紫花苜蓿根際土壤細菌群落對腐熟牛糞響應
2022-03-28 04:51:06李文辰趙金波張智慧
草地學報 2022年3期
關鍵詞:物種
王 芳, 李 偉, 劉 鑫, 李文辰, 趙金波, 張智慧, 楊 曌
(1.齊齊哈爾大學生命科學與農林學院, 抗性基因工程與寒地生物多樣性保護黑龍江省重點實驗室, 黑龍江 齊齊哈爾 161006;2. 黑龍江省農業科學院畜牧獸醫分院, 黑龍江 齊齊哈爾 161005; 3.黑龍江省牧草育種與種質資源利用工程技術研究中心,黑龍江 齊齊哈爾 161005)
微生物是土壤生態系統中的重要組成部分,驅動著土壤系統中各種生物進程。微生物對環境變化敏感,施肥顯著影響土壤微生物種群特性,長期施用氮、磷、鉀化肥可降低土壤微生物活性及多樣性[1]。添加糞肥、植物殘體等有機肥可改善土壤肥力、提高土壤有機質含量,穩定土壤微生物系統[2-4]。不同養分含量的有機肥對土壤微生物主要類群的影響不同。新鮮豬糞或發酵豬糞可以明顯提高水稻(Oryzasativa)根際土壤微生物的物種豐富度和群落均勻度,加快有機物質在土壤中的分解轉化[5]。蚯蚓糞會增加土壤有益微生物種類和數量,提高土壤細菌多樣性,增強土壤酶活[6],對番茄(Solanumlycopersicum)連作土壤細菌群落結構的調節效果優于稻草、雞糞、牛糞有機物料[7],但有研究表明牛糞可以對放線菌優勢群落產生顯著影響[8]。
土壤細菌作為土壤肥力和養分循環的驅動者,在維持農業生態系統的生產力和穩定性方面具有不可替代的地位[9-10]。細菌群落對土壤條件包括水分、溫度、pH值、土壤類型和養分有效性等變化的反應快于其它化學或物理性質的反應[11],細菌群落結構和多樣性可以作為評估農業措施對土壤質量影響的重要指標[12-13]。研究發現有機肥處理的土壤中細菌的優勢種群明顯不同于單施化肥和不施肥[14]。
紫花苜蓿(Medicagosativa)為多年生豆科植物,含有豐富的營養成分,被譽為“牧草之王”,因其較高的營養價值和較強的地域適應性,在我國北方畜牧業發展中具有舉足輕重的地位。為了滿足苜蓿正常生長對養分的需求,常需通過施肥來補充土壤養分。牛糞是畜牧產業的廢棄物,將其作為有機肥料用于作物種植,形成種養有機結合的循環模式,不但解決了糞便污染和化肥過量使用的問題,而且可以改良土壤、培肥地力、提高作物產量及品質,實現經濟及生態效益雙豐收。本研究基于16S rRNA基因高通量測序技術,探索不同添加比例腐熟牛糞對苜蓿根際土壤細菌豐度、多樣性及群落結構組成的影響,旨在為腐熟牛糞在農業中的應用及施肥策略制定提供科學依據。
1 材料與方法
1.1 試驗設計與樣品采集
試驗于2020年 4月至9月進行。供試紫花苜蓿品種為‘龍牧801’,土壤采自黑龍江農業科學院畜牧獸醫科研試驗地(齊齊哈爾市富拉爾基區)。采用盆栽(盆缽25 cm ×30 cm),每盆均播種50粒種子。4種質量百分比添加腐熟牛糞,① O組(零添加組,0% 添加量);② D1組(3% 添加量);③ D2組(6% 添加量);④ D3組(9% 添加量),分別在播種后(苜蓿分枝期)的30 d(T1),60 d(T2),90 d(T3)取樣,共12個處理,每個處理5次重復。此外,在添加牛糞前,取全土(不添加牛糞,WT)和全糞(不添加土壤,WF)兩個樣品,以了解未經處理及種植的土壤及腐熟牛糞細菌群落情況。采用抖落法收集苜蓿根際土壤,混勻后分裝在聚乙烯袋中,-80℃保存,作為總DNA的提取樣本。
