山東半島不同種群鰻草種子形態(tài)學特征及萌發(fā)特性研究?
2022-03-26 02:10:46謝坤秀牛淑娜遲會凱張沛東
中國海洋大學學報(自然科學版) 2022年4期
謝坤秀,牛淑娜,遲會凱,張沛東
(海水養(yǎng)殖教育部重點實驗室(中國海洋大學),山東 青島 266003)
海草床作為三大典型近海海洋生態(tài)系統(tǒng)之一,具有極高的生態(tài)服務價值,不僅能為眾多的海洋動物提供食物來源與產(chǎn)卵、育幼、棲息、庇佑場所,還能起到減緩水流、凈化水體、穩(wěn)定沉積物、固碳等作用[1-7]。然而,海草床屬脆弱性生態(tài)系統(tǒng),受人類活動及自然因素影響,當前世界范圍內(nèi)的海草床不斷退化,消失速率超過每年7%[8]。由于海草種子具有體積小、便于運輸、能保持遺傳多樣性等優(yōu)勢,利用種子種植法進行海草床規(guī)模化修復逐漸成為近年來研究的熱點。
種子種植法的成功主要取決于種子萌發(fā)與實生苗建成的效果,而種子的萌發(fā)受到不同種群的顯著影響。van Lent等[9]發(fā)現(xiàn),荷蘭西南部和法國西部不同種群的鰻草(Zosteramarina)種子萌發(fā)率相差約15%。Koch等[10]發(fā)現(xiàn),美國南部佛羅里達海域的川蔓草(Ruppiamaritima)種子萌發(fā)率顯著低于東南部的北卡羅來納州種群。影響種子萌發(fā)及實生苗活力的因素通常分為外部因素和內(nèi)部因素,其中外部因素主要是種子萌發(fā)的環(huán)境條件,內(nèi)部因素則主要包括種子規(guī)格和營養(yǎng)貯藏物質(zhì)的含量[11-13]。目前,對于陸生植物種群間種子形態(tài)及營養(yǎng)物質(zhì)含量對種子萌發(fā)及幼苗生長的研究已有許多報道[14-16],但關于海草的相關研究還較少。
鰻草是中國溫帶海域海草的優(yōu)勢種類,主要分布于山東、河北、遼寧沿海[17]。調(diào)查顯示,山東半島日照海域和乳山海域的鰻草海草床已經(jīng)消失,而萊州灣芙蓉島附近海域的鰻草海草床面積則從1 300 hm2退化至不足5 hm2[17]。當前,山東半島主要以威海近海和青島近海分布有面積較大的鰻草海草床。然而不同種群的鰻草種子規(guī)格、萌發(fā)特性和實生苗生長狀況等存在的差異尚不明確。因此,探明不同種群鰻草種子的形態(tài)學特征和萌發(fā)特性,是實現(xiàn)優(yōu)質(zhì)種源篩選以及成功開展海草種子種植的關鍵。
本研究以鰻草為對象,分別采集山東威海天鵝湖海域、俚島灣海域和青島匯泉灣海域的鰻草種子,比較分析不同種群間鰻草種子的形態(tài)學及內(nèi)部營養(yǎng)物質(zhì)含量的異同;通過室內(nèi)萌發(fā)實驗,探討不同種群間鰻草種子萌發(fā)率、實生苗存活率和生長的差異,從而查明不同種群鰻草種子的萌發(fā)策略,為優(yōu)化鰻草種子種植技術提供理論參考。
1 材料與方法
1.1 種子采集及保存
2019年7月,分別于山東省威海市天鵝湖海域、俚島灣海域和山東省青島市匯泉灣海域采集鰻草種子(見圖1)。首先選取鰻草分布較多的區(qū)域,通過人工方式收集生殖枝,收集過程中未破壞鰻草植株的營養(yǎng)枝及其地下組織,且未在同一區(qū)域進行大量采集。摘取具有種子的佛焰苞放入33 cm× 33 cm(長×寬)、網(wǎng)目大小為 1.1 mm的尼龍網(wǎng)兜中,將其置于自然海域并固定。待尼龍網(wǎng)兜中的佛焰苞充分降解后,將其取出倒入塑料桶中反復攪拌,去除漂浮的有機碎屑及小型貝類、蟲類外殼等雜質(zhì),篩選出成熟的鰻草種子。篩選后的鰻草種子收集至廣口瓶帶回實驗室,置于溫度4 ℃、鹽度30的海水中儲存,期間每2天換水1次,每天定期攪拌以減少水中的真菌數(shù)量。
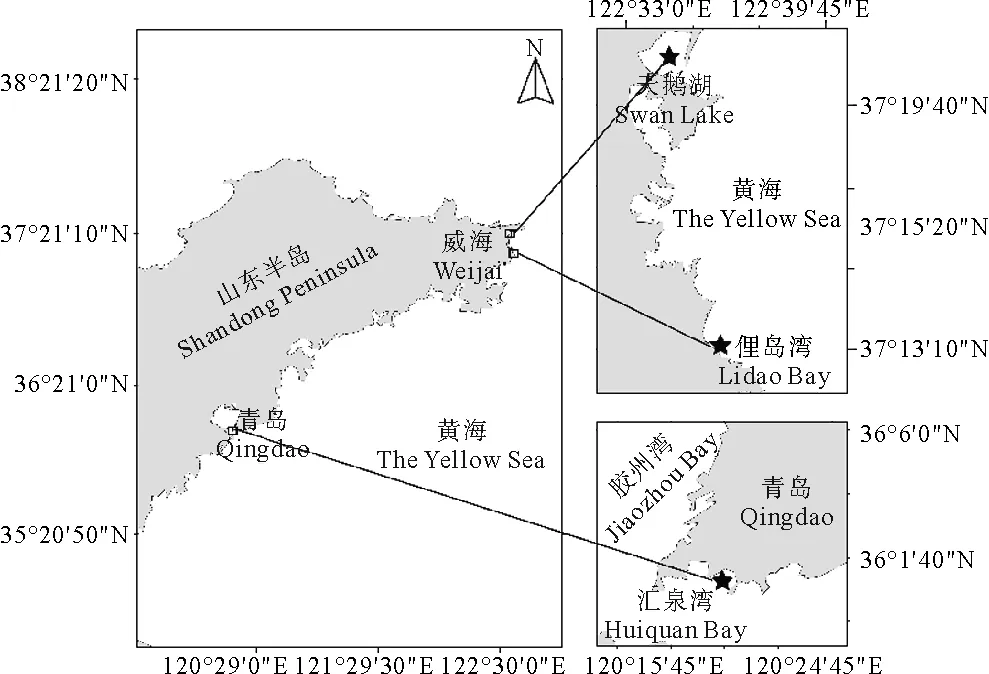
(星號代表采集地點。Asterisk indicates collection site.)
