不同B細胞亞群在多發性硬化中的作用①
2022-03-22 12:53:38孫美麗李呼倫哈爾濱醫科大學神經生物學教研室哈爾濱150081
中國免疫學雜志 2022年4期
孫美麗 李呼倫(哈爾濱醫科大學神經生物學教研室,哈爾濱 150081)
多發性硬化(multiple sclerosis,MS)是青少年神經功能障礙最常見的原因之一,其特征是持續的炎癥,脫髓鞘和對中樞神經系統(central nervous system,CNS)不可修復的損傷[1-2]。MS經典臨床分型包括復發緩解型多發性硬化癥(relapsing remitting multiple sclerosis,RRMS)、繼發進展型多發性硬化癥(secondary progressive multiple sclerosis,SPMS)和原發進展型多發性硬化癥(primary progressive multiple sclerosis,PPMS)[3]。其中以RRMS最為常見。MS的實驗動物模型和MS 患者是其主要的研究對象。實驗性自身免疫性腦脊髓炎(experimental autoimmune encepha?lomyelitis,EAE)是RRMS 的公認實驗模型[4],而泰勒氏鼠腦脊髓炎病毒(TMEV)感染是進行性MS 的公認實驗模型[5]。MS 病因尚不清楚,其具有遺傳復雜性,被認為是環境(例如:低水平的維生素D、感染EB病毒、吸煙和過早肥胖)-遺傳相互作用的結果[6]。全基因組關聯研究MS的200多個遺傳風險變異,其中許多編碼了參與免疫系統反應的分子[7],支持了MS 主要是一種免疫介導性疾病的流行概念。已知固有免疫系統和適應性免疫系統及其效應細胞(如B 淋巴細胞、T 淋巴細胞、活化的小膠質細胞和巨噬細胞)會影響MS 的發病機制[2],且T 細胞已被普遍認為是MS 免疫病理機制和CNS 定向自身免疫調節的核心[8-9]。其中CD4+T 細胞、CD8+T 細胞和γδT 細胞在MS的發病機制中發揮相應作用,尤其是CD4+T細胞[10-13]。已知調節性T 細胞(Treg)和CNS 反應性效應T細胞(Teff)之間的平衡失調是MS新發病的基礎,并且糾正這種平衡失調對MS 具有治療效果[14-15]。但在臨床上針對CD4+T 細胞靶向治療MS并沒有達到預期效果[16-22]。因此,可能存在其他因素參與MS 發病機制。值得注意的是,抗CD20 治療MS 顯著有效的結果突出了B 細胞在新的疾病攻擊中的中心作用,包括呈遞抗原、分泌細胞因子和產生抗體[14,23-24]。同時也證實在MS中B 細胞對CD4+T細胞和CD8+T 細胞存在調控作用,但與γδT 細胞的關系還鮮有報道,這值得進一步探索。但隨著在MS中B 細胞耗竭治療(B cell depletion therapy,BCDT)的進行,其副作用問題也漸漸凸顯出來,且在MS 的動物模型EAE 發病早期和高峰期過程中,BCDT 的應用分別出現加劇和抑制疾病癥狀的不同狀態[25-27]。存在這些現象的原因可能是在EAE 免疫病理過程中存在不同的B細亞群所致。目前,在MS中根據起源和功能對B 細胞進行分類具有相應的報道,但在發育過程中對B 細胞的分類還鮮有報道[14,28-29]。B 細胞發育過程中的調節異常會導致自身免疫性疾病的發生[30]。本課題組認為對MS 患者不同發育階段的B 細胞進行研究將為深入探討MS發病機制和臨床治療策略提供新思路。因此,本文將從B 細胞發育過程中不同的B 細胞亞群在MS 發病過程中的作用進行描述。