1.2 土壤總DNA提取、PCR擴增及基因測序
每個樣品稱取0.5 g,根據Mo BIO Power Soil DNA Kit說明書提取總DNA,1% 瓊脂糖凝膠電泳檢測DNA完整性。以稀釋后的基因組DNA為PCR模板,利用細菌通用性引物338F(5′-ACTCCTACGGGAGGCAGCA-3′),806R(5′-GGACTACHVGGGTWTCTAAT-3′)擴增細菌16S rRNA V3-V4可變區。擴增體系為 25 μL,5×FastPfu緩沖液 4 μL,2 μL 2.5 mmol·L-1dNTPs,0.8 μL引物(5 μmol·L-1),0.4 μL FastPfu聚合酶;10 ng DNA模板。PCR擴增程序為:95℃預變性 2 min,30 個循環(95℃變性 30 s,55℃退火 30 s,72℃延伸 30 s),最后 72℃延伸 10 min。PCR產物經1.2% 的瓊脂糖凝膠電泳,利用SanPrep柱式DNA膠回收試劑盒(生工生物工程,上海)進行純化。獲得PCR產物送至派森諾生物科技有限公司(上海)進行Illumina-Miseq高通量測序。原始數據提交至NCBI的Sequence Read Archive (SAR)數據庫,Accession號碼:PRJNA741810。
1.3 原始數據處理
采用DADA2方法[15]對序列進行質控、去噪(Denoise)、拼接和去嵌合體,提高后續序列融合比率。利用Vsearch方法[16]聚類獲得操作分類單元(Operational taxonomic units,OTU),統計長度分布情況,檢查目的片段長度范圍及異常長度序列情況。隨機抽取序列,構建稀釋性曲線。
1.4 物種分類學注釋及物種組成分析
利用QIIME2軟件,選用Greengenes數據庫(http:// greengenes.secondgenome.com/)進行細菌16S rRNA基因注釋。使用QIIME2(2019.4)及自編perl腳本,依據序列物種分類學注釋的結果以及選擇的樣品,統計這些樣本的物種注釋結果中域、門、綱、目、科、屬、種七個分類水平各自含有的分類單元數量,用柱狀圖呈現分析結果。
1.5 Alpha多樣性分析
利用派森諾基因云平臺(https://www.genescloud.cn/)進行OTU豐度和Alpha多樣分析,Chao1和Observed species指數表征豐富度,Shannon和Simpson指數表征多樣性。
1.6 Beta多樣性分析
使用抽平后OTU表,基于Bray-Curtis距離矩陣進行PCoA(Principal coordinate analysis)分析,用QIIME2使分析結果可視化,R腳本輸出樣本點的PCoA坐標,繪制二維散點圖。
