急性熱應激對睪丸免疫微環境相關蛋白的影響①
2022-03-23 01:17:56胡素芹劉殿龍李春蕊許亞輝崔力丹北京中醫藥大學中醫學院北京102488
中國免疫學雜志 2022年5期
胡素芹 胡 珂 劉殿龍 李春蕊 許亞輝 崔力丹 郭 健(北京中醫藥大學中醫學院,北京 102488)
近年來,男性生殖健康一直被廣泛關注。研究證實,環境因素顯著影響精子數量和質量,嚴重阻礙男性生殖功能[1-2]。陰囊溫度時常會受到自身坐勢、穿著、生活方式、職業等外部因素的影響[3]。在眾多環境影響因素中,盡管睪丸熱應激被證明是男性生育能力降低的主要危害之一,但卻最易被忽略[4-5]。此外,隨著全球氣候變暖,有害物質的不利影響將逐漸惡化[6]。
睪丸是典型的免疫豁免組織,具有特殊的免疫調節機制,該機制在精子發生過程中發揮重要作用。免疫豁免和免疫防御兩方面協調存在,共同維持了正常的睪丸免疫微環境平衡[7]。當這種免疫微環境穩態失衡時,睪丸就會發生炎癥并損害精子發生過程,影響男性生殖功能。血睪屏障由睪丸支持細胞通過細胞間緊密連接、縫隙連接和錨定連接構成,是執行免疫豁免功能的天然物理屏障,其嚴格限制分子和細胞通過,阻止病原微生物或抗原的侵襲,同時可阻止循環中的抗體進入生殖道管腔,避免因免疫應答或炎癥反應造成局部損傷,為精子發生提供了適宜的免疫微環境[8-9]。血睪屏障的破壞可導致免疫豁免功能的破壞,進而誘發機體自身免疫應答,導致不育[10]。而高溫對睪丸內免疫微環境的影響尚不清楚。本實驗旨在觀察急性熱應激后不同時間睪丸內血睪屏障相關分子及經典炎癥通路NF-κB 信號通路的變化,進而探討熱應激對睪丸免疫微環境的影響。
1 材料與方法
1.1 材料
1.1.1 實驗動物 SPF 級成年雄性SD 大鼠49 只,8 周齡,體質量310~340 g,購自北京斯貝福實驗動物有限公司,許可證號:SCXK(京)2019-0010。大鼠飼養環境溫度控制在22~25℃,相對濕度40%~70%,晝夜節律每12 h 交替,自由飲水和進食,所有大鼠均適應性喂養1周后進入本實驗。
1.1.2 主要試劑 生理鹽水(石家莊四藥有限公司);甲醇(北京化工廠);中性裂解液(北京普利萊基因技術有限公司);苯甲基磺酰氟(phenylmethanesulfonyl fluoride,PMSF)、蛋白磷酸酶抑制劑(北京索萊寶科技有限公司);BCA 蛋白定量試劑盒、30%丙烯酰胺、10×TBST、山羊抗兔IgG(H+L)Biotin-conjugated、過氧化物酶標記鏈酶親和素(康為世紀生物科技有限公司);TEMED(BioRule 公司);Page Ruler Prestained Protein Ladder(Thermo Scientific 公司);PVDF 膜(美國millipore 公司);兔抗大鼠Vimentin、NF-κB p65、phospho-NF-κB p65、IκBα、phospho-IκBα單克隆抗體(美國Cell Signaling Technology 公司);兔抗大鼠occludin、β-actin 多克隆抗體(美國Proteintech 公司);兔抗大鼠ZO-1 單克隆抗體(美國賽默飛invitrogin 公司);IL-1β ELISA 試劑盒(美國Raybiotech公司)。
