運(yùn)動(dòng)疲勞狀態(tài)下大鼠腦皮層單羧酸轉(zhuǎn)運(yùn)蛋白表達(dá)的變化及意義
2022-03-16 03:41:44李玉榮裴文娟
關(guān)鍵詞:檢測(cè)
高 晨,王 菀,李玉榮,裴文娟
(聯(lián)勤保障部隊(duì)第九四〇醫(yī)院全科醫(yī)學(xué)科,蘭州 730050)
疲勞發(fā)生在外周肌肉和中樞神經(jīng)兩個(gè)部位,決定后者的關(guān)鍵因素是腦能量代謝失衡,從而難以維持中樞神經(jīng)系統(tǒng)持續(xù)向外周發(fā)放神經(jīng)沖動(dòng)[1],其機(jī)制與運(yùn)動(dòng)缺氧導(dǎo)致靜息狀態(tài)下腦電活動(dòng)減弱并反饋性抑制中樞神經(jīng)系統(tǒng)和脊髓前角α運(yùn)動(dòng)神經(jīng)元活動(dòng)有關(guān)[2-4]。研究表明,極量運(yùn)動(dòng)或缺氧狀態(tài)下,無(wú)氧代謝所產(chǎn)生的乳酸能夠被大腦攝取并作為能量底物為神經(jīng)元供能[5],且乳酸優(yōu)先于葡萄糖被神經(jīng)元利用氧化供能[6]。這一機(jī)制在劇烈活動(dòng)時(shí)對(duì)于維持突觸傳遞具有重要作用。然而,另有研究表明,缺血缺氧或腦損傷后乳酸堆積卻通過(guò)降低pH值而引發(fā)一系列病理反應(yīng),從而損傷神經(jīng)元[7-8]。機(jī)體乳酸代謝轉(zhuǎn)運(yùn)的載體是廣泛分布于組織細(xì)胞膜上的單羧酸轉(zhuǎn)運(yùn)蛋白(monocarboxylate transporters,MCTs),其可分為14種亞型[9]。其中,MCT1主要在微血管內(nèi)皮細(xì)胞、室管膜細(xì)胞、星形膠質(zhì)細(xì)胞和少突膠質(zhì)細(xì)胞中表達(dá),MCT2主要在神經(jīng)元中表達(dá),而MCT4幾乎只在星形膠質(zhì)細(xì)胞中表達(dá)[10]。研究證實(shí),血乳酸難以通過(guò)血腦屏障,星形膠質(zhì)細(xì)胞是腦乳酸的主要來(lái)源[11]。廣泛表達(dá)于星形膠質(zhì)細(xì)胞和神經(jīng)元的MCT1、MCT2和MCT4在中樞乳酸的產(chǎn)生和利用中起到至關(guān)重要的作用[12-13]。據(jù)此推測(cè),運(yùn)動(dòng)疲勞過(guò)程中,正是由于MCTs的存在,腦乳酸在氧化供能和腦損傷之間維系了一種平衡狀態(tài)。目前關(guān)于運(yùn)動(dòng)疲勞狀態(tài)下腦內(nèi)MCTs表達(dá)的變化尚無(wú)明確結(jié)論。本研究通過(guò)檢測(cè)運(yùn)動(dòng)疲勞模型動(dòng)物的運(yùn)動(dòng)力竭時(shí)間變化以及大腦皮層運(yùn)動(dòng)區(qū)MCT1、MCT2和MCT4的表達(dá),初步探討運(yùn)動(dòng)耐力變化與腦乳酸代謝轉(zhuǎn)運(yùn)之間的相關(guān)性,為運(yùn)動(dòng)疲勞的醫(yī)學(xué)干預(yù)奠定理論基礎(chǔ)。