1 不同B細胞亞群在MS發病過程中的作用
為了更好地了解不同B 細胞亞群在MS 免疫病理過程中的作用,課題組首先總結了B 細胞的發育過程(圖1)。B 細胞最早可分為B1 和B2 兩個亞群,通常所說的B 細胞都是后天發育成熟的,統稱為B2細胞。只有成熟的B細胞才能進入脾臟和淋巴結的淋巴濾泡,從而有效參與免疫應答。為了完成發育,骨髓中未成熟的B 細胞作為過渡性B 細胞(tran?sitional B cells)遷移到脾臟經歷T1 B 細胞和T2 B 細胞兩個過渡階段[31]。在進入外周免疫器官之前,未成熟的B 細胞進行負選擇防止自身反應性的B 細胞進入外周免疫器官[32]。脾臟中過渡性B 細胞通過B細胞受體(B cell receptor,BCR)和其他受體接收的信號分化為邊緣區(marginal zone,MZ)B 細胞或濾泡(follicular,FO)B 細胞[33]。MZ B 細胞和FO B 細胞在外周淋巴組織經歷T 細胞依賴性抗原(TD-Ag)或T 細胞非依賴性抗原(TI-Ag)激活后分化為漿母細胞、漿細胞和記憶B 細胞參與體液免疫反應[34]。而一種先天發育成熟的特殊B 細胞稱為B1細胞,不同于B2 細胞的發育途徑,在無刺激和優先接受TI-Ag激活后分別產生天然免疫球蛋白M(IgM)和后天IgM[35]。B1細胞是具有自我更新能力的CD5+的B細胞,主要來自腹膜腔和胸膜腔,是天然IgM 分泌的主要來源,并參與固有免疫反應[35-36]。值得注意的是,有證據表明幾乎所有的B細胞亞型都可以通過涉及炎癥信號和BCR 識別的機制分化為調節性B 細胞(Breg)[37]。
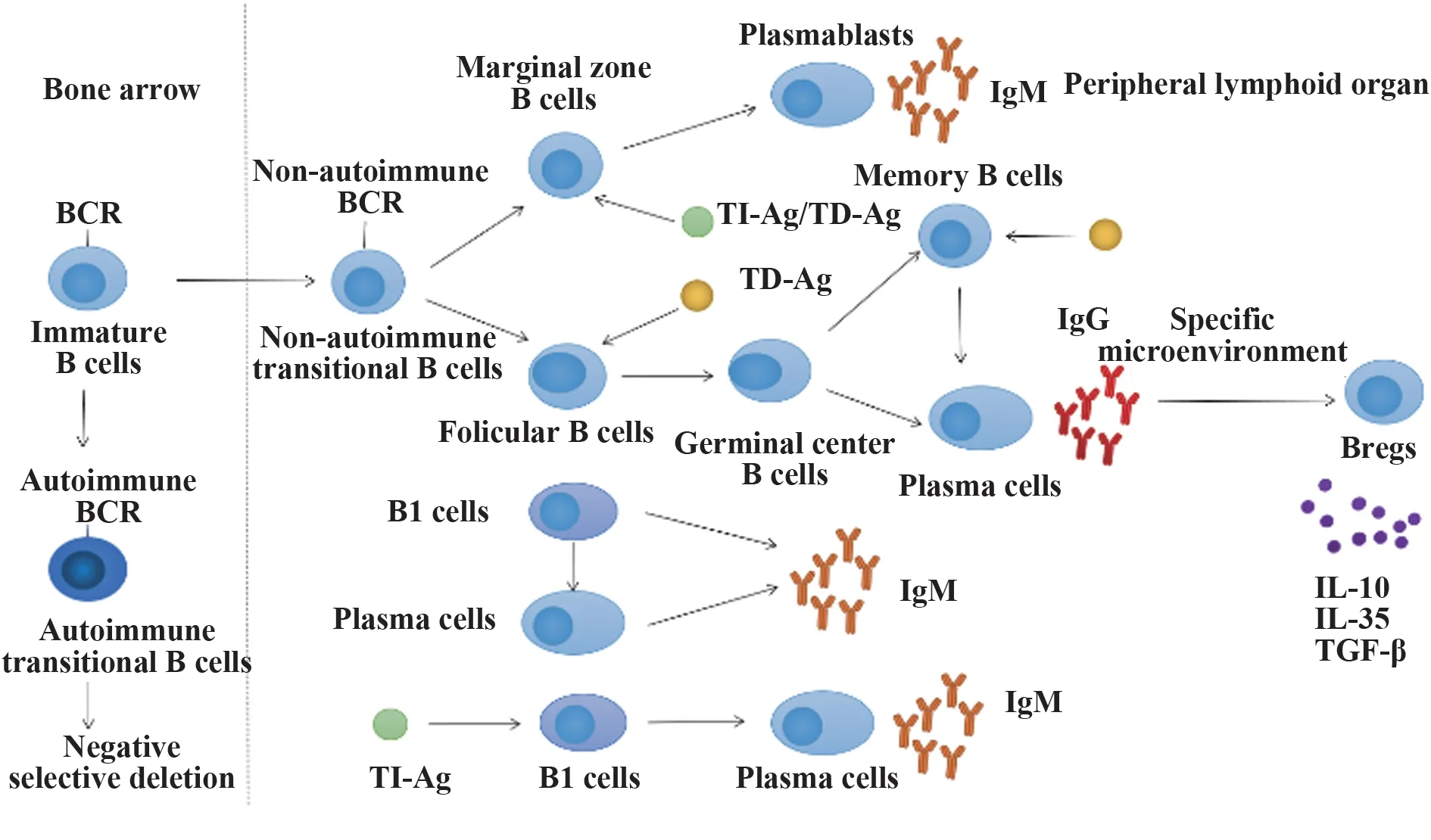
圖1 B細胞的發育過程Fig.1 Process of B cell development
1.1 過渡性B細胞在MS免疫病理過程中的作用過渡性B細胞過渡性發育階段是人體對自身反應性B 細胞存活與凋亡的關鍵選擇點。共刺激分子CD40、BAFF-R、TACI 和TLRs 可能在表達自身免疫BCR 的過渡性B 細胞發育為成熟的幼稚B 細胞的過程中發揮積極作用[38]。KLHL6 基因缺陷有利于過渡性B 細胞存活和分化為成熟的幼稚B 細胞,為自身免疫性疾病的治療提供了新的理論基礎[39]。在自身免疫性疾病系統性紅斑狼瘡(systemic lupus erythematosus,SLE)和干燥綜合征(Sj?gren syndrome,SS)患者血液中,B 細胞穩態失調(主要包括過渡性B 細胞增多)導致各種器官功能障礙[40]。在放射學孤立綜合征(RIS)和臨床孤立綜合征(CIS)中,過渡性B 細胞亞群比例改變[41]。以上說明過渡性B 細胞在自身免疫性疾病過程中發揮重要作用。因此,自身反應性過渡性B 細胞的清除可能為MS 或自身免疫性疾病治療的對策。