1.7 數據統計分析及制圖
使用IBM SPSS Statistics 19.0 統計分析軟件對Alpha多樣性指標、樣本多樣性數據進行二因素方差分析(WF和WT兩個樣本不參加方差分析),多重比較采用Duncan檢驗法,差異顯著性水平為P<0.05。
2 結果與分析
2.1 測序數據分析
將測序平臺得到的原始數據去除低質量、去噪、去嵌合體、拼接后,供試樣本共獲得984 200條高質量序列,去除singleton后的序列量是942 003條,總堿基數為392 582 624 bp,堿基數分布在403~442 bp的序列占序列總數的99.98%,平均序列長度為 417 bp。14個樣品的稀釋性曲線均趨于平坦(圖1),表明測序數據接近飽和,測序深度合理,更多的數據量對發現新的OTU貢獻率較小。同時,稀釋曲線在一定程度上也可以反應在一系列給定的測序深度下,可能包含的物種總數及其中每個物種的相對豐度。在相同的測序深度下,比較不同樣本中OTU數的多少,在一定程度上衡量每個樣本的多樣性高低。由圖1所示,WT樣本的多樣性最高,其次為O組、D1,D2,D3組,WF樣本的多樣性最低。

圖1 樣品稀釋曲線Fig.1 Rarefaction curves for samples
2.2 物種分類學注釋
將獲得OTU從門到種進行物種分類學注釋分析。所有供試土壤樣品共注釋到細菌分類85門,335綱,362目,1 828科,4 441屬,368種。樣品WF在門和屬水平上注釋到的OTU數目在所有樣品中均為最低,分別為16和1 807,在種水平上為最高368個。WT在門水平上注釋到的85個OTU,在所有樣品中為最高。在屬水平上,OT3注釋到的OTU數目最多,為4 441個;在種水平上注釋到的OTU數目為最低182個(圖2)

圖2 各樣品在不同分類水平上的OTU注釋數目Fig.2 OTU annotation number at varied classification level of different samples
2.3 物種分類學組成分析
由圖3所示,樣品中鑒定到的OTU主要分布在以下10個門:放線菌門(Actinobacteria)、變形菌門(Proteobacteria)、厚壁菌門(Firmicutes)、綠彎菌門(Chloroflexi)、酸桿菌門(Acidobacteria)、擬桿菌門(Bacteroidetes)、芽單胞菌門(Gemmatimonadetes)、Patescibacteria、疣微菌門(Verrucomicrobia)、消化螺旋菌門(Nitrospirae)。其中,Actinobacteria最為豐富,在14個樣品中相對豐度為55.9%~32.2%;其次為Proteobacteria,相對豐度為36.9%~20.5%。
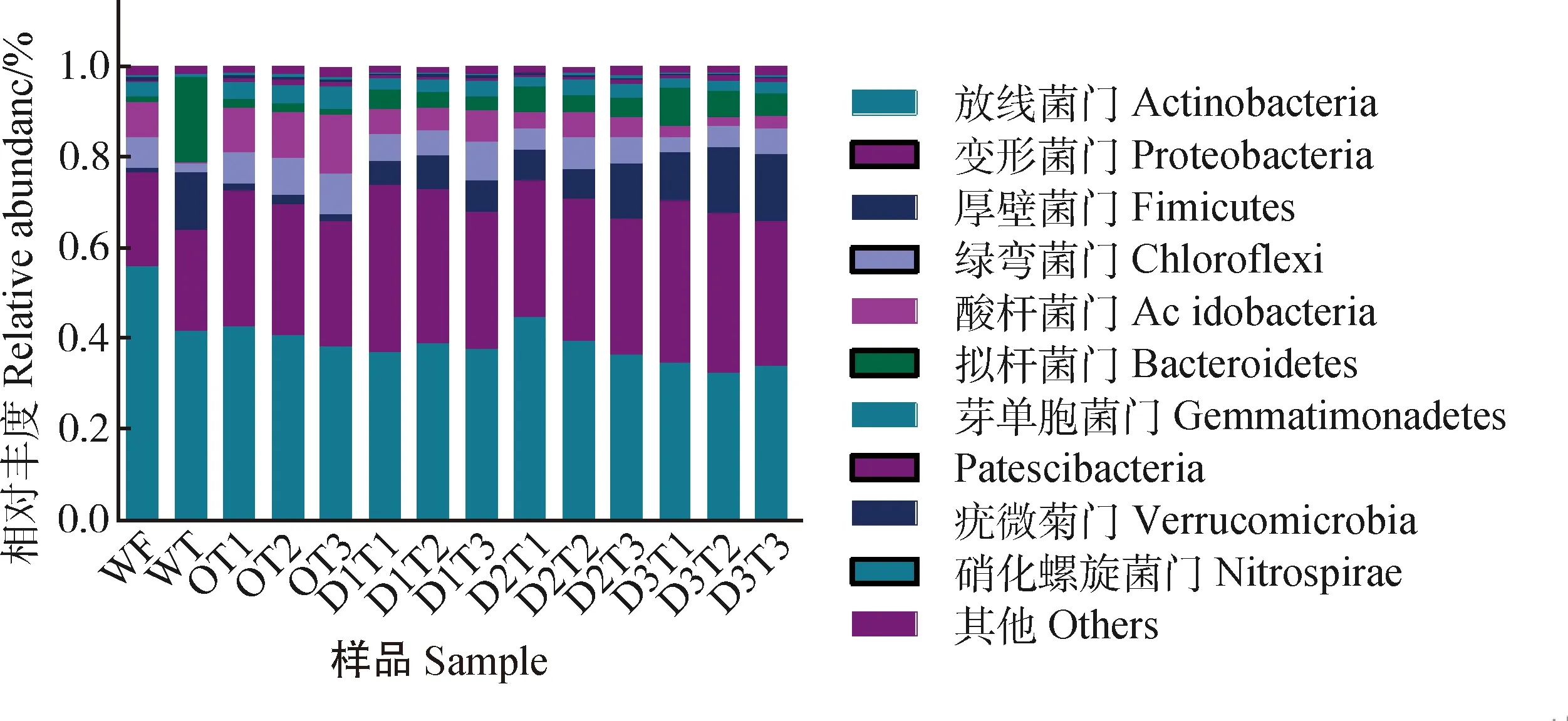