1 材料與方法
1.1 實(shí)驗(yàn)動(dòng)物及分組
8周齡清潔級(jí)雄性SD大鼠42只,體質(zhì)量(280±10)g,由聯(lián)勤保障部隊(duì)第九四〇醫(yī)院動(dòng)物實(shí)驗(yàn)科提供[SCXK(軍)2017-0023]。大鼠常規(guī)分籠飼養(yǎng)于屏障設(shè)施[SYXK(軍)2017-0047],正常飲水、飲食。環(huán)境溫度維持在18~24℃,相對(duì)濕度40%~60%。動(dòng)物實(shí)驗(yàn)經(jīng)聯(lián)勤保障部隊(duì)第九四〇醫(yī)院倫理委員會(huì)審核同意(編號(hào)2020KYLL032)。大鼠隨機(jī)分為7組,每組6只。對(duì)照組(Control)正常飼養(yǎng),不運(yùn)動(dòng);其余6組為疲勞1~6組(Fatigue 1-6)分別進(jìn)行力竭運(yùn)動(dòng)1、3、5、7、9、14 d,以達(dá)到運(yùn)動(dòng)疲勞狀態(tài)。
1.2 實(shí)驗(yàn)設(shè)備及試劑
ZH-PT型動(dòng)物跑臺(tái)為安徽淮北正華生物儀器設(shè)備有限公司產(chǎn)品。MCT1(貨號(hào)PAB16059)和MCT4(貨號(hào)PAB21410)多克隆抗體均購(gòu)自亞諾法(Abnova)生技股份有限公司,MCT2(貨號(hào)SC-50323)購(gòu)自美國(guó)Santa公司,膠質(zhì)纖維酸性蛋白(glial fibrillary acidic protein,GFAP)(貨號(hào)3670)單克隆抗體購(gòu)自美國(guó)Cell Signaling Technology公司,神經(jīng)元核抗原(neuronal nuclei antigen,NeuN)(貨號(hào)MAB377)單克隆抗體購(gòu)自美國(guó)Millipore公司。熒光單克隆抗體購(gòu)自北京康為世紀(jì)生物科技有限公司。熒光顯微鏡(型號(hào)AX80)購(gòu)自日本Olympus株式會(huì)社。
1.3 運(yùn)動(dòng)疲勞模型建立
采用Bedford等[14]的遞增負(fù)荷運(yùn)動(dòng)方案建立大鼠運(yùn)動(dòng)疲勞模型。疲勞組大鼠先行在動(dòng)物跑臺(tái)上進(jìn)行3 d適應(yīng)性訓(xùn)練(速度為15 m∕min,時(shí)間為5 min∕d,坡度0°),之后分別進(jìn)行為期1、3、5、7、9和14 d的遞增負(fù)荷運(yùn)動(dòng)(速度為8.2 m∕min×15 min+15 m∕min×15 min+20 m∕min持續(xù),直至力竭,1次∕日)。力竭標(biāo)準(zhǔn):大鼠跑步姿態(tài)由蹬地式變?yōu)榉厥剑瑴襞艿篮?∕3處達(dá)3次以上,各種刺激驅(qū)趕無(wú)效,且停跑后表現(xiàn)為呼吸急促、神情倦怠、反應(yīng)遲鈍[15]。分別記錄各組大鼠負(fù)荷運(yùn)動(dòng)最后1 d達(dá)到力竭狀態(tài)的運(yùn)動(dòng)時(shí)間,取平均值。