1.2 實驗過程與樣品分析
1.2.1 不同種群鰻草種子的形態(tài)學比較 從不同種群的鰻草種子中隨機選取成熟飽滿種子各100粒,吸干表面水分后分別測定種子的質(zhì)量、長徑、短徑、縱肋數(shù)和最大橫截面積等形態(tài)學指標。根據(jù)種子的長徑和短徑,計算種形系數(shù)(長徑/短徑)[18]。
1.2.2 不同種群鰻草種子的內(nèi)部貯存物質(zhì)測定 從不同種群的鰻草種子中隨機選取成熟飽滿種子各1 000粒,隨機分為10份,分別對每份種子測定水分、總蛋白、總脂肪和總糖等內(nèi)部營養(yǎng)貯存物質(zhì)的含量,其中水分使用烘干法[19],總蛋白含量使用凱氏定氮法[20],總脂肪含量使用酸水解法[21],總糖含量使用3,5-二硝基水楊酸比色法[22]。
1.2.3 不同種群鰻草種子萌發(fā)的比較 于2019年11月,從不同種群的鰻草種子中隨機選取成熟飽滿的種子各300粒,先使用75%乙醇溶液浸泡0.5 min,無菌海水清洗3遍,再使用0.1%氯化汞溶液浸泡15 min,無菌海水清洗3遍。消毒后將各種群的種子平均分為3份,每份100粒,分別放入100 mL燒杯,燒杯中加入80 mL等量的鹽度10的海水,置于光照培養(yǎng)箱(GXZ-0288,寧波江南儀器廠)中進行種子的快速萌發(fā)。萌發(fā)過程無光照,溫度15 ℃。萌發(fā)實驗共進行30 d,期間每天觀察種子萌發(fā)情況并統(tǒng)計萌發(fā)數(shù),將已萌發(fā)的種子挑出進行下一步的實生苗培養(yǎng)實驗,每2天換水1次。實驗結束后,計算各種群種子的累積萌發(fā)率、發(fā)芽指數(shù)(GI)和平均萌發(fā)歷期(MTG)。
萌發(fā)率=萌發(fā)的種子數(shù)/種子總數(shù)×100%;
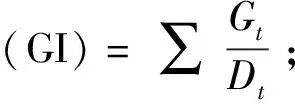
式中:Dt為發(fā)芽日數(shù);Gt為與Dt對應天數(shù)的萌發(fā)粒數(shù);n為相應天數(shù)萌發(fā)的種子數(shù);d為計算種子萌發(fā)數(shù)量時的萌發(fā)天數(shù);N為萌發(fā)種子的總數(shù)。
1.2.4 不同種群鰻草實生苗存活、生長及光合生理的比較 經(jīng)種子快速萌發(fā),天鵝湖、俚島灣和匯泉灣種群分別獲得124、113和81株實生苗。將不同種群獲得的實生苗以鹽度每天升高5的速率進行鹽度馴化,至鹽度30停止。