 其中無細胞濾液對羊棲菜合子的抑制作用最強, 其次是活細胞懸浮液, 再次是細胞破碎液。這表明東海原甲藻能夠通過向細胞外釋放化感物質對羊棲菜合子產生抑制作用, 而細胞破碎液中可能含有某些能夠刺激羊棲菜合子生長的物質, 從而在一定程度上抵消了化感物質的抑制作用。JIPtest分析結果表明東海原甲藻分泌的化感物質主要抑制了PSⅡ中的電子傳遞, 即通過抑制被捕光子激發電子傳遞的效率(PET)來抑制羊棲菜合子的光合作用。此外, 化感物質還能引起PSⅡ的部分RCs失活, 從而使大量的激發能轉化為熱能耗散掉, 進一步降低了羊棲菜合子的光合活性。