1.1.3 主要儀器 SSW-420-2S 恒溫水浴鍋(上海博訊實業有限公司醫療設備廠);VCX150 超聲破碎儀(美國SONICS 公司);Mini Trans-Blot cell 電泳槽、ChemiDoc MP 凝膠成像系統、iMark 酶標儀(美國Bio-Rad公司);DYY-6C電泳儀(北京市六一儀器廠)。
1.2 方法
1.2.1 分組與造模 SD 大鼠按照體質量隨機分為7組,即1個對照組和6個熱應激組(0.5 h組、2 h組、4 h 組、6 h 組、24 h 組和48 h 組),7 只/組。適應性喂養1 周后,除對照組以外,其余6 組大鼠用2%戊巴比妥鈉(45 mg/kg)麻醉后,43℃恒溫水浴20 min,并分別于水浴結束后0.5 h、2 h、4 h、6 h、24 h 和48 h的6個時間點摘取睪丸,-80℃冰箱凍存。
1.2.2 Western blot 檢測大鼠睪丸內蛋白表達變化 睪丸組織加入適量PMSF、磷酸酶抑制劑和中性裂解液并研磨破碎,以12 000 r/min 低溫離心5 min,收集上清液。BCA 蛋白定量試劑盒檢測蛋白濃度后,100℃煮沸5 min 進行蛋白變性。10%SDS-聚丙烯酰胺凝膠電泳,電轉至PVDF 膜(孔徑0.45 μm)。PVDF 膜用5%BCA 封閉液室溫封閉1.5 h,而后用1%BCA 稀釋的一抗孵育,4℃過夜。第2 天PVDF 膜室溫搖床0.5 h,TBST 清洗4 次,5 min/次。PVDF 膜放入山羊抗兔IgG(H+L)Biotin conjugated 溶液(TBST 稀釋,1∶1 000),37℃孵育1.5 h。TBST 洗膜5 min×4 次,過氧化物酶標記鏈酶親和素溶液(TBST 稀釋,1∶1 000)37℃孵育0.5 h。TBST 洗膜5 min×4 次后,PVDF 膜上加ECL 發光液反應1~3 min,然后置于凝膠成像系統中檢測,所得圖片使用Image J軟件分析,檢測各條帶的灰度值。
1.2.3 ELISA法檢測大鼠睪丸內IL-1β表達 采用ELISA法檢測睪丸組織內IL-1β的表達,嚴格按照試劑盒說明書進行操作。微孔板用酶標儀檢測每孔OD 值,450 nm 為檢測波長,630 nm 為參考波長。所得OD 值用ELISA Cal 軟件繪制標準曲線,計算各組IL-1β濃度。