各種群馴化后的實生苗以20株為1組種植于底部鋪設5 cm厚細沙的100 mL燒杯中,株數(shù)取整后各種群分別使用4~6個燒杯(4~6個重復),并用防水紙標識組別,然后將燒杯放入40 cm × 40 cm × 20 cm(長×寬×高)的玻璃水槽中(實際水體25 L),置于上述的光照培養(yǎng)箱中培養(yǎng)30 d。期間保持水溫17 ℃、鹽度30、光照強度45 μmol·m-2·s-1、光照周期12 L∶12 D、pH=8.3±0.2,使用充氧泵持續(xù)充氧保持DO≥6.0 mg·L-1,每2天使用虹吸法換水1次以保持水體清澈,燒杯在玻璃水槽中的位置每天隨機調(diào)換以避免光照條件差異的影響。經(jīng)30 d培養(yǎng)結束后統(tǒng)計各燒杯實生苗的存活數(shù),計算各種群的實生苗存活率;測定實生苗的葉長、葉寬、葉鞘長、莖長及根長等生長指標;隨后在每個種群的實生苗中隨機選取5株用于實生苗呼吸速率及光合速率的測定(黑白瓶法),隨機選取5株用于實生苗光合色素含量的測定(浸提法[23])。
1.3 數(shù)據(jù)分析
對不同種群鰻草種子和幼苗的所有指標進行單因素方差分析,若存在顯著差異再使用Duncan’s分析進行多重比較,結果用平均值±標準誤表示(Mean±SE),顯著性水平設置為p=0.05。對不同種群鰻草種子的形態(tài)學、生理學及萌發(fā)指標進行聚類分析及nMDS分析;對各種群鰻草種子的萌發(fā)率、幼苗存活率與種子形態(tài)學、生理學指標進行主成分分析(Principal component analysis,PCA),并對關鍵指標進行線性擬合。數(shù)據(jù)統(tǒng)計分析采用SPSS 25.0軟件進行,分析結果使用Origin 2018 C、R語言進行繪圖。
2 結果
2.1 不同種群鰻草種子的形態(tài)學特征
三個種群鰻草種子的形態(tài)學特征存在明顯差異。其中匯泉灣種群的種子規(guī)格最大,天鵝湖、俚島灣種群的種子規(guī)格較小,如表1所示。單因素方差分析顯示,匯泉灣鰻草種子的質(zhì)量顯著高于天鵝湖、俚島灣鰻草種子的質(zhì)量(p<0.05),是2個種群種子的1.8倍。同時,匯泉灣鰻草種子的長徑、短徑及最大橫截面積也顯著高于其他2個種群的鰻草種子(p<0.05),但3個種群種子的種形系數(shù)無顯著差異(p>0.05)。
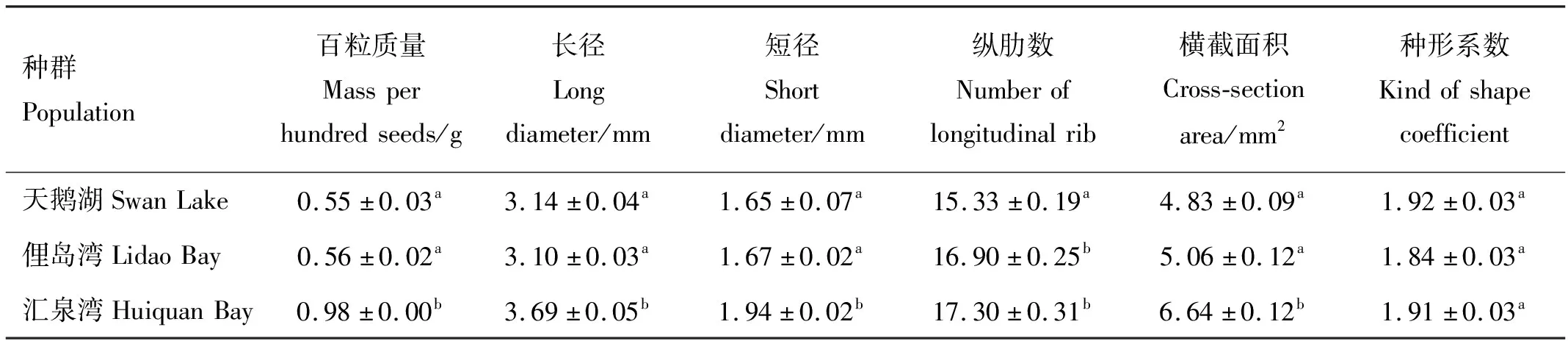
表1 不同種群鰻草種子的形態(tài)學特征
2.2 不同種群鰻草種子的貯存物質(zhì)含量