此外,CIS和RRMS患者外周血中的過渡性B 細胞減少,但CD80 陽性活化狀態的過渡性B 細胞增加,同時在腦脊液(CSF)中發現存在過渡性B 細胞[42]。在CIS 和MS 患者體內,過渡性B 細胞中整合素α4 和β1 鏈表達均升高(僅MS 患者α4 鏈的升高沒有統計學意義)[42-43],說明過渡性B 細胞高表達CD80 和黏附分子的特殊狀態可能是其穿越血腦屏障的必備條件,是MS 疾病早期階段的一個特征。基于此開發的抗VLA-4 抗體Natalizumab 阻斷整合素α4β1 已被保留用于高度活躍的RRMS 患者[44]。因此尋找抑制整合素α4、β1或CD80的方法可能是抑制過渡性B細胞進入早期MS患者CNS和分化為自身反應性成熟的幼稚B細胞治療MS的另一重要方向。
相反,在EAE 中過渡性B 細胞能作為Breg 產生IL-10 發揮其調節作用[45-46]。未來在充分理解自身免疫性過渡性B細胞存活與凋亡的具體信號通路和過渡性B細胞發育調節機制及其遷移和逃逸途徑的基礎上,研發過渡性B細胞清除和阻礙遷移的藥物可能是治療MS或其他自身免疫性疾病的重要策略。
1.2 濾泡B細胞在MS免疫病理過程中的作用 淋巴濾泡生發中心是活化的FO B 細胞發生體細胞超突變、類轉換重組和親和力成熟選擇分化為記憶性B細胞或分泌高親和力抗體的漿細胞的微環境[47-48]。有報道稱在CNS 自身免疫性疾病慢性炎癥條件下,免疫特權較低的腦膜腔室是異位淋巴濾泡優先發育的部位,致病性B 細胞反應可在此維持[49]。MS患者的腦膜中包含帶有生發中心的異位淋巴樣結構,包括增殖B 細胞、T 細胞和漿細胞以及產生CXCL13的濾泡樹突狀細胞網絡,其形成可能是維持體液自身免疫和疾病惡化的關鍵步驟[50-51]。SPMS 患者軟腦膜的蛛網膜下腔中,濾泡樣結構與皮質脫髓鞘增加、神經元丟失、腦膜浸潤和CNS 炎癥以及發展為嚴重殘疾和死亡有關[52]。綜上說明,FO B 細胞在活化后既可分化為產生抗體的漿細胞參與適應性體液免疫應答,又能作為成熟B 細胞發揮抗體非依賴性功能。因此,深入了解FO B細胞向漿細胞分化的機制和作為成熟B 細胞發揮的功能可能有助于MS或自身免疫性疾病發病機制的解析和治療對策的研究。轉錄因子B 淋巴細胞誘導成熟蛋白1(Blimp1)在協調漿細胞分化中發揮多種功能[53]。激活的B 細胞的增殖需要促進Blimp1 表達,但需要終止增殖以避免過度的免疫反應[54]。B 細胞特異性激活蛋白1(AP-1)轉錄因子Fra1缺失的小鼠體內漿細胞分化和抗體反應增加,證實Fra1 通過抑制Blimp1的表達來抑制漿細胞分化[54]。目前還沒有相關報道闡明Fral對MS或其他自身免疫性疾病的影響,值得進一步探索。