1.4 蛋白質(zhì)印跡檢測(cè)
不同時(shí)間疲勞模型建立后,各組分別取3只大鼠在異氟烷麻醉下斷頭處死。完整取出腦組織,嗅球至小腦蚓部前后冠狀均分為5等份,第2和3等份最表層主要為運(yùn)動(dòng)區(qū)皮層。取運(yùn)動(dòng)區(qū)皮層組織,勻漿后裂解,提取總蛋白,BCA法測(cè)定濃度。然后行聚丙烯酰胺凝膠電泳,每孔上樣蛋白總量為20μg,電泳時(shí)間約120 min。電泳結(jié)束后以350 mA恒流帶電轉(zhuǎn)膜(120 min),5%牛血清白蛋白室溫振蕩封閉1 h。5%脫脂奶粉稀釋后,分別加入一抗(包括MCT1、MCT2、MCT4和αtubulin抗體,體積稀釋比例分別為1∶500、1∶1 000、1∶500和1∶1 000),4℃孵育過(guò)夜;加入辣根過(guò)氧化物酶標(biāo)記的二抗(體積稀釋比例為1∶5 000),在室溫下振蕩孵育2 h[16]。滴加發(fā)光檢測(cè)液,在凝膠成像系統(tǒng)中拍照,并采用Image J軟件進(jìn)行灰度量化分析。實(shí)驗(yàn)重復(fù)3次,取平均值。
1.5 大鼠腦切片制備及免疫熒光檢測(cè)
各組剩余3只大鼠在異氟烷麻醉下經(jīng)心臟灌流處死,斷頭完整取出腦組織。用質(zhì)量分?jǐn)?shù)4%的多聚甲醛溶液固定24 h,蔗糖PB溶液階梯脫水48 h,包埋后自嗅球到小腦蚓部前后5等份連續(xù)冠狀位冰凍切片(厚度25μm)。取第2和3等份切片進(jìn)行免疫熒光雙標(biāo)法檢測(cè)。
鼠腦切片用1%牛血清蛋白+0.3%Triton X-100在室溫下封阻非特異性結(jié)合,并破膜1 h。PBS清洗后,加入一抗MCT1(1∶200)+GFAP(1∶500)、MCT2(1∶500)+NeuN(1∶500),或MCT4(1∶200)+GFAP(1∶500)稀釋液,4℃緩慢振蕩孵育過(guò)夜。PBS清洗后,加入1∶100稀釋的熒光二抗異硫氰酸熒光素和異硫氰酸羅丹明,室溫下緩慢振蕩孵育2 h。PBS清洗后,加入1∶2 000稀釋的DAPI,室溫下避光孵育10 min。PBS清洗后,鼠腦切片轉(zhuǎn)移至載玻片上,60%甘油PBS封固,熒光顯微鏡下觀察并進(jìn)行皮層定位圖像采集。FITC激發(fā)∕發(fā)射波長(zhǎng)為480 nm∕535 nm,TRITC激發(fā)∕發(fā)射波長(zhǎng)為535 nm∕610 nm。用Images J軟件進(jìn)行圖像分析。
1.6 統(tǒng)計(jì)學(xué)方法
采用SPSS 13.0統(tǒng)計(jì)軟件包對(duì)本研究數(shù)據(jù)進(jìn)行分析。計(jì)量資料采用±s表示。不同組樣本均數(shù)間比較采用單因素方差分析,并根據(jù)各組總體方差齊同與否,選擇Bonferroni法或Tamhane's T2法修正結(jié)果,以P<0.05為差異有統(tǒng)計(jì)學(xué)意義。
2 結(jié)果