不同種群鰻草種子的內(nèi)部貯存物質(zhì)含量也不同。其中天鵝湖種群種子的營養(yǎng)物質(zhì)含量較高,匯泉灣種群種子較低,如圖2所示。三個種群鰻草種子的水分含量均高于40%。各種群種子內(nèi)部營養(yǎng)貯存物質(zhì)中均以總脂肪含量最少(≤1%),總糖含量最高,且不同種群間鰻草種子的總糖含量具有顯著差異(p<0.05),其中天鵝湖種群種子的總糖含量最高,平均達到62%,分別是俚島灣和匯泉灣種群種子的1.3和1.1倍。
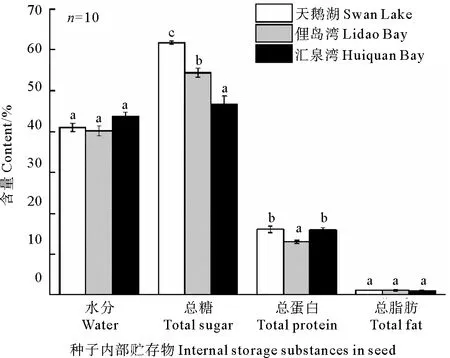
(不同字母表示不同種群間差異顯著(p<0.05)。Different letters indicate significant differences among the different populations(p<0.05).)
2.3 不同種群鰻草種子的萌發(fā)率與實生苗存活率
不同種群鰻草種子的萌發(fā)率、發(fā)芽指數(shù)及實生苗存活率均存在明顯差異,其中天鵝湖種群最高,如表2所示。天鵝湖種群鰻草的種子萌發(fā)率、發(fā)芽指數(shù)(GI)和實生苗存活率均顯著高于匯泉灣種群(p<0.05),分別是其1.5、2.2和2.9倍。各種群鰻草種子的平均萌發(fā)歷期(MTG)均在12 d左右,無顯著差異(p>0.05)。
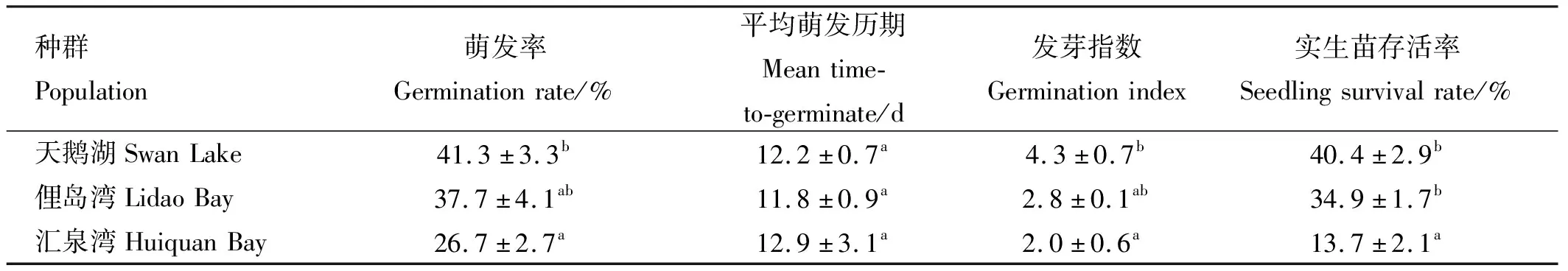
表2 不同種群鰻草種子萌發(fā)指標與實生苗存活率
2.4 不同種群鰻草種子的聚類分析
對山東半島不同種群鰻草種子的形態(tài)特征和內(nèi)部貯存物質(zhì)等指標進行聚類分析,發(fā)現(xiàn)不同種群間存在明顯區(qū)別,其中天鵝湖、俚島灣種群的鰻草種子較為接近,聚為一類;匯泉灣種群鰻草種子與其它2個種群存在較大差異,單獨聚為一類(見圖3(A))。nMDS分析進一步驗證了聚類分析的結果,應力值為0.039(見圖3(B))。
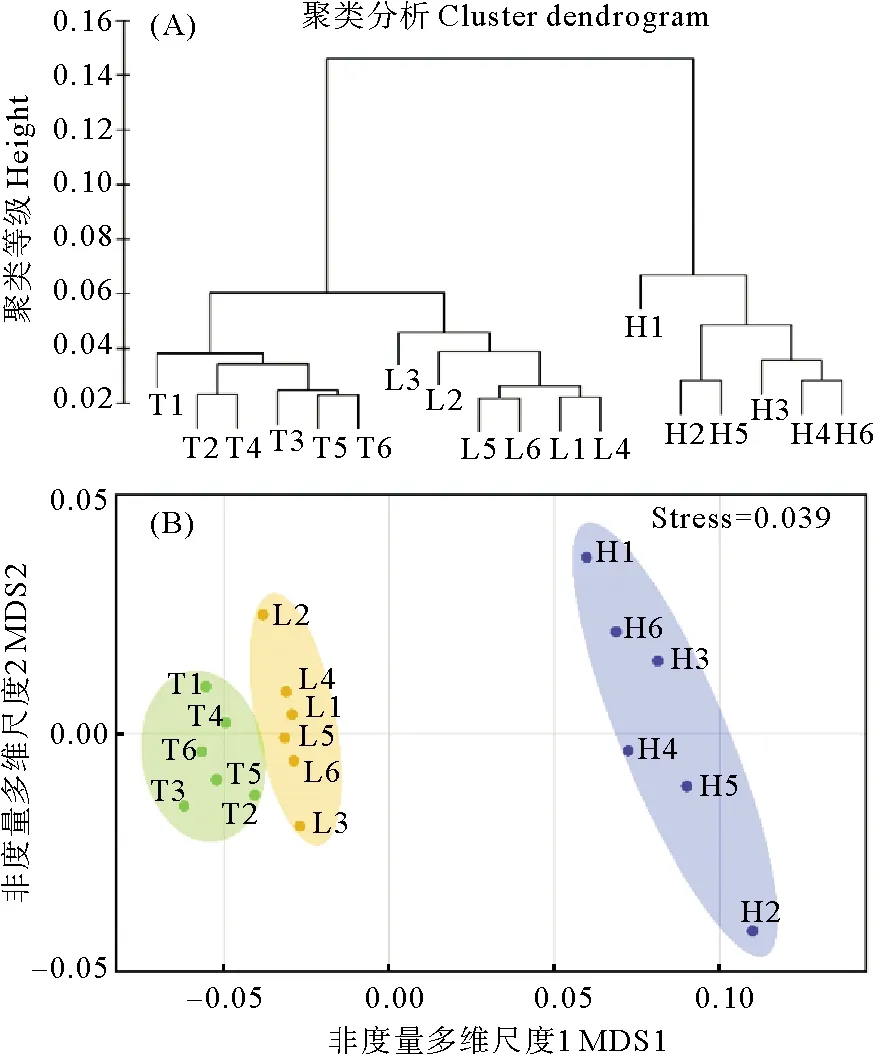
(圖中T1~T6表示天鵝湖種群;L1~L6表示俚島灣種群;H1~H6表示匯泉灣種群。T1~T6 in the figure represent the Swan Lake population; L1~L6 represent Lidao Bay population; H1~H6 represent Huiquan Bay population.)
2.5 不同種群鰻草種子萌發(fā)、實生苗存活與種子形態(tài)特征、內(nèi)部貯存物質(zhì)間的關系