1.3 漿母細胞和漿細胞在MS 免疫病理過程中的作用 在RRMS 中,外周血濾泡輔助性T 細胞(TfH)與濾泡調節性T 細胞(Tfr)比值(TfH/Tfr)增加,且與血液和CSF 中的異常IgG 產生相關,表明源自外周生發中心反應失調的抗體產生細胞(漿母細胞/漿細胞)在CNS 上定殖[55]。除了產生抗體,在小鼠中漿母細胞/漿細胞是產生細胞因子IL-10、IL-35、TNF-α、IL-17 和GM-CSF 的活化B 細胞的主要類型,表明存在不同的漿母細胞/漿細胞亞群,這些亞群可通過細胞因子的表達模式來區分[56]。此外,在EAE 中漿細胞通過IL-10、IL-35細胞因子的產生調節免疫[56]。
1.4 記憶B細胞在MS免疫病理過程中的作用 在MS 中,自身反應性B 細胞逃避了外周選擇,并可能在次級淋巴器官淋巴濾泡內分化為記憶群,然后進入CNS[57-58]。與外周血相比,MS 患者CNS 記憶B 細胞顯著升高[59]。MS 患者CSF 中存在的寡克隆區帶(OCB)提示這些記憶B 細胞在局部發生反應從而再活化,進一步發展為產生免疫球蛋白(Ig)的記憶細胞或漿細胞[50]。Atacicept(阿塞西普)可增強記憶B 細胞反應,并促使MS 復發,說明記憶B 細胞在復發性MS 中至關重要[60],同時臨床上Atacicept 的應用帶來疾病的復發值得進一步關注。在MS 中記憶B細胞接受BCR和CD40刺激后分泌LT和TNF-α[61]。MS 患者的記憶B 細胞能有效地向T 細胞呈遞神經抗原,激活T 細胞[62-63]。有報道指出,在MS 患者不同的CNS 隔室中選擇性富集CXCR3+IgG1+記憶B 細胞,并且IFN-γ 和TLR9 信號能觸發該細胞群分化[64],因此,確定MS 患者CNS 這些記憶細胞群特點,并尋找能夠調節它們形成,進行招募和局部反應性的外界因素并加以抑制可能有效緩解MS 的病理變化。B 細胞淋巴瘤-6蛋白(Bcl-6)對記憶B 細胞的生成和維持其更新的能力是必需的[65-66]。此外,TfH分化受Bcl-6調控,且TfH調節異常與自身免疫性疾病SLE、MS 和類風濕關節炎(rheumatoid arthritis,RA)有關[67-68]。已有報道指出Bcl-6可通過調控Tfr和巨噬細胞活性來影響自身免疫的發生與發展[69-70]。目前尚無Bcl-6通過記憶B細胞對MS或其他自身免疫性疾病影響的相關報道,值得進一步探索。
1.5 B1 細胞與MZ B 細胞在MS 免疫病理過程中可能存在的作用 盡管B1細胞與MZ B細胞的發育關系尚不明確,二者可能來源于不同的譜系,但二者在某些功能上似乎有相同的作用[71-73]。B1 細胞與MZ B 細胞能夠通過TLR 快速對脂多糖刺激進行反應,并分化為短壽命的漿母細胞,產生大量的IgM 及同型轉換抗體,這些與生俱來的淋巴細胞的特殊功能彌補了早期固有免疫反應和較慢的適應性抗體反應之間的差距[74]。
在B 細胞非依賴性EAE 中,IFN-β 改善了神經炎癥,而在依賴B 細胞的EAE 中,IFN-β 對疾病沒有影響,且在兩種EAE 模型中,IFN-β 均增加了MZ B細胞[75],這些發現表明增加的MZ B 細胞可能影響EAE 中IFN-β 的功效。MZB1 是B 細胞特異性ER 分子伴侶復合物的一部分,在MZ B 細胞和B1 細胞中高表達,能調節成熟IgM 的適當組裝和分泌[76-77]。在自身免疫性疾病SLE 患者淋巴結和RA 患者滑膜組織標本中的MZB1+MZ B 細胞增加,因此MZB1 可能是SLE 和RA 潛在的治療靶標[78]。在MS 中MZB1是否存在異常和是否能夠作為潛在的治療靶標值得進一步探索。