2.1 大鼠力竭運(yùn)動(dòng)時(shí)間變化情況
如表1所示,疲勞1組大鼠平均運(yùn)動(dòng)力竭時(shí)間為(120.17±11.18)min。運(yùn)動(dòng)5 d后疲勞3組的平均運(yùn)動(dòng)力竭時(shí)間達(dá)到最短,為(67.00±7.07)min。此后,大鼠運(yùn)動(dòng)能力逐漸恢復(fù),運(yùn)動(dòng)14 d后疲勞6組力竭時(shí)間恢復(fù)至(91.17±11.43)min。與疲勞1組相比,各組大鼠的平均運(yùn)動(dòng)力竭時(shí)間均明顯縮短,差異均有統(tǒng)計(jì)學(xué)意義(P<0.05)。與力竭時(shí)間最短的疲勞3組相比,疲勞5組和疲勞6組的大鼠力竭時(shí)間明顯延長(zhǎng),表明其運(yùn)動(dòng)能力的恢復(fù)具有統(tǒng)計(jì)學(xué)意義(P<0.05)。
表1 各疲勞組大鼠平均力竭時(shí)間對(duì)比Table 1 The exhaustive time of rats in fatigue groups(±s,n=6)

表1 各疲勞組大鼠平均力竭時(shí)間對(duì)比Table 1 The exhaustive time of rats in fatigue groups(±s,n=6)
注:Fatigue 1-6分別為疲勞1~6組。與Fatigue 1組比較,*P<0.05;與Fatigue 3組比較,#P<0.05;與Fatigue 5組比較,△P<0.05。單因素方差分析采信Tamhane’s T2修正值。
組別Fatigue 1 Fatigue 2 Fatigue 3 Fatigue 4 Fatigue 5 Fatigue 6遞增負(fù)荷運(yùn)動(dòng)天數(shù)/d 135791 4平均運(yùn)動(dòng)力竭時(shí)間/min 120.17±11.18#△81.17±10.57*67.00±7.07*△75.67±4.97*89.17±9.45*#91.17±11.43*#
2 .2大鼠腦運(yùn)動(dòng)區(qū)皮層MCT1、MCT2及MCT4表達(dá)變化情況
用蛋白質(zhì)印跡法檢測(cè)各組大鼠運(yùn)動(dòng)區(qū)皮層MCT1、MCT2及MCT4表達(dá)的變化情況。如圖1所示,Control組大鼠運(yùn)動(dòng)區(qū)皮層MCT1、MCT2及MCT4均少量正常表達(dá)。疲勞負(fù)荷運(yùn)動(dòng)5 d后,F(xiàn)atigue 3組鼠腦運(yùn)動(dòng)區(qū)皮層中MCT2表達(dá)較Control組明顯升高74.2%。疲勞負(fù)荷運(yùn)動(dòng)7 d后,F(xiàn)atigue 4組鼠腦運(yùn)動(dòng)區(qū)皮層中MCT1和MCT4表達(dá)較Control組分別明顯升高約89.5%和92.0%,差異均有統(tǒng)計(jì)學(xué)意義(P<0.05)。此后,F(xiàn)atigue 5組和6組鼠腦運(yùn)動(dòng)區(qū)皮層中MCT1、MCT2和MCT4均維持高表達(dá),與Control組相比差異均有統(tǒng)計(jì)學(xué)意義(P<0.05)。