主成分分析顯示,種子萌發(fā)率、發(fā)芽指數(shù)、實生苗存活率與種子總糖含量、種子長短徑、橫截面積、種子質(zhì)量、縱肋數(shù)、水分含量主要組成了第一排序軸,其中,以種子的萌發(fā)率、實生苗存活率、種子總糖含量和種子形態(tài)學這四個指標貢獻較大。如圖4所示,不同種群鰻草種子的萌發(fā)率、實生苗存活率與種子的總糖含量呈顯著的正相關,而與種子水分含量、形態(tài)學指標呈顯著的負相關。選取貢獻較大的種子總糖含量、種子質(zhì)量與種子萌發(fā)率、實生苗存活率進行曲線擬合顯示,種子總糖含量與種子萌發(fā)率、實生苗存活率呈顯著的線性正相關(p<0.05,見圖5A),而種子質(zhì)量與種子萌發(fā)率、實生苗存活率呈顯著的線性負相關(p<0.05,見圖 5B)。
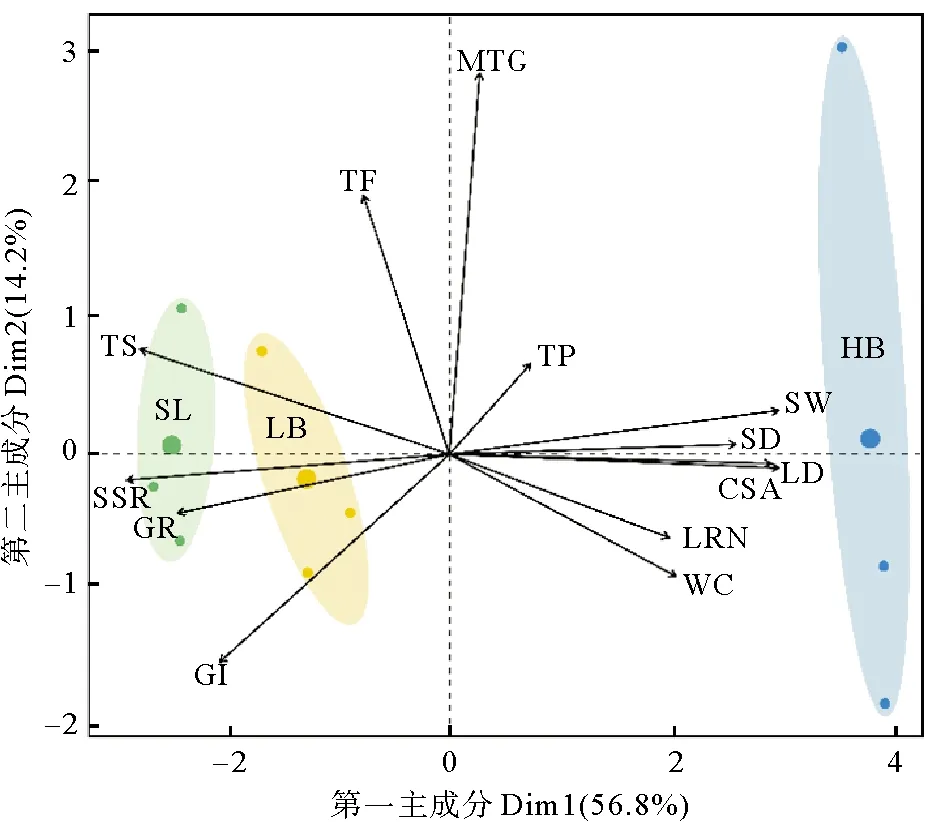
(圖中SL為天鵝湖種群,LB為俚島灣種群,HB為匯泉灣種群;GR為萌發(fā)率,SSR為實生幼苗存活率,GI為發(fā)芽指數(shù),TS為總糖,TF為總脂肪,MTG為平均萌發(fā)歷期,TP為總蛋白,SW為質(zhì)量,SD為短徑,LD為長徑,CSA為橫截面積,LRN為縱肋數(shù),WC為水分。SL in the figure is Swan Lake population, LB is Lidao Bay population, HB is Huiquan Bay population; GR is germination rate, SSR is seedling survival rate, GI is germination index, TS is total sugar, TF is total fat, MTG is mean time of germination, TP is total protein, SW is seed mass, SD is short diameter, LD is long diameter, CSA is cross-section area, LRN is number of longitudinal ribs, WC is water content.)
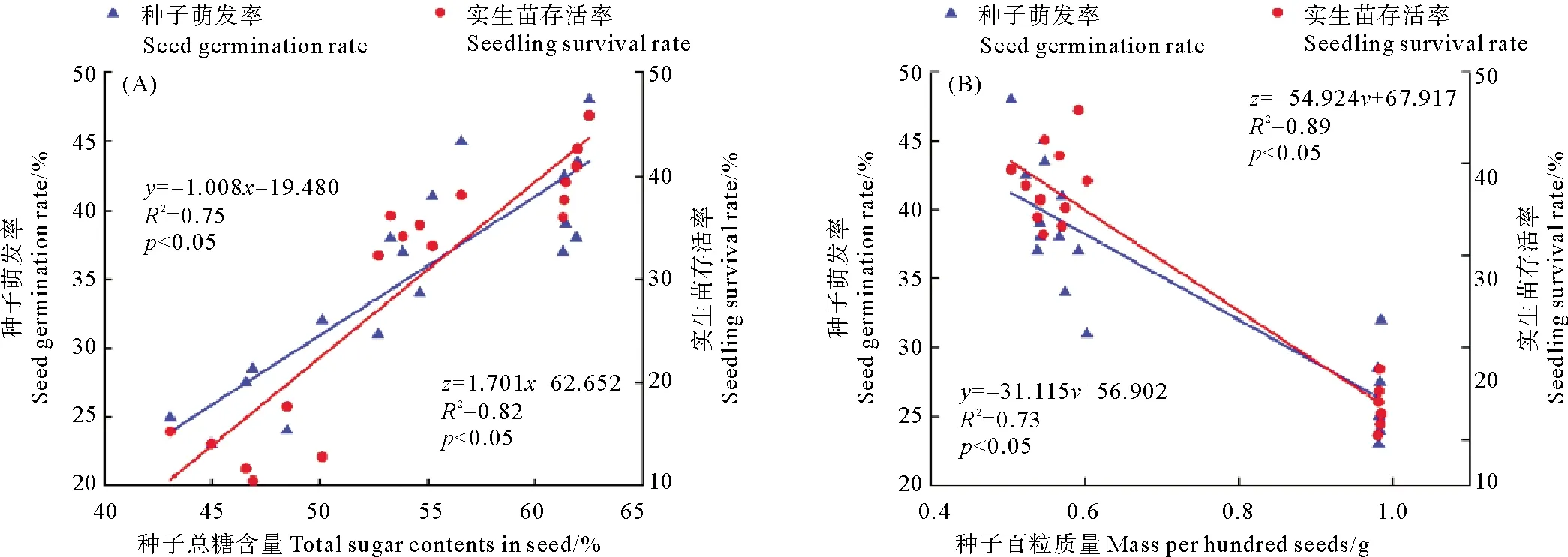
(圖中y表示種子萌發(fā)率;z表示實生苗存活率;x表示種子總糖含量;v表示種子百粒質(zhì)量。In the figure, y is seed germination rate; z is seedling survival rate; x is the total sugar content of seed; v is the mass per hundred seeds.)