圖3 樣品在門水平的群落相對豐度Fig.3 Relative abundance of samples bacterial diversity at phylum level
Actinobacteria和Proteobacteria也是樣品WT與WF的優勢菌門,Proteobacteria,Firmicutes和Bacteroidetes在WF種的相對豐度分別高于在WT中的豐度。與O組相比,添加牛糞后D1,D2和D3組的Firmicutes和Bacteroidetes相對豐度顯著增加(P< 0.05),Proteobacteria在D3組達到顯著性增加(P< 0.05);Chloroflexi,Acidobacteria,Gemmatimonadetes,Verrucomicrobia,Nitrospirae豐度呈顯著性下降(P< 0.05),Actinobacteria在D3組下降顯著(P< 0.05)。Patescibacteria變化不顯著(表1)。
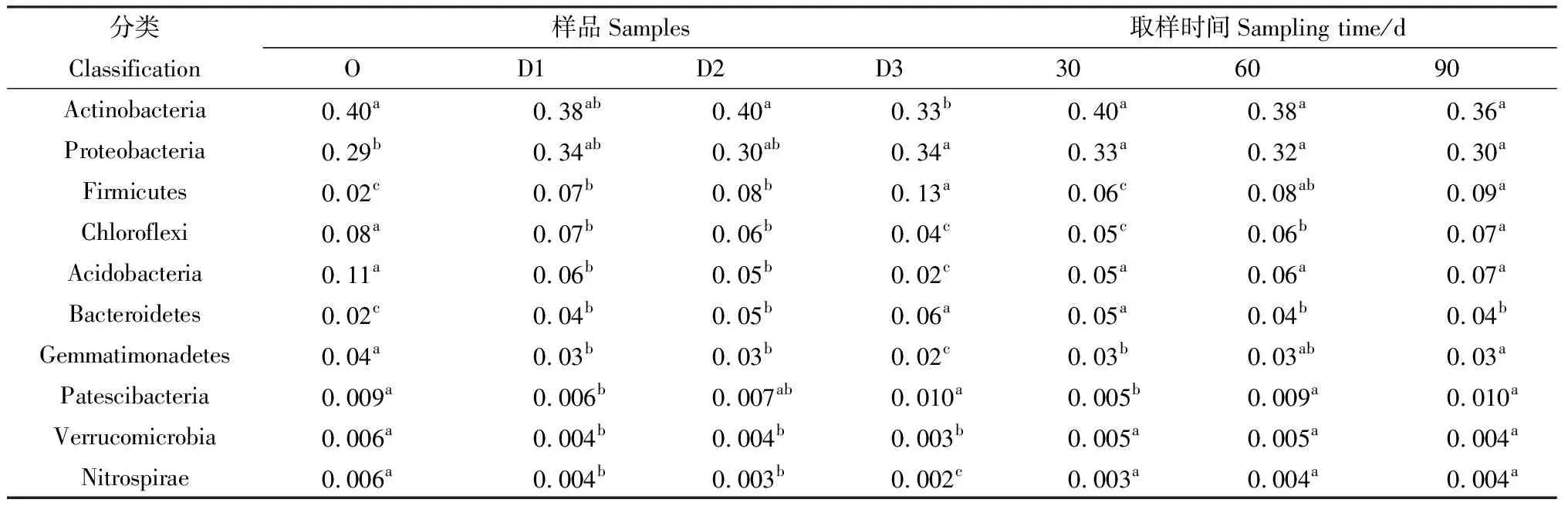
表1 不同添加量及取樣時間下各樣品在門水平的相對豐度Table 1 Relative abundance of bacterial taxa classified to the phylum level at different adding amount and sampling time
隨著取樣時間的延伸,Firmicutes,Chloroflexi,Gemmatimonadetes,Patescibacteria的相對豐度呈現顯著增加(P< 0.05),Bacteroidetes呈現下降趨勢。
在屬分類水平上,豐度較低的稀有種群占78.03%~84.67%,表明土壤中存在大量待發掘的細菌物種。為了便于分析鑒定到的優勢菌群對添加牛糞及取樣時間的響應,將相對豐度位于前10位的優勢菌屬作圖。如圖4所示,樣品WT及WF具有不同的優勢菌屬,WT的優勢菌屬有Subgroup_6(酸桿菌)、鞘氨醇單胞菌屬(Sphingomonas)、假諾卡氏菌屬(Pseudonocardia)、芽球菌屬(Blastococcus)、KD4-96(綠彎菌)、67-14(放線菌);WF中具有相對豐度較高的芽孢桿菌屬(Bacillus)、馬杜拉放線菌屬(Actinomadura)、BIrii41(厚壁菌)。
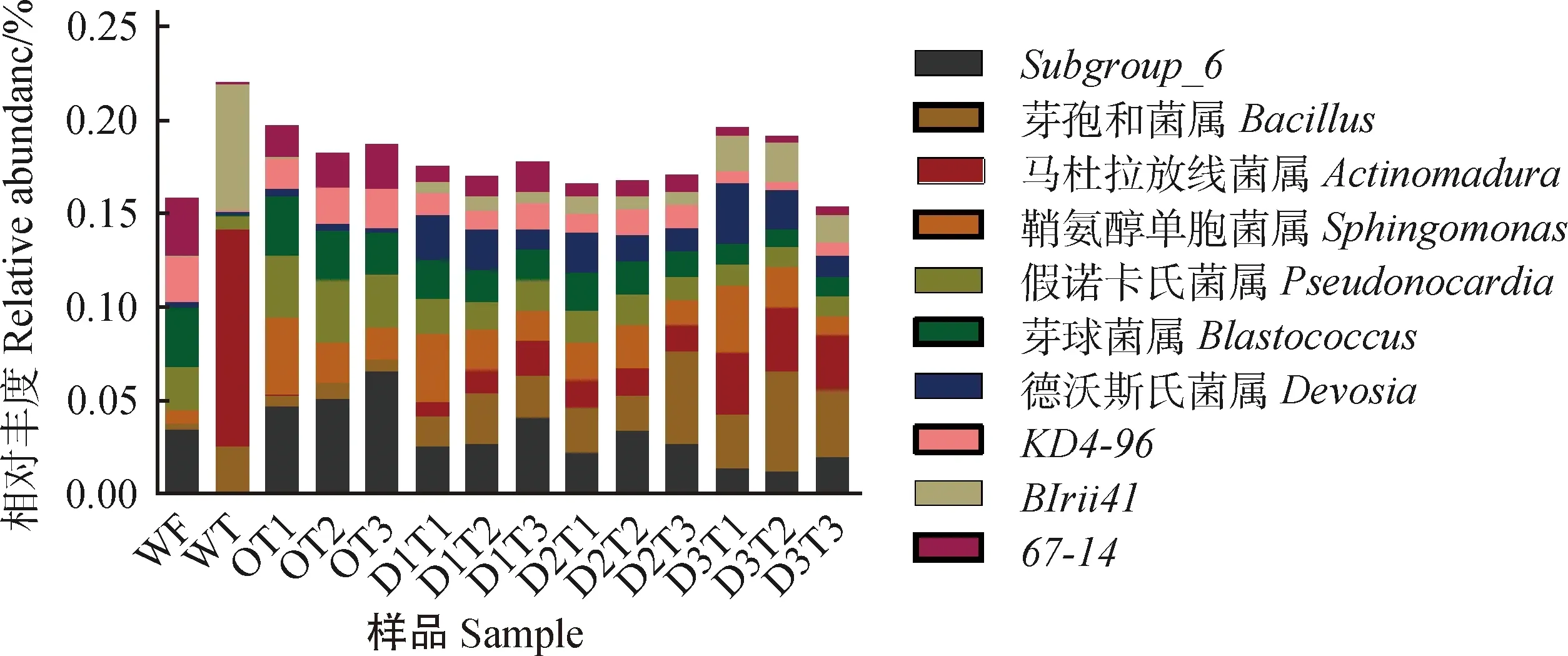
圖4 樣品在屬水平的群落相對豐度Fig.4 Relative abundance of samples bacterial diversity at genus level
Bacillus和Actinomadura的相對豐度均隨著牛糞添加比例的增加而升高,且三個水平添加組顯著高于O組(P< 0.05);德沃斯氏菌屬(Devosia)與BIrii41在三個添加組中的豐度也均高于O組。Subgroup_6,Pseudonocardia,Blastococcus,67-14,KD4-96的相對豐度均隨著添加比例的增加呈顯著下降,Sphingomonas豐度在添加至D2組時顯著低于O組(表3)(P< 0.05)。