圖1 蛋白質(zhì)印跡法檢測(cè)各組大鼠腦運(yùn)動(dòng)區(qū)皮層中MCT1、MCT2和MCT4表達(dá)比較Figure1ExpressionofMCT1,MCT2andMCT4incerebralmotorcortexofratsindifferentgroupsdetectedby Westernblotting
進(jìn) 一 步 分 別 利 用MCT1∕GFAP、MCT2∕NeuN或MCT4∕GFAP免疫熒光雙標(biāo)技術(shù)對(duì)比檢測(cè)鼠腦運(yùn)動(dòng)區(qū)皮層膠質(zhì)細(xì)胞中MCT1、MCT2和MCT4的表達(dá)變化情況。如圖2所示,與Control組相比,疲勞負(fù)荷運(yùn)動(dòng)7 d后鼠腦運(yùn)動(dòng)區(qū)皮層膠質(zhì)細(xì)胞中MCT1與MCT4表達(dá)均明顯升高并維持穩(wěn)定;MCT2的表達(dá)也呈現(xiàn)出逐漸增高的趨勢(shì),疲勞負(fù)荷運(yùn)動(dòng)5 d即維持于高表達(dá)狀態(tài)。熒光檢測(cè)結(jié)果與蛋白質(zhì)印跡檢測(cè)結(jié)果相一致。
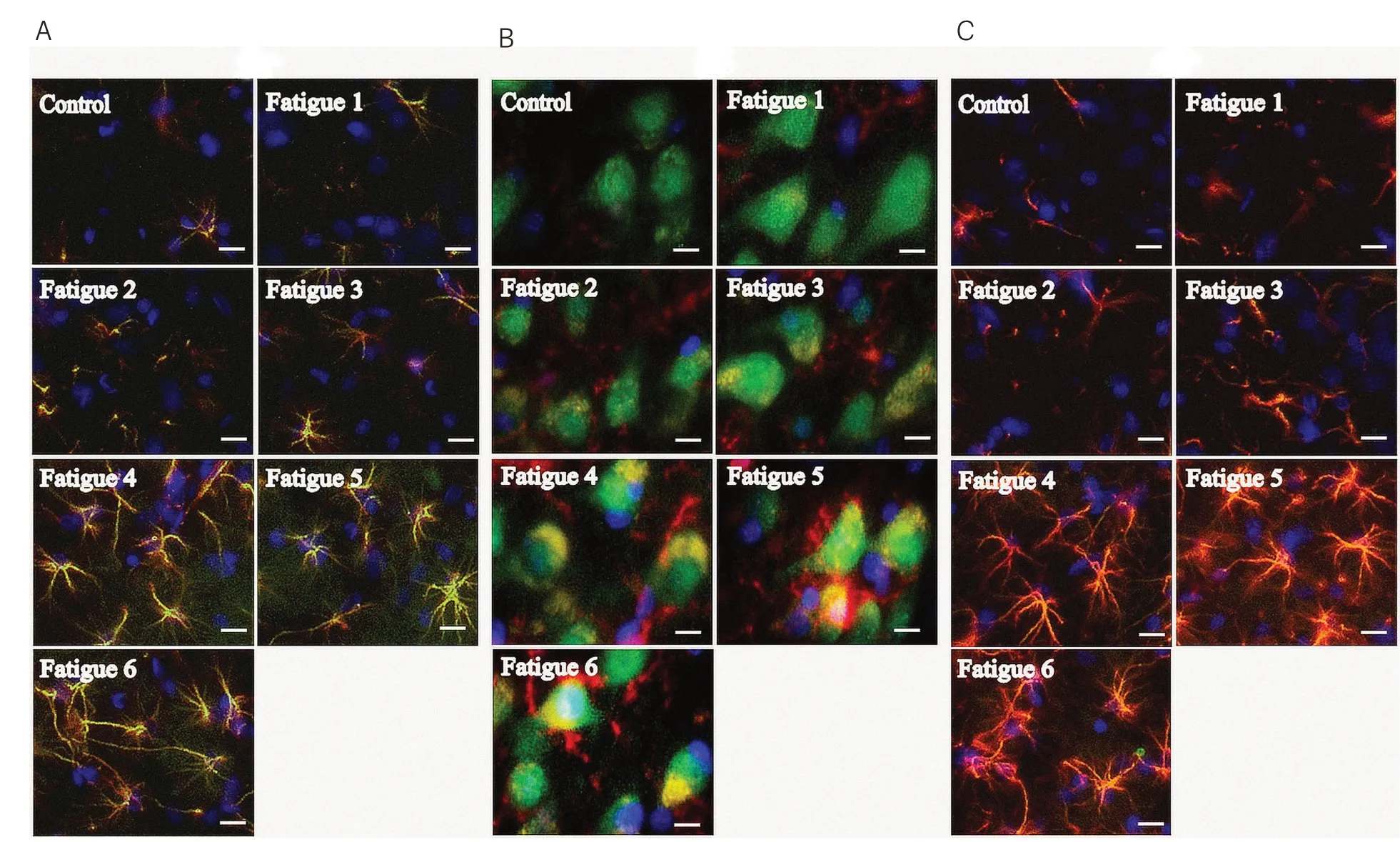
圖2 免疫熒光法檢測(cè)各組大鼠腦運(yùn)動(dòng)區(qū)皮層MCT1、MCT2和MCT4表達(dá)(×400)Figure2ExpressionofMCT1,MCT2andMCT4incerebralmotorcortexofratsindifferentgroupsdetectedby imunofluorescence(×400)
結(jié)果提示每日疲勞負(fù)荷運(yùn)動(dòng)后,大鼠運(yùn)動(dòng)能力迅速下降,在運(yùn)動(dòng)5 d后達(dá)到最低點(diǎn)。此后繼續(xù)運(yùn)動(dòng),大鼠腦內(nèi)MCTs表達(dá)水平逐漸升高,腦乳酸轉(zhuǎn)運(yùn)及代謝被激活,大鼠運(yùn)動(dòng)能力在一定程度上恢復(fù)。這種由重復(fù)運(yùn)動(dòng)誘導(dǎo)的對(duì)運(yùn)動(dòng)疲勞的適應(yīng)性,在運(yùn)動(dòng)1周后維持于相對(duì)平穩(wěn)的狀態(tài)。
3 討論
乳酸(C3H6O3)是糖代謝過(guò)程的中間產(chǎn)物。與通常情況下機(jī)體對(duì)葡萄糖的有氧氧化過(guò)程不同,乳酸的產(chǎn)生來(lái)源于無(wú)氧狀態(tài)下的糖酵解過(guò)程。雖然無(wú)氧糖酵解過(guò)程的能量供應(yīng)效率低于糖的有氧氧化,但對(duì)于劇烈運(yùn)動(dòng)狀態(tài)下的能量代謝維持至關(guān)重要。通常,運(yùn)動(dòng)過(guò)程中,肌肉是產(chǎn)生乳酸最多的部位[17]。