基于加權基因共表達網絡分析探討黃芩抗肝細胞癌的相關分子機制*
2022-03-28 02:31:10徐玉平蔣向輝
世界科學技術-中醫藥現代化 2022年12期
徐玉平,譚 榮,蔣向輝
(凱里學院大健康學院 凱里 556011)
肝細胞癌(HCC)是一種死亡率很高的原發性肝癌,發病率居世界第六位,死亡率居世界第四位[1],其在40歲前發生率較少,而后隨著年齡逐漸升高,70歲達到頂峰,男性發病率是女性的2-4倍。酒精性肝硬化、肝炎病毒感染、吸煙等是其重要的致病原因。HCC早期不易察覺,病情發展至中晚期后,常出現食欲減退、乏力、腹脹、肝區疼痛等癥狀,還可兼發上消化道出血、肝昏迷、肝癌破裂出血、肝腎衰竭等嚴重并發癥。HCC惡性程度高、易轉移、預后轉歸較差,因此,有“癌中之王”之稱[2]。目前,肝細胞癌的治療手段有手術切除、局部消融、肝臟移植、放化療、介入栓塞術等,但存在供體有限、費用高昂、毒副作用多、易產生耐藥性及腫瘤異質性強[3-5]等問題。近年來很多研究表明中藥可通過多通路抗肝癌組織血管生成,抑制癌細胞增殖和轉移,對肝細胞癌具有一定治療作用[6],而且還能增強機體抵抗力、改善臨床癥狀、緩解放化療帶來的不良反應,提高患者生活質量。
黃芩(Scutellaria baicalensisGeorgi),性寒,味苦,具有清熱燥濕、瀉火解毒、止血、安胎等功效,主治濕溫、暑濕,濕熱痞滿,瀉痢,黃疸,癰腫瘡毒,胎動不安。研究發現,黃芩具有保肝、利膽、抗炎、抗菌、增強免疫、抗腫瘤、神經系統保護和心腦血管保護等多種藥理活性[7],在榮震主任醫師治療HCC的學術經驗整理研究中[8],無論是復方還是單味藥治療使用頻次最高的前10味中藥都包括黃芩。文獻報道,黃芩湯輔助序貫肝動脈化療栓塞術對原發性肝癌患者的治療總有效率為67.86%,顯著高于不加黃芩湯對照組的48.21%,治療后患者的NF-κB、HIF-1α水平、AFP、腫瘤體積顯著低于對照組(P<0.01),抑制了肝癌細胞的生長和擴散[9]。然而關于黃芩治療HCC的臨床研究和相關分子機制卻少見報道。因此,從基因水平深入探討黃芩抗肝細胞癌的核心靶點及其相關分子機制,對HCC的防治具有重要意義,也為黃芩及其有效成分在防治肝癌方面的深入研究、開發和臨床應用提供參考。
加權基因共表達網絡分析(WGCNA)是一種系統生物學方法,通過引入加權的相關系數,構建共表達的基因模塊,能從高通量數據中挖掘出模塊信息,描述不同樣品之間的基因關聯模式,根據模塊的內聯性和模塊與表型之間的關聯性鑒定候補生物標記物或治療標靶。曾祎凡等[10]利用WGCNA技術挖掘促進乳腺癌循環腫瘤細胞轉移的關鍵基因,發現DDX5和ACTB在循環腫瘤細胞內高表達,其中DDX5很可能是促進乳腺癌循環腫瘤細胞轉移的關鍵基因,在臨床上,DDX5可作為乳腺癌轉移早期鑒別的循環腫瘤細胞標志物,也能為改善乳腺癌預后提供新的治療靶點。由此可見,WGCNA技術廣泛應用于基因靶點或疾病生物標志物的篩選之中,適用于中藥生物活性成分結合高通量數據聯合闡述疾病潛在的作用基因及多成分、多靶點、多通路的分子機制。
本研究基于網絡藥理學的方法篩選黃芩的有效活性成分及基因靶點,通過WGCNA分析TCGA和GEO中的HCC相關數據,挖掘HCC的hub基因,并對這些基因進行總生存期、無病生存期分析以及免疫組化驗證,同時聯合GO生物學功能富集分析、KEGG信號通路富集分析進一步探索黃芩抗HCC的相關分子機制,為后期開展黃芩防治HCC的實驗和臨床研究提供理論參考。
1 材料與方法
1.1 黃芩活性成分及其靶點的篩選與PPI網絡構建