從取樣時間上看,Sphingomonas和Devosia的相對豐度受取樣時間影響顯著,隨取樣時間延長呈顯著下降(P< 0.05)。Blastococcus相對豐度在30 d和90 d呈顯著性差異(P< 0.05)。Pseudonocardia,Actinomadura和綠彎菌KD4-96的相對豐度在三個取樣時間上沒有顯著差異。Subgroup_6,Bacillus,BIrii41和67-14的相對豐度隨取樣時間延長呈增加趨勢。

表2 不同添加量及取樣時間樣品在屬水平的相對豐度Table 2 Relative abundance of bacterial taxa classified to the genus level at different adding amount and sampling time
2.4 Alpha多樣性分析
與O組相比,不同添加比例對Alpha多樣性指數Chao1,Observed_species,Shannon,Simpson均產生顯著性的影響(P< 0.05)(表3)。O組的四個指數均為最高,隨著土壤中牛糞含量的增加,四個指數均呈明顯下降趨勢,表明細菌群落的豐富度及多樣性在下降(P<0.05)。四個指數在D1組和D2組之間差異不顯著。

表3 不同添加比例細菌Alpha多樣性指數Table 3 Bacterial Alpha diversity indices at different adding amount
2.5 Beta多樣性的主坐標軸分析
基于樣本間Bray-Curtis距離,PCoA主坐標分析樣本群落之間的相似度和差異性。由圖5所示,主坐標軸PCo1可解釋全部樣品細菌群落豐度和多樣性方差的22.4%,主坐標軸PCo2可解釋細菌群落豐度和多樣性方差的的11.1%。不同樣本處理組群落組成及多樣性之間明顯分離,可以分為3大區域:樣本D3組(第四象限)主要分布在主坐標PCo1附近,說明D3組苜蓿根際土壤細菌群落結構受PCo1影響明顯。第二區域主要分布在PCo2附近(第三象限),即O組在PCo2主軸上分開,說明O組根際土壤細菌群落結構主要受PCo2影響。第三區域為D1組和D2組(第一象限),距離PCo1及PCo2上均較遠,D1組內樣本間距離也較遠,表明D1組的細菌群落結構差異大于D2組。
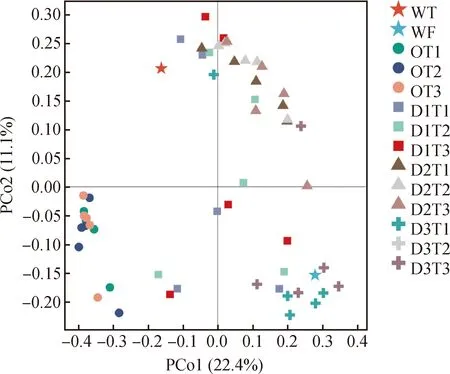
圖5 基于Bray-Curtis差異PCoA分析土壤樣品細菌群落Fig.5 PCoA analysis with Bray-Curtis dissimilarity of the bacterial communities between samples
2.6 樣本間物種差異分析