大強(qiáng)度運(yùn)動(dòng)時(shí),糖酵解生成的乳酸使細(xì)胞內(nèi)H+濃度上升,pH值降低。機(jī)體對(duì)乳酸的代謝起始于細(xì)胞間乳酸的穿梭,而后進(jìn)一步通過(guò)糖異生等途徑供應(yīng)能量。乳酸的穿梭過(guò)程依賴于MCTs。研究表明,運(yùn)動(dòng)強(qiáng)度的變化是MCTs含量改變的重要因素。擁有14種亞型的MCTs分布于不同的組織細(xì)胞膜表面,其中MCT1、MCT2和MCT4對(duì)運(yùn)動(dòng)狀態(tài)下的乳酸轉(zhuǎn)運(yùn)至關(guān)重要[13]。由于乳酸穿過(guò)細(xì)胞膜進(jìn)行轉(zhuǎn)運(yùn)是一個(gè)主動(dòng)耗能的過(guò)程,因此較大強(qiáng)度的耐力運(yùn)動(dòng)后細(xì)胞膜上與MCTs共同起到協(xié)同轉(zhuǎn)運(yùn)作用的鈉氫交換蛋白(sodium hydrogen exchanger 1,NHE1)的表達(dá)亦相應(yīng)升高,從而利用Na+∕H+ATP酶產(chǎn)生的化學(xué)梯度能量完成整個(gè)轉(zhuǎn)運(yùn)過(guò)程,同時(shí)維持細(xì)胞內(nèi)外pH值的相對(duì)穩(wěn)定。研究表明,這種協(xié)同轉(zhuǎn)運(yùn)是乳酸與H+以1∶1的等摩爾方式相偶聯(lián)的同向轉(zhuǎn)運(yùn),其中NHE1和MCTs對(duì)細(xì)胞內(nèi)外pH調(diào)節(jié)至關(guān)重要,特別是在代謝活躍的狀態(tài)下。從這一意義上來(lái)講,MCTs在機(jī)體乳酸循環(huán)、能量代謝以及內(nèi)環(huán)境pH值的調(diào)節(jié)方面發(fā)揮著重要作用[18-19]。外周肌肉組織中,MCT1主要存在于氧化型慢肌細(xì)胞膜和細(xì)胞內(nèi)部,調(diào)控乳酸的攝取與內(nèi)流。MCT4主要在酵解型快肌中高表達(dá),調(diào)控乳酸的排出與外流[20]。而攜帶MCT2的細(xì)胞表現(xiàn)出強(qiáng)烈的乳酸消耗傾向,這與該亞型在諸如神經(jīng)元這類氧化高度活躍的細(xì)胞中高表達(dá)相一致[21]。因此,MCT1、MCT2和MCT4在不同細(xì)胞間對(duì)乳酸轉(zhuǎn)運(yùn)作用各有側(cè)重,才得以協(xié)同完成乳酸穿梭的整個(gè)過(guò)程。Eydoux等[22]研究顯示,短時(shí)間低強(qiáng)度運(yùn)動(dòng)對(duì)外周橫紋肌中MCT1、MCT2和MCT4表達(dá)的影響不甚明顯,而大強(qiáng)度力竭運(yùn)動(dòng)后三者的表達(dá)水平均明顯升高。本研究則針對(duì)運(yùn)動(dòng)后中樞神經(jīng)系統(tǒng)中MCTs表達(dá)的變化趨勢(shì)進(jìn)行了針對(duì)性探究。