2.6 不同種群鰻草實生苗的生長特性及光合生理
不同種群鰻草實生苗的生長存在差異。其中以天鵝湖種群的實生苗規(guī)格最大(見圖6)。單因素方差分析顯示,天鵝湖種群鰻草實生苗的葉寬、莖長和根長均顯著大于俚島灣鰻草實生苗(p<0.05),分別是其1.1、1.2和1.3倍;匯泉灣種群鰻草實生苗的莖長顯著小于天鵝湖種群實生苗的莖長(p<0.05),僅為天鵝湖種群實生苗莖長的84%。
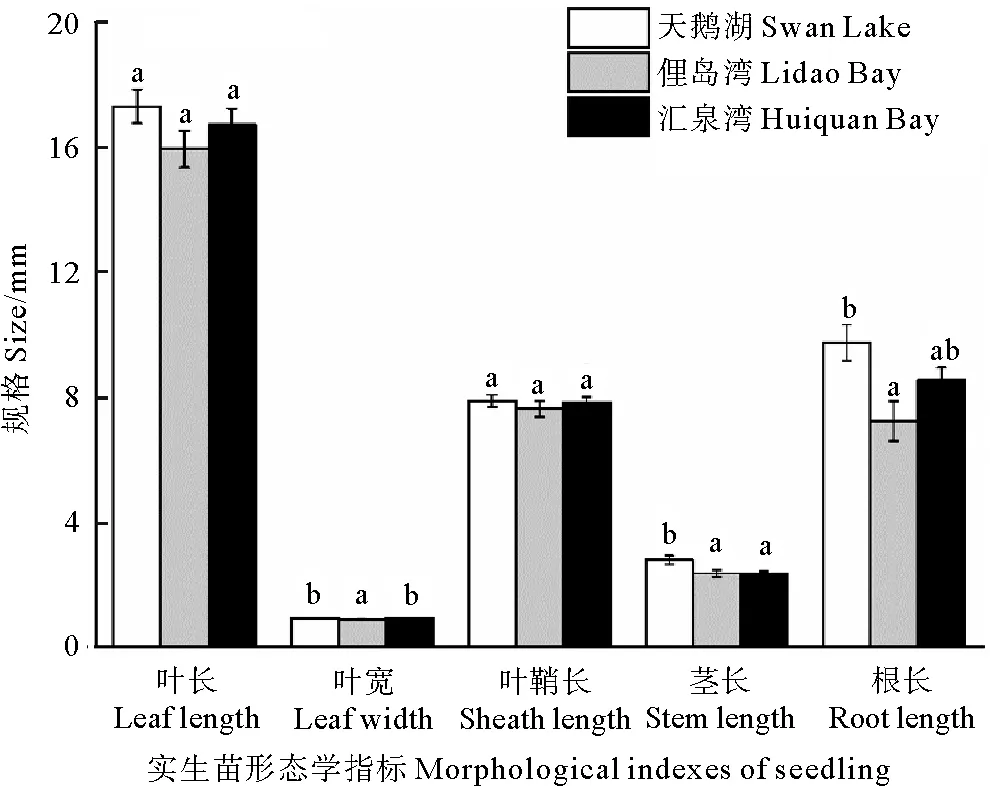
(樣本數(shù)量為10~40。不同字母表示不同種群間差異顯著(p<0.05)。The sample size is 10 to 40.Different letters indicate significant differences among the different populations(p<0.05).)
不同種群鰻草實生苗的光合色素含量和光合速率均存在差異。其中,天鵝湖種群實生苗的光合色素含量和光合速率最高,而俚島灣種群實生苗最低(見圖7和8)。單因素方差分析顯示,天鵝湖種群鰻草實生苗的光合色素含量均顯著高于俚島灣種群和匯泉灣種群實生苗(p<0.05),其總葉綠素含量達到6.5 mg·g-1,是俚島灣、匯泉灣實生苗葉綠素含量的1.5和1.2倍;天鵝湖鰻草實生苗的凈光合速率和呼吸速率也顯著高于俚島灣種群實生苗(p<0.05),是其1.2倍。
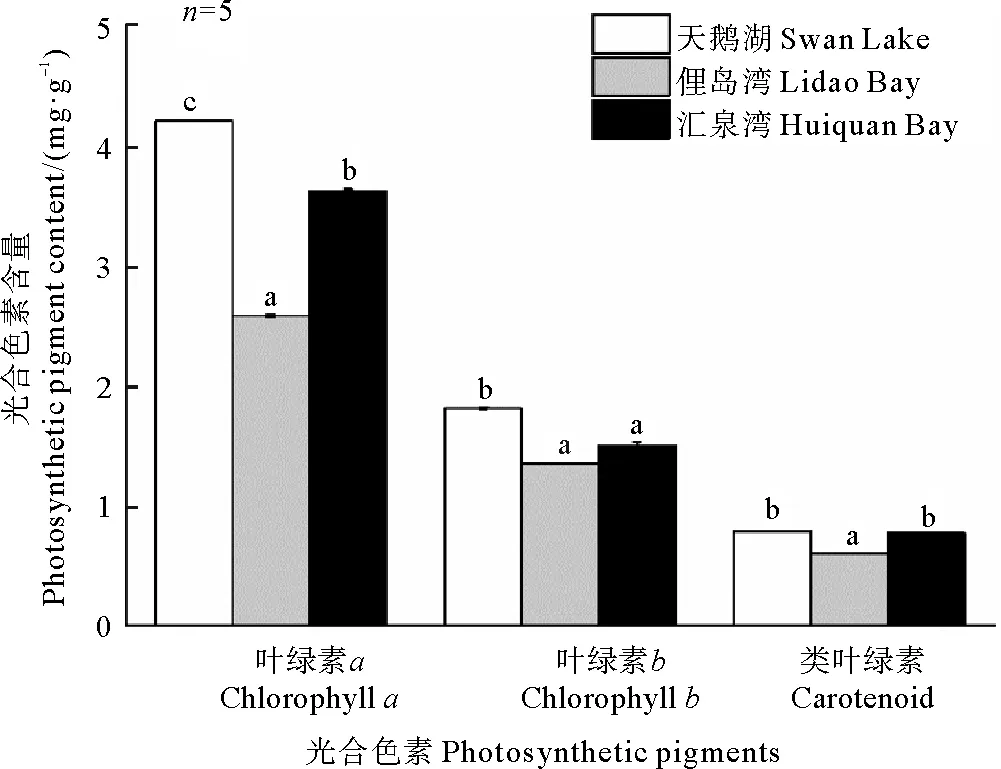
(不同字母表示不同種群間差異顯著(p<0.05)。Different letters indicate significant differences among the different populations(p<0.05).)