為研究不同樣本間共有及獨有的物種情況,使用OTU豐度表制作花瓣圖進行群落分析。如圖6所示,不同比例牛糞在添加30 d,OT1組具有最多的OTU分布數目,其次為D3T1組,D1T1組為最少,四組一共有OTU 45 004個,共有OTU數目為1 880個,約占4.2%。添加60 d,OT2組具有最多的OTU分布數目,其次為D1T2組,D3T2組為最少,四組一共有47 737個,共有的OTU數目為1 464個,約占3.1%。添加90 d,OT3組具有最多的OTU分布數目,其次為D1T3組,D3T3組為最少,四組一共有OTU 48 321個,共有的OTU數目為1 331個,約占2.7%。從時間看,隨著取樣時間的延伸,D1組(添加3%)和D2組(添加6%)的OTU數目呈增加趨勢,D3組(添加9%)OTU呈下降趨勢;四組樣本共有的OTU數目比例下降,說明物種的多樣性呈增加趨勢。
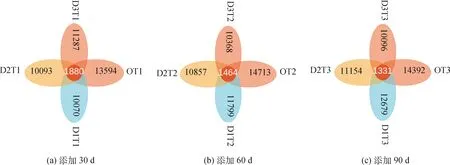
圖6 不同取樣時間下樣本間共有和獨有OTU數目的花瓣圖Fig.6 The petals figure based on common or unique OTU number among treatments at sampling time
2.7 物種組成差異熱圖
為了進一步比較樣本間的物種組成差異,將平均豐度前20位的屬的豐度數據進行聚類分析(圖7)。如圖7所示,Lysobacter,BIrii41,Devosia,Steroidobacter的豐度在WF與WT及O組間相似,在添加牛糞后豐度增加。Cellvibrio(纖維菌屬)、Planifilum(直絲菌屬)、Bacillus、Saccharomonospora(糖單胞菌屬)豐度在WF,WT及O組差異不明顯,在土壤中添加牛糞后,明顯高于WF,WT,O組,并隨著牛糞含量的增加呈增加趨勢,在D3組處理中均達到最高峰,表明混合土壤會促進這些菌屬的豐度。Actinoplanes(游動放線菌屬),67-14,Blastococcus,KD4-96,Skermanella(紅弧菌屬),RB41,Subgroup_6,Pseudonocardia(假諾卡氏菌屬)的豐度在WT中含量最高,隨著牛糞比例的增加明顯下降,在WF組豐度最低。Actinomadura和Haloactinopolyspora在WF中高豐度的細菌。
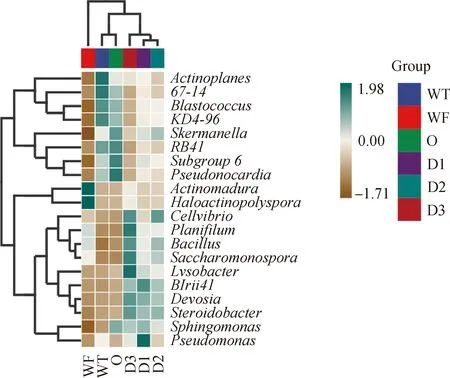
圖7 屬水平物種組成差異熱圖Fig.7 Heatmap tree for different treatments cluster analysis at genera level注:標尺顏色深淺表示樣本物種豐度變化Note:Scale plate color shades indicate changes in species abundance of samples
與WT相比,O組的Skemanella,Subgroup_6,Pseudonocardia,Sphingomonas豐度增加,Actinoplanes和Pseudomonas豐度下降,其余細菌豐度變化不明顯。
3 討論