中藥系統藥理學(TCMSP)數據庫收納了在中國藥典中注冊的499種中草藥,包含29 384種成分,3311種靶標和837種相關疾病,提供十二種與ADME(Absorption、Distribution、Metabolism、Excretion)相關的重要特性,如口服生物利用度、類藥性、半衰期、Caco-2滲透性、血腦屏障等,用于藥物篩選和評估[11],其中口服生物利用度(OB)、腸上皮細胞通透性(Caco-2)和類藥性(DL)等被認為是中草藥有效性的關鍵指標[12]。本研究通過TCMSP數據庫(https://tcmspw.com/tcmsp.php)檢索黃芩的全部活性成分及其作用靶點,以口服生物利用度(OB)≥40%、類藥性(DL)≥0.18為篩選條件[11],得到黃芩主要的有效活性成分及其對應的作用靶點。再通過檢索UniProt數據庫(https://www.uniprot.org/)獲得去重復項后的靶點蛋白的基因symbol,將靶點上傳到STRING數據庫(https://string-db.org/),設置“highest confidence(≥0.900)”,獲得靶點蛋白的相互作用信息及其相互作用(Protein-protein interaction,PPI)網絡圖,接著導入Cytoscape3.6.1,利用cytoHubba插件計算MCC(中心性),篩選連接程度排名前15的基因為黃芩抗HCC候選基因靶點。
1.2 HCC數據下載
從TCGA數據庫(https://portal.gdc.cancer.gov/)下載與HCC相關的424個轉錄組數據,其中包括正常樣本50個,腫瘤樣本374個,相關參數設置為:數據類別勾選轉錄組數據、表達類別勾選基因表達量、Workflow類別勾選HTSeq-Counts、組織類別勾選肝和肝內膽管、數據庫勾選TCGA,數據集勾選樣本量最多的一個數據庫(即TCGA-LIHC)。
從TCGA數據庫下載與HCC相關的377個臨床數據,其中包括病人的生存時間、生存狀態、性別、年齡、肝癌分期等數據,相關參數設置為:數據類別勾選臨床數據、數據格式勾選bcr xml,組織類別、數據庫、數據集與下載轉錄組數據時的參數一致。
從NCBI的GEO數據庫下載GSE41804數據集的探針矩陣文件,該數據集含有20個HCC腫瘤樣品和20個非腫瘤樣品,通過基因芯片平臺GPL570 [HGU133_Plus_2] Affymetrix Human Genome U133 Plus 2.0 Array下載數據集GSE41804的相關注釋信息文件。
1.3 HCC差異基因篩選
利用R語言(4.0.3版)對TCGA轉錄組數據進行整理,使用edgeR數據包篩選差異表達基因,以log|FC|>1、FDR<0.05為篩選條件,通過limma數據包獲得差異基因(DEGs),再使用ggplot2、pheatmap數據包得到差異基因的聚類熱圖和火山圖,并輸出差異基因的RPKM值用于后續分析。
GEO數據同樣利用R語言(4.0.3版)進行處理,通過使用limma、ggplot2、pheatmap數據包得到差異基因及其聚類熱圖和火山圖。
1.4 WGCNA分析
利用R語言(4.0.3版)的WGCNA包構建HCC加權基因共表達網絡,對基因和模塊、基因與臨床性狀進行相關性分析。先通過樣本聚類樹算法過濾掉離群樣本,以保證共表達網絡構建的穩定性,同時使共表達網絡中的基因之間的連接服從無尺度網絡分布,然后利用Pick Soft Threshold函數選擇最合適的軟閾值β。隨后,將鄰接矩陣轉換為拓撲重疊矩陣(TOM)以降低虛假相關性和噪聲,計算出1-TOM作為共表達網絡互連的重要生物學指標和基因聚類的距離指標,再采用動態樹切割算法識別基因模塊,每個模塊至少包含50個基因。最后,計算基因模塊的特征值(ME),對模塊進行層次化聚類,以0.25為剪切高度合并相似模塊,并用不同顏色加以標記,根據模塊特征關系(ME與臨床特征之間成對的皮爾遜相關性,相關系數>0.80)確定具體的模塊。繪制出聚類模塊特征熱圖,并計算MM值(基因顯著性與模塊顯著性的相關系數)和GS值(基因與臨床特征的相關系數)。基因顯著性(GS)定義為基于臨床特征與基因表達之間的線性回歸P值(GS=logP)的log10變換,模塊顯著性(MS)是指模塊中所有基因的GS的平均值。