3 討論
3.1 不同種群海草的種子形態(tài)學特征及其環(huán)境基礎
同一植物種類的不同種群為更好地適應環(huán)境可能會發(fā)生遺傳變異,并將穩(wěn)定的性狀表現(xiàn)在種子的品質(zhì)和形態(tài)等方面,從而導致相同種類不同種群的種子形態(tài)學特征和內(nèi)部貯存物質(zhì)含量等存在顯著差異[24]。本研究發(fā)現(xiàn),匯泉灣鰻草種子的平均百粒質(zhì)量達0.98 g,長徑達3.69 mm,規(guī)格顯著大于天鵝湖和俚島灣種群的鰻草種子,與Xu等[25]研究結果相一致。Combs等[26]發(fā)現(xiàn),采集于美國北卡羅萊納州以北三個不同地點的鰻草種子大小顯著不同,鮮質(zhì)量在1.5~4.3 mg。對比其他研究發(fā)現(xiàn),本研究中匯泉灣種子比丹麥利姆海峽的鰻草種子(長徑3.4 mm)[27]和美國加利福尼亞灣北部的鰻草種子(長徑3.0~3.5 mm)[28]更大,說明不同種群間鰻草種子的形態(tài)大小因受不同地區(qū)影響而存在較大差異。
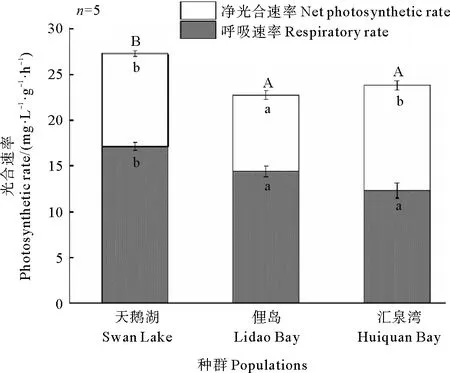
(不同字母表示不同種群間差異顯著(p<0.05)。Different letters indicate significant differences among the different populations(p<0.05).)
個體間種子規(guī)格的變異一般是遺傳型和環(huán)境因子共同作用的結果[29]。遺傳型包括細胞核遺傳、細胞質(zhì)遺傳和胚乳核遺傳[30]。Smith等[31]發(fā)現(xiàn),亞麻(Linumusitatissimum)植株母體的細胞質(zhì)直接影響種子大小, 并且這種影響可能延續(xù)到子二代和子三代。然而李淵等[32]發(fā)現(xiàn),愛爾蘭、日本、韓國和中國青島膠南、威海俚島灣、威海天鵝湖等地的鰻草種群間不存在地理隔離形成的基因分化,因此鰻草種子種群間的形態(tài)學差異可能主要是由不同的環(huán)境條件所造成。
研究表明,不同的緯度、種子分布深度及水動力等環(huán)境條件對植物個體間種子規(guī)格變異有明顯影響。張志翔等[33]發(fā)現(xiàn),低緯度地區(qū)的小桐子(Jatrophacurcas)種子比高緯度地區(qū)的種子規(guī)格更大;Wyllie等[34]發(fā)現(xiàn),美國部分海域鰻草種子的質(zhì)量隨緯度的降低而增大。本研究結果與這些研究相類似,三個種群鰻草種子的質(zhì)量隨天鵝湖、俚島灣、匯泉灣緯度的降低而逐漸增大。種子分布深度也是影響種子規(guī)格的關鍵因素。彭鴻嘉等[35]發(fā)現(xiàn),當種子處于較深底質(zhì)時,較大的種子因儲存更多的能量而形成更長的莖,從而順利萌發(fā)并存活。匯泉灣底質(zhì)為砂質(zhì)[36],較天鵝湖的泥沙底質(zhì)[22]和俚島灣的礫沙底質(zhì)[37]更松散,鰻草種子更易被深埋,因此,為適應環(huán)境并保證實生苗的順利萌出,匯泉灣種子的個體更大。此外,Orth等[38]發(fā)現(xiàn),海草植株在水流湍急的水域中會產(chǎn)生質(zhì)量較大的種子,以便種子能快速沉降,減少被帶至不宜生長區(qū)域的幾率。匯泉灣水深7 m[36],該海域的鰻草植株通常生長于水流較急的潮下帶,只有質(zhì)量更大的鰻草種子才能保留下來;而天鵝湖、俚島灣的鰻草植株生活在2 m以內(nèi)水流較緩的潮下帶[37],種子可以很快沉降,因而這兩個地區(qū)的鰻草種子較小。
3.2 不同種群海草種子的萌發(fā)特性及其關鍵影響因素
種子萌發(fā)是實生苗建成的前提,進而影響整個群體的形成。本研究發(fā)現(xiàn),天鵝湖種群鰻草種子萌發(fā)率和實生苗存活率均達40%以上,顯著高于匯泉灣種群,分別是其1.6和3.0倍。此外,天鵝湖實生苗規(guī)格、光合色素含量和光合速率均為最高值,顯著高于俚島灣種群。Tanner等[39]發(fā)現(xiàn),收集于美國馬里蘭州和弗吉尼亞州的鰻草種子的室內(nèi)萌發(fā)率為33%,實生苗存活率為20%。Cabaco等[40]將大西洋西部四個不同種群鰻草種子進行室內(nèi)萌發(fā)實驗,發(fā)現(xiàn)僅有兩個種群的鰻草種子萌發(fā),且最高萌發(fā)率僅5%,在萌發(fā)的種子中,僅15%的種子能實現(xiàn)幼苗建成。綜合對比發(fā)現(xiàn),本研究地區(qū)及其他不同地區(qū)種群的鰻草種子萌發(fā)特性和實生苗存活存在顯著差異,這可能與種子規(guī)格及營養(yǎng)貯存物質(zhì)的含量有關[11-13, 41]。