施用有機肥是提高作物產量及改善土壤理化性質的主要措施,不同類型肥料及施用時間對土壤微生物群落的影響不同。有研究結果顯示,短期施肥對土壤微生物的影響不顯著,長期合理施肥可以豐富土壤微生物多樣性,恢復土壤中固氮菌群的多樣性和群落結構[14,17]。也有研究表明施肥對影響土壤細菌群落豐度、土壤微生物多樣性的影響具有一定的局限性[18-19]。本研究結果表明腐熟牛糞在短期內降低苜蓿根際土壤細菌Alpha多樣性。這與趙鳳艷等研究結果相似,牛糞降低設施番茄根際細菌多樣性和豐富度[20]。Zhang等[21]在茶園施用1.5%牛糞和尿素后,Alpha多樣性指數均低于未施肥,且在取樣時間上土壤細菌的多樣性變化趨勢相似:春季至夏季顯著增加,夏季至秋季顯著減少。也有研究發現細菌豐富度指數Chao和ACE兩個指數在添加固體牛糞和豬糞后下降,在取樣季節之間差異顯著,在不同肥料間差異不顯著[22]。類似研究結果顯示季節對細菌群落的影響大于肥料的營養處理[23]。本研究的多樣性指數和豐富度指數均呈現O> D1> D2> D3,3%的牛糞添加量可使Alpha多樣性顯著下降,土壤細菌的群落結構發生顯著變化。一方面是由于盆栽種植苜蓿,土壤細菌菌群的豐度及多樣性變化會更加顯現;另一方面可能是本試驗施用了相對較高的肥料添加比例導致。
Actinobacteria是所有測試樣品中的優勢菌群,在WT和WF中相對豐度分別達56%和42%,并且隨著牛糞添加比例的增加而呈下降,添加至9%時與O組差異達到顯著。隨著取樣時間的延長也表現下降趨勢。Actinobacteria功能多樣,有助于有機物的分解,研究表明Actinobacteria在未受到干擾的土壤中易富集[24]。Proteobacteria是測試樣品中的另一優勢菌群,在WT中相對豐度最低。Proteobacteria能夠適應營養豐富的環境,Acidobacteria通常被認為喜歡資源有限的寡養環境[25-27]。前人研究發現Proteobacteria與Acidobacteria的比例可以反映土壤營養狀況:二者比例越高意味著更多的有機質投入[28]。本研究也證實了這結論,隨著牛糞添加比例的增加,Proteobacteria豐度增加,Acidobacteria豐度降低,二者比例呈增加趨勢。
Bacteroidetes是一類厭氧菌,菌門成員具有降解復雜有機化合物如纖維素的功能[23]。本研究結果表明土壤中添加牛糞顯著增加Bacteroidetes豐度。Firmicutes也是一類厭氧菌,高碳利用率可能導致其豐度增加。本研究發現隨取樣時間延長,Firmicutes豐度表現增加趨勢,30 d與90 d達到顯著性差異,推測在苜蓿的一個生長周期,碳利用率在持續增加。Gemmatimonadetes可以對土壤中碳和氮含量進行微調,平衡作物代謝需求,適應不斷變化的環境,對土氮素循環中反硝化作用具有重要意義[29]。添加牛糞后Gemmatimonadetes相對豐度顯著下降,但隨著時間延長呈顯著增加,受到作物生長階段影響顯著。