乳酸在腦內(nèi)的代謝與外周有所不同。實(shí)驗(yàn)表明,體外灌流乳酸鈉使血液中乳酸濃度顯著升高,但頸內(nèi)靜脈中乳酸濃度變化不大,表明血乳酸變化對(duì)腦乳酸變化影響較小,提示血腦屏障的存在使得外周乳酸很難進(jìn)入腦中。腦內(nèi)胞外乳酸是獨(dú)立于血乳酸而存在的[8,11]。1998年,Pellerin等[11]研究發(fā)現(xiàn)神經(jīng)元與神經(jīng)膠質(zhì)細(xì)胞間的乳酸存在一種特殊的分配狀態(tài),即腦內(nèi)乳酸來(lái)源于星形膠質(zhì)細(xì)胞,而神經(jīng)元?jiǎng)t成為吸收和利用乳酸的部位。其后,有關(guān)腦內(nèi)乳酸作用的研究結(jié)果不盡相同。Chih等[23]綜合了Pellerin等[24]、Wada等[25]和Takata等[26]的研究結(jié)果,提出葡萄糖仍然是神經(jīng)膠質(zhì)細(xì)胞和神經(jīng)元的主要能量底物,但在低血糖等特殊情況下,或由于腦神經(jīng)的多樣性,機(jī)體中某些神經(jīng)細(xì)胞也可能利用乳酸暫時(shí)供能。運(yùn)動(dòng)狀態(tài)下,腦能量供應(yīng)的特殊情況正符合上述研究結(jié)論。目前學(xué)術(shù)界公認(rèn)的是,乳酸由神經(jīng)膠質(zhì)細(xì)胞無(wú)氧糖酵解葡萄糖產(chǎn)生,作為能量底物為神經(jīng)元供能,尤其在劇烈活動(dòng)時(shí)對(duì)維持突觸傳遞具有重要作用。特別是在無(wú)氧或有氧條件下,哺乳動(dòng)物中樞神經(jīng)系統(tǒng)中興奮性神經(jīng)元優(yōu)先利用胞外的乳酸,而不是葡萄糖,作為主要的能量底物[8,27]。以上研究多基于代謝水平的生物化學(xué)檢測(cè)得出結(jié)論,而對(duì)于乳酸代謝轉(zhuǎn)運(yùn)主要媒介MCTs在分子水平的變化情況,尚缺乏系統(tǒng)的研究。因此,本研究重點(diǎn)關(guān)注了MCTs在劇烈運(yùn)動(dòng)的特殊條件下腦內(nèi)表達(dá)的變化情況。研究結(jié)果表明,大鼠運(yùn)動(dòng)能力在疲勞負(fù)荷初期下降較為明顯,此時(shí)MCTs表達(dá)變化不明顯;隨著運(yùn)動(dòng)時(shí)間延長(zhǎng),1周左右后大鼠運(yùn)動(dòng)能力逐漸恢復(fù),相應(yīng)地,鼠腦內(nèi)運(yùn)動(dòng)區(qū)皮層MCT1、MCT2和MCT4的表達(dá)也在相應(yīng)時(shí)間段內(nèi)呈現(xiàn)出逐漸升高的趨勢(shì)。相對(duì)于MCT1和MCT4,皮層運(yùn)動(dòng)神經(jīng)元表達(dá)的MCT2在負(fù)荷運(yùn)動(dòng)后升高較早且趨勢(shì)平緩;這就表明疲勞負(fù)荷運(yùn)動(dòng)后,鼠腦內(nèi)乳酸排出與攝取利用的整個(gè)鏈路均表現(xiàn)出適應(yīng)運(yùn)動(dòng)疲勞的積極變化。上述變化趨勢(shì)正是對(duì)代謝水平乳酸生化檢測(cè)結(jié)論的有力印證。