1.3 統計學處理 采用SPSS20.0軟件進行數據分析,計量資料以表示。方差齊,多組間比較采用單因素方差分析(One-Way-ANOVA),兩兩比較采用LSD檢驗,以P<0.05為差異有統計學意義。
2 結果
2.1 急性熱應激下調睪丸內波形蛋白vimentin 及血睪屏障緊密連接相關蛋白occludin和ZO-1表達與對照組相比,6個熱應激組大鼠睪丸內vimentin和occludin 的表達均顯著降低(P<0.01);除0.5 h 組外,其余熱應激組ZO-1表達相較于對照組均顯著降低(P<0.01),如圖1。
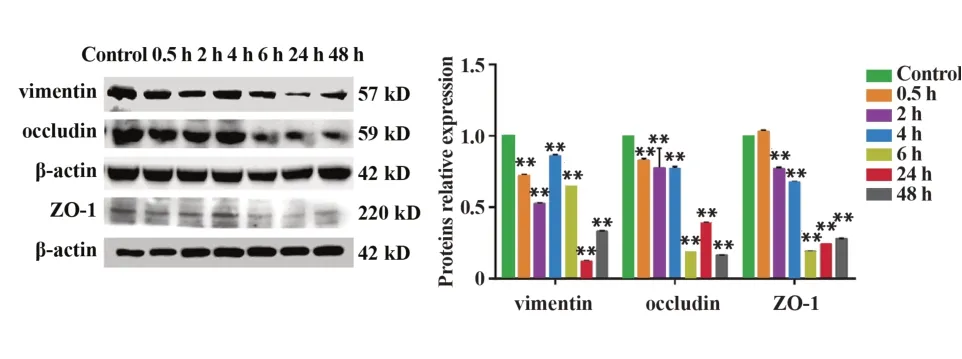
圖1 各組大鼠睪丸內vimentin、occludin和ZO-1表達Fig.1 Expressions of vimentin,occludin and ZO-1 in testis of rats in each group
2.2 急性熱應激上調睪丸內NF-κB 信號通路關鍵分子的表達 與對照組相比,0.5 h 組、2 h 組和4 h組p-NF-κB p65 及p-IκBα 表達變化差異無統計學意義(P>0.05),6 h 組、24 h 組和48 h 組p-NF-κB p65及p-IκBα的表達均顯著增加(P<0.01),如圖2。
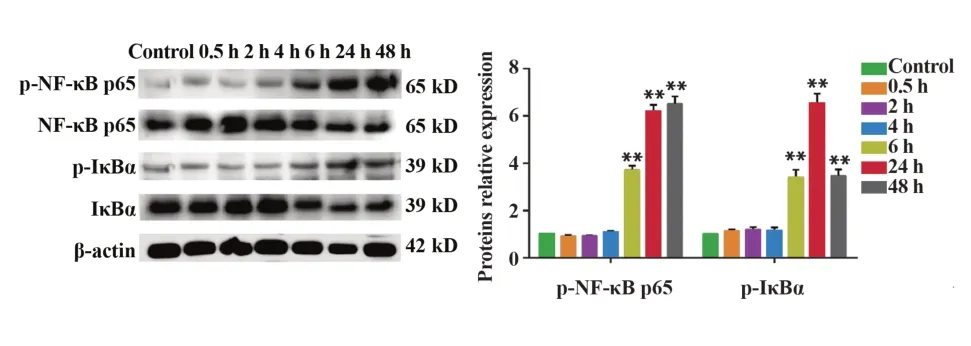
圖2 各組大鼠睪丸內p-NF-κB p65和p-IκBα表達Fig.2 Expressions of p-NF-κB p65 and p-IκBα in testis of rats in each group
2.3 急性熱應激后睪丸內IL-1β 的表達 與對照組相比,24 h 和48 h 熱應激組大鼠睪丸內IL-1β 表達的差異無統計學意義(P>0.05),見表1、圖3。
表1 各組大鼠睪丸組織內IL-1β的表達(,n=6)Tab.1 Expression of IL-1β in testis of rats in each group(,n=6)

表1 各組大鼠睪丸組織內IL-1β的表達(,n=6)Tab.1 Expression of IL-1β in testis of rats in each group(,n=6)
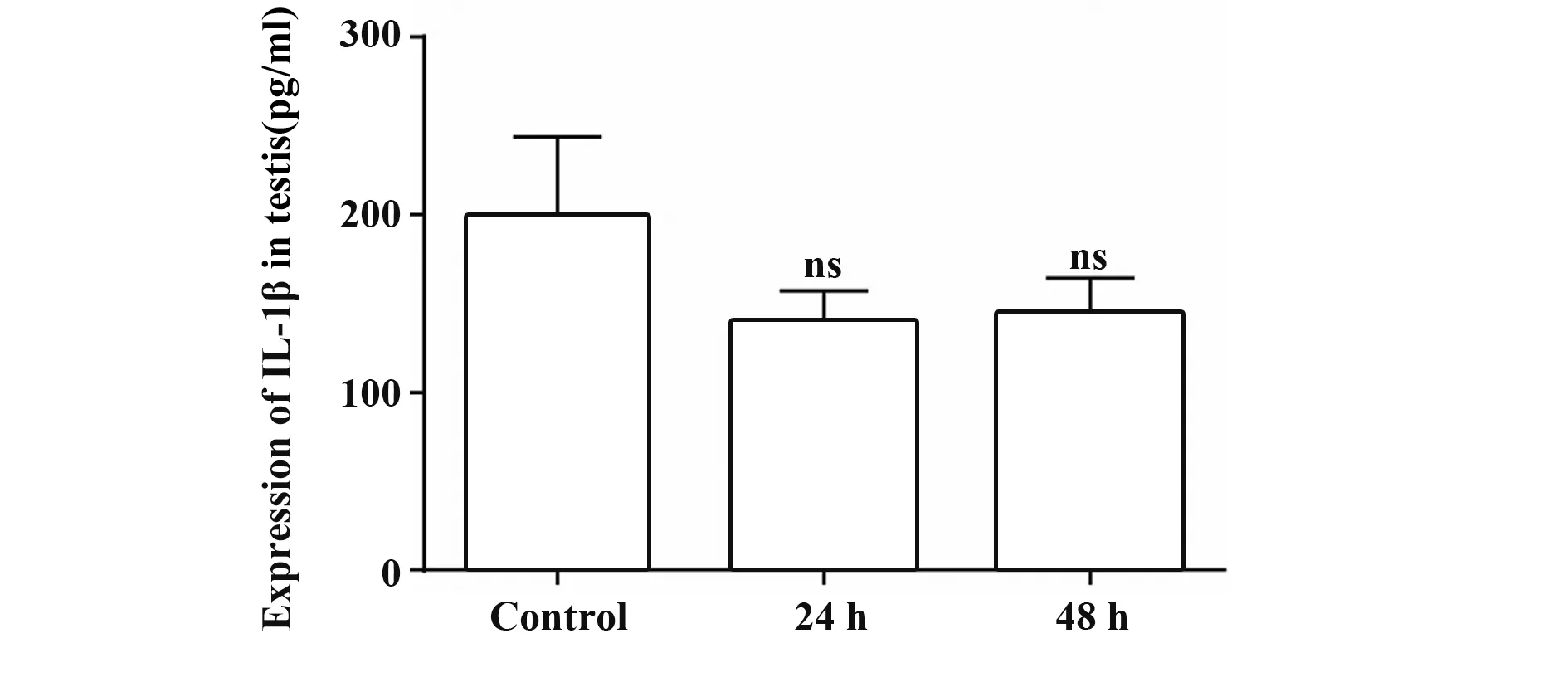
圖3 大鼠睪丸內IL-1β的表達變化Fig.3 Expression change of IL-1β in testis of rats
3 討論
精子發生是一個復雜的過程,會產生抗原性蛋白卻并未誘發免疫炎癥反應,這有賴于睪丸特殊的免疫微環境和免疫調節功能[11]。血睪屏障作為天然物理屏障,在睪丸的免疫豁免功能中發揮重要作用[12-13]。本課題組前期研究發現五子衍宗丸復方及枸杞多糖,菟絲子黃酮可有效干預熱應激后導致的血睪屏障破壞及生精細胞的凋亡[14-16]。此外,有研究發現熱應激可誘發機體各種炎癥反應[17-19]。因此推測急性熱應激會引起睪丸內免疫微環境變化,該變化可能誘發了炎癥反應。