Chloroflexi和Patescibacteria的相對豐度隨作物生長呈現顯著增加,這一結果與劉靜平等研究結果相似[30]。但添加牛糞會顯著降低Chloroflexi豐度。Tian等[31]發現其相對豐度與有機肥施用量呈負相關關系。Chloroflexi作為重要的有機物分解者,和放線菌一樣對有機肥施用非常敏感[32],施用有機肥反而會抑制Chloroflexi和Actinobacteria的生長和繁殖,降低二者相對豐度。Nitrospirae是共營養細菌,適合高pH值環境條件生存,施肥降低其豐度[33-34]。本研究結果表明添加牛糞會降低其豐度。
在屬水平上,WT及WF具有不同的優勢菌屬。添加牛糞的土壤可顯著增加有益菌屬芽孢桿菌屬(Bacillus)、馬杜拉放線菌屬(Actinomadura)、BIrii41、德沃斯氏菌屬(Devosia),降低Subgroup_6、Pseudonocardia、芽球菌屬(Blastococcus)、67-14、KD4-96(綠彎菌)的豐度。變形菌門根瘤菌目Devosia豐度在WT及WF樣品中均很低,但在添加牛糞土壤中,該菌屬的豐度顯著增加,達45倍以上,但也隨著苜蓿生長顯著下降。Devosia與水生豆科植物Neptunianatans形成獨特的固氮共生體,有推測Devosia可能有助于速效氮含量的增加[35]。BIrii41顯示與Devosia相同變化,在添加牛糞土壤中,該菌屬的豐度顯著增加,并且隨苜蓿生長時期延長呈增加趨勢。厚壁菌門中與有機化合物分解和植物抗病性相關的Bacillus和對溫度適應的多樣性Planifilum,放線菌門中能降解半纖維素和蛋白質的Saccharomonospora,變形菌門中與植物的抗病性相關的Lysobacter、降解纖維素的Cellvibrio和具有反硝化能力的Steroidobacter[36],這些菌群的豐度在WF,WT,O組差異不顯著,在土壤中添加牛糞后,顯著高于WF,WT,O組,并隨著牛糞含量升高而增加。雖然添加牛糞降低Sphingomonas的豐度,但該菌屬的豐度變化并沒有隨添加比例呈現規律性變化,反而隨著取樣時間延伸呈現出顯著性的降低。
4 結論
添加腐熟牛糞降低盆栽苜蓿土壤細菌Alpha多樣性。Actinobacteria和Proteobacteria是土壤、腐熟牛糞及混合土壤樣品的優勢菌。添加腐熟牛糞增加土壤Firmicutes,Bacteroidetes和Proteobacteria豐度;顯著提高Lysobacter,Bacillus,Saccharomonospora,Planifilum,Cellvibrio,Steroidobacter,Devosia的豐度。不同添加比例均降低土壤細菌OTU數量,但添加比例至6%牛糞的土壤細菌OTU數目隨著苜蓿生長持續增加,高添加比例(添加9%)的細菌OTU數目持續降低。通過不同添加比例的腐熟牛糞對苜蓿土壤細菌結構及多樣性的影響分析,推薦腐熟牛糞添加量低于6%的施肥方式為佳。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