1.5 HCC交集基因篩選
一般認為hub基因需要滿足以下條件:GS>0.2,MM>0.8,P≤0.05[13]。MS與GS的絕對值越大,說明與疾病越相關。根據WGCNA分析所獲得的聚類模塊特征熱圖,選擇與正常樣本、腫瘤樣本正相關性最高的兩個基因模塊進行數據合并,對TCGA、GEO數據進行相同處理后,利用R語言(4.0.3版)的VennDiagram數據包對上述合并后的兩個模塊基因與TCGA、GEO的差異基因取交集,輸出文本文件并繪制Venn圖,得到最終的HCC相關基因,并將得到的交集基因與黃芩的15個候選基因靶點進行比對,獲得黃芩抗HCC的潛在基因靶點。
1.6 GO與KEGG富集分析
利用R語言(4.0.3版)的org.Hs.eg.db數據包對上述交集基因進行ID轉換,轉換后獲得相應的entrezID,再利用 R 語言(4.0.3版)的 clusterProfiler、enrichplot、ggplot2等數據包對交集基因進行GO、KEGG富集分析。
1.7 hub基因識別
將WGCNA構建的hub基因模塊中的全部基因取交集后,導入STRING數據庫,基因間相關性打分設置為0.900,從而獲得交集基因網絡關系文件,通過Cytoscape3.6.1對網絡文件進行可視化分析,下載插件CytoHubba計算每個節點的最大團中心性分數(MCC值)并排序,將得分排名前10的基因作為hub基因,并將hub基因與黃芩抗HCC的潛在基因靶點整合,用于下一步分析。
1.8 OS與DFS分析
利用GEPIA數據庫對hub基因進行OS、DFS分析,根據HCC患者hub基因表達量的中位數進行分組,將HCC患者分為低表達組和高表達組,置信區間設置為95%并計算P值,癌癥類型選擇LIHC(肝細胞癌),繪制生存曲線用于可視化分析,篩選與HCC相關的核心基因。
1.9 免疫組化驗證
進入HPA數據庫(https://www.proteinatlas.org/),分別在搜索攔中輸入上述篩選到的核心基因名點擊搜索,選擇“pathology”進入病理界面,看該基因在腫瘤樣品中的免疫組化圖譜,找到“protein expression”,找到肝癌,點擊進入肝癌頁面,向下拉可以找到免疫組化的圖片。抗體HPA代碼必須與正常樣品保持一致。如若要看基因在正常樣品中的免疫組化圖片,可以在搜索欄輸入基因名后,點擊“Tissue”進入組織頁面,余下操作同腫瘤樣品。收集免疫組化圖譜信息,根據蛋白質在組織細胞中的染色程度以及染色細胞百分比,比較正常組織和HCC組織中蛋白表達的差異,并下載保存相應的免疫組化圖片。
2 結果
2.1 黃芩活性成分及靶點與PPI網絡
通過TCMSP數據庫共檢索到黃芩化學成分143種,以OB≥40%、DL≥0.18為篩選條件,得到19種主要活性成分,分別為黃芩新素、欖核蓮黃酮、甲氧基黃酮、黃芩黃酮II、甲氧基查爾酮、二氫木蝴蝶素、苯基苯并吡喃酮、鼠尾草素、表兒茶素、二甲氧基黃酮、蘇薺苧黃酮、豆甾醇、二羧酸苯酯、鄰苯二甲酸二異辛酯、表小檗堿、千層紙素、黃烷酮、紅花素、二氫黃芩苷,見表1,其中黃芩新素的OB值、DL值最高,可能是黃芩抗HCC最主要的生物活性成分。

表1 黃芩主要活性成分及其OB、DL值表
19種主要活性成分的靶點去重復項后,導入STRING數據庫構建PPI網絡,見圖1。將靶點蛋白的相互作用信息文件導入Cytoscape3.6.1,利用cytoHubba插件提取中心子網絡,以MCC進行排序獲得的前15個候選基因靶點依次為 NCOA1、NCOA2、ESR1、AR、RXRA、PPARG、PGR、ESR2、MAPK14、ADRA1B、ADRB2、