同時(shí),本研究結(jié)果也提示腦內(nèi)MCTs的表達(dá)變化與大鼠對(duì)運(yùn)動(dòng)疲勞的適應(yīng)性存在某種關(guān)聯(lián)性,這種適應(yīng)性在疲勞負(fù)荷運(yùn)動(dòng)1周后維持于相對(duì)平穩(wěn)的狀態(tài)。綜合前述的相關(guān)研究,筆者認(rèn)為腦內(nèi)MCTs表達(dá)變化所代表的腦乳酸代謝調(diào)控可能是機(jī)體對(duì)運(yùn)動(dòng)疲勞(尤其是中樞性運(yùn)動(dòng)疲勞)產(chǎn)生適應(yīng)性的機(jī)制之一。
當(dāng)然,疲勞產(chǎn)生的原因是多方面的,外周肌肉及中樞神經(jīng)系統(tǒng)都參與其中。僅就中樞疲勞而言,興奮性與抑制性氨基酸平衡狀態(tài),腦內(nèi)多巴胺能神經(jīng)元、5-羥色胺等抑制性神經(jīng)遞質(zhì)以及支鏈氨基酸等因素近年來(lái)均有相關(guān)研究[28]。本研究?jī)H對(duì)運(yùn)動(dòng)疲勞狀態(tài)下腦內(nèi)MCTs變化進(jìn)行了初步探索,進(jìn)一步的研究將著重于MCTs分子表達(dá)水平干預(yù)后,運(yùn)動(dòng)疲勞電生理及動(dòng)物行為學(xué)的變化,以期為相關(guān)研究結(jié)果提供進(jìn)一步佐證。
[作者貢獻(xiàn)]
高晨:負(fù)責(zé)實(shí)驗(yàn)總體設(shè)計(jì),數(shù)據(jù)統(tǒng)計(jì)及課題論文撰寫(xiě);
王菀:負(fù)責(zé)標(biāo)準(zhǔn)化大鼠運(yùn)動(dòng)疲勞模型構(gòu)建及運(yùn)動(dòng)能力檢測(cè);
李玉榮:負(fù)責(zé)標(biāo)鼠腦皮層蛋白提取及蛋白質(zhì)印跡檢測(cè);
裴文娟:負(fù)責(zé)鼠腦切片制備及免疫熒光檢測(cè)。
[利益聲明]所有作者均聲明本文不存在利益沖突。
猜你喜歡
中國(guó)設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
 實(shí)驗(yàn)動(dòng)物與比較醫(yī)學(xué)2022年1期
實(shí)驗(yàn)動(dòng)物與比較醫(yī)學(xué)2022年1期
- 實(shí)驗(yàn)動(dòng)物與比較醫(yī)學(xué)的其它文章
- 疼痛抑郁共病動(dòng)物模型及評(píng)價(jià)方法研究進(jìn)展
- 江西省實(shí)驗(yàn)動(dòng)物科技發(fā)展回顧與思考
- 《實(shí)驗(yàn)動(dòng)物與比較醫(yī)學(xué)》2022年專題征稿啟事
- 《實(shí)驗(yàn)動(dòng)物與比較醫(yī)學(xué)》稿約
- 《實(shí)驗(yàn)動(dòng)物與比較醫(yī)學(xué)》有關(guān)實(shí)驗(yàn)動(dòng)物福利倫理內(nèi)容的說(shuō)明
- 《實(shí)驗(yàn)動(dòng)物與比較醫(yī)學(xué)》有關(guān)作者投稿的說(shuō)明