本研究發(fā)現(xiàn),鰻草種子萌發(fā)率、實生苗存活率與種子質(zhì)量、長徑、短徑等形態(tài)學指標呈負相關,該結果與其他學者的研究存在不同。Jarvis等[42]認為,大規(guī)格的鰻草種子具有更高的活力、更短的萌發(fā)時間和更高的萌發(fā)率。Combs等[26]也發(fā)現(xiàn),與小規(guī)格的種子相比,大規(guī)格的鰻草種子可以獲得更多的能量資源,從而提高種子萌發(fā)率。而J?rgensen等[27]發(fā)現(xiàn),鰻草種子的規(guī)格對出苗率和出苗時間無明顯影響,但質(zhì)量較大的種子壽命更長,產(chǎn)生的幼苗生物量顯著提高。這些結果表明,在種子萌發(fā)期和幼苗初始發(fā)育過程中代謝貯存物的調(diào)動及其含量可能起到重要作用。
在本研究中,鰻草種子營養(yǎng)貯存物含量為總糖>總蛋白>總脂肪,其中各種群種子總糖和總蛋白含量存在顯著差異,并以天鵝湖種群最高,分別達到62%和16%。對比發(fā)現(xiàn),天鵝湖種子的總糖含量高于日本岡山Hinase灣的鰻草種子(粗糖含量約48%)[43]及美國加利福尼亞Hadiona瀉湖的鰻草種子(糖含量約51%)[44]。此外,盡管俚島灣的鰻草種子總蛋白含量最低,僅13%,但仍高于日本Otsuchi灣的鰻草種子(蛋白含量約9%)。Belzunce等[45]認為,大洋波喜蕩草(Posidoniaoceanica)種子的內(nèi)部營養(yǎng)貯存物質(zhì)越充足,其幼苗存活及初始生長發(fā)育越好。本研究發(fā)現(xiàn),鰻草種子的總糖含量越高,種子萌發(fā)率和實生苗存活率也越高,實生苗生長更好。
通過主成分分析和曲線擬合發(fā)現(xiàn),種子總糖對鰻草種子的萌發(fā)與實生苗生長起主要作用,鰻草種子萌發(fā)率及實生苗存活率與種子總糖含量呈顯著的線性正相關,糖含量較高的種子具備更高的萌發(fā)力。黃升謀等[46]發(fā)現(xiàn),小麥(Triticumaestivum)種子萌發(fā)前期需要消耗大量的糖,以保證萌發(fā)時有足夠能量及結構物質(zhì)來合成新的細胞和細胞器。蔣敏明等[47]發(fā)現(xiàn),水稻(Oryzasativa)種子總淀粉積累量與發(fā)芽率、發(fā)芽指數(shù)及活力指數(shù)顯著相關。對于海洋植物種子的報道中也有類似發(fā)現(xiàn)。Kuo和den Hartog[48]指出,鰻草種子的下胚軸僅含有少量蛋白質(zhì),但淀粉含量卻很高,淀粉在種子萌發(fā)和幼苗發(fā)育過程中起主要供能作用。Taylor[49]也發(fā)現(xiàn),種子總糖是鰻草種子萌發(fā)早期主要的能量來源,在種皮破裂和下胚軸伸長過程中起關鍵作用,充足的糖含量才能保證種子萌發(fā)。鰻草種子在萌發(fā)過程中呼吸作用逐漸增強,作為鰻草種子的主要營養(yǎng)貯存物,充足的糖能為種子提供足夠的呼吸作用底料,以支持萌發(fā)過程所需的大量能量,使種子萌發(fā)成功的幾率大大提升。此外,糖也為幼苗形成初期提供必須的能量及養(yǎng)料。Venable[50]發(fā)現(xiàn),種子的營養(yǎng)貯存物質(zhì)所占比例越大,幼苗可獲取的能量就越多、存活率越高。孫德蘭等[51]發(fā)現(xiàn),在蓮(Arabidopsisthaliana)幼苗未具備自養(yǎng)能力之前,其生長主要依靠種子內(nèi)部貯存的淀粉粒,淀粉粒不僅提供能量,其包含的DNA也為幼苗形態(tài)建成提供了核苷酸等原料。Sugiura等[52]發(fā)現(xiàn),鰻草種子內(nèi)部的淀粉降解產(chǎn)物不僅是胚芽突破種皮的主要能量來源,也是實生幼苗發(fā)育的必需代謝產(chǎn)物,對鰻草實生苗的建成與生長起決定性作用。鰻草種子糖含量越高,供給實生幼苗生長的能量及合成原料就越充足,幼苗就會產(chǎn)生更長的根系,具有更高的水分和養(yǎng)分吸收能力,從而利于植株的建成[53]。
本研究還發(fā)現(xiàn),種子萌發(fā)率及實生苗存活率與種子質(zhì)量存在顯著的線性負相關關系。宗文杰等[54]發(fā)現(xiàn),在青藏高原地區(qū),菊科植物小規(guī)格種子具有更快的萌發(fā)速率和更高的萌發(fā)率,由此來適應高寒草甸環(huán)境。劉振恒等[55]發(fā)現(xiàn),較小的種子可以通過迅速萌發(fā)的方式,優(yōu)先占據(jù)資源及生存空間。本研究中天鵝湖、俚島灣的年水溫變化幅度較大,鰻草葉片通常于冬季脫落。在這種環(huán)境下,規(guī)格較小的種子更快萌發(fā),從而獲得競爭優(yōu)勢。
4 結語
探明不同種群鰻草種子的形態(tài)學特征及萌發(fā)特性,是實現(xiàn)優(yōu)質(zhì)種源篩選及成功開展海草種子種植的首要關鍵。本研究結果發(fā)現(xiàn),山東半島不同種群的鰻草種子在形態(tài)學特征和萌發(fā)特性上存在顯著差異,種子萌發(fā)及幼苗建成的關鍵在于種子內(nèi)部的糖含量。因此,在篩選種源時,可挑選糖含量高的鰻草種子作為海草床修復的材料。由于種子萌發(fā)與幼苗生長不僅受內(nèi)在因素的影響,還受到外界多種環(huán)境因素的影響,關于不同種群所處的環(huán)境條件與種子內(nèi)部營養(yǎng)物質(zhì)含量對種子萌發(fā)及幼苗生長的影響是否存在交互作用等科學問題,還需進一步探討。