血睪屏障由多種支持細胞膜蛋白(如ZO-1、occludin 和claudins)組成,是睪丸免疫耐受的結構基礎[20]。其可限制大分子物質,特別是蛋白質通過細胞間隙,確保曲細精管與間質內的環境顯著不同,并為生精細胞的正常發育創造了獨特的環境,保證生精細胞免受免疫系統的影響[21]。一旦緊密連接結構相關蛋白表達缺失,破壞了睪丸的免疫豁免屏障,則有可能導致自身免疫炎癥反應的發生[22]。而vimentin 出現在支持細胞的基底和核周區域,并向頂端細胞質輻射,負責將生精細胞錨定到生精上皮[23]。與血睪屏障共同維持睪丸的免疫豁免環境。vimentin 塌陷會破壞生精上皮結構的完整性,造成生精細胞脫落、凋亡[24]。本研究結果表明急性熱應激后0.5 h 即出現睪丸內生精上皮結構和血睪屏障通透性被破壞,并且熱應激后48 h 內破壞程度隨著時間的增長有增強的趨勢。提示急性熱應激生精障礙早期干預治療對減輕睪丸組織損傷十分關鍵。
NF-κB p65 是存在于胞漿中的一種核轉錄因子,參與并調控炎癥反應和免疫應答。細胞在靜息狀態下,NF-κB p65 與IκBα 相結合,以聚合物的形式存在于胞漿內,無轉錄活性[25]。當受到外界刺激時,IκBα 磷酸化,隨即從聚合體中解離,再經泛素化修飾后,IκBα 發生降解[26-27]。而受到IκBα 抑制的NF-κB p65 得以解離并迅速進入細胞核內,發生磷酸化并誘導產生炎癥介質(如IL-1、TNF-α、IL-6等)[28]。本實驗結果證明急性熱應激6 h 以后NF-κB 信號通路才被激活,其中24 h 和48 h 差異最顯著。因此實驗檢測了這兩個時間點熱應激大鼠睪丸內IL-1β 的表達變化。IL-1β 是由NF-κB 信號通路調控的下游重要炎癥相關因子之一,能夠阻遏睪酮合成蛋白酶基因的表達和轉錄,阻礙精子發生,降低精子活力,減少精子數量,從而導致不育[29-30]。有趣的是,本實驗結果表明,雖然NF-κB 信號通路被激活,但睪丸內炎癥因子卻沒有增多,并未發生免疫炎癥反應。這可能是因為睪丸內免疫豁免功能由多種具有免疫抑制功能的細胞和復雜的細胞因子網絡共同構成和調節[7,31]。急性熱應激的刺激程度未超出睪丸免疫抑制的調節范圍。
綜上所述,本研究結果表明急性熱應激在早期即可破壞睪丸內血睪屏障及生精上皮結構,破壞睪丸免疫豁免的屏障,同時在中期即激活NF-κB 信號通路。但該熱激刺激并未導致下游炎癥因子增多,未誘發炎癥反應。有研究表明,持續性的熱刺激更能誘發睪丸產生明顯的炎癥反應和相應的炎癥損傷[32-33]。熱應激誘發睪丸局部炎癥反應需要時間界點和溫度閾值。因此,繼續加大熱刺激程度觀察睪丸內免疫微環境的改變及其對睪丸組織細胞的影響,將是本課題組今后的研究重點。