MS 患者的腦脊液中表達CD5 的B 細胞增加并且具有致病作用,RRMS 患者的外周血中B1 細胞亞群減少[79-80]。這些現象表明MS 患者外周血B1 細胞可能進入CNS 后影響疾病病理變化。先天性反應激活劑IRA-B細胞(來源于B1a細胞)產生的GM-CSF能加速病原體的清除,預防敗血性休克[81]。在動脈粥樣硬化中IRA-B 細胞以GM-CSF 依賴的方式激活Th1 適應性免疫進而加重疾病進展[82]。GM-CSF 是罕見的MS 發病機制中必需的細胞因子之一[83],目前尚無IRA-B 細胞與MS 相關的報道,值得進一步思考。
此外MZ B細胞和B1細胞是脾臟中B細胞IL-10的主要來源[84]。在狼瘡小鼠中,TLR-9 激活的MZ B細胞通過分泌IL-10調節T細胞介導的炎癥反應[85]。在膠原蛋白誘導性關節炎(collagen-induced arthritis,CIA)(常用的RA 模型)緩解階段,表達T-bet 的MZ B細胞向CXCR3配體遷移并在炎癥刺激后(anti-IgM、CpG 和IFN-γ)分泌IL-10 調節CIA 的緩解[86]。目前激活的MZ B 是否也存在同樣的緩解MS 的作用鮮有報道,值得進一步探索。
1.6 Breg 在MS 免疫病理過程中的作用 在慢性炎癥疾病(如MS)過程中,免疫系統被持續激活,其特征是Breg 在循環和炎癥部位的數量與功能不足[87]。在MS 和其他自身免疫性疾病中,Breg 在抑制疾病活動和發生與發展中發揮重要作用[88]。在MS 中Breg 通過抑制樹突狀細胞(dendritic cells,DCs)產生促炎細胞因子間接地抑制致病性T 細胞分化[89];Breg 通過同源相互作用誘導Treg 生成而抑制自身免疫性疾病的發生與發展[90];此外,Breg 通過分泌細胞因子IL-10、IL-35、TGF-β1 抑制炎癥反應[91]。有證據表明,幾乎所有的B 細胞亞群都可以通過涉及炎癥信號和BCR 識別的機制分化為Breg[37]。MS 的未來研究和治療策略中,識別誘導B細胞成為Breg 必需的刺激物是一個重要的考慮因素,如TLR 和/或CD40 激活是已知誘導其分化的最有特征的信號[92]。當然不能排除將來可能會發現定義Breg功能的轉錄調控因子。
綜上所述,不同B細胞亞群在MS中的相對貢獻存在差異,每種B 細胞亞群同時具有致病作用和保護作用,兩種作用失衡可能是導致疾病發展的主要原因。因此,消除或抑制致病性B 細胞亞群或尋找抑制該亞群形成的因素可能對MS具有治療作用。
2 結語與展望
在MS的發病過程中B細胞發揮重要作用,然而BCDT 存在一定的副作用,這使研究者的關注點集中在B 細胞亞群上。然而不同B 細胞亞群的相對貢獻存在差異,是否存在其他對MS有影響的B細胞亞群還未知。目前在MS 中靶向特異B 細胞亞群的治療方法很少,在臨床上,針對不同B細胞亞群開發和應用特異性的靶向致病性B細胞亞群的藥物來治療MS是未來需要解決的重要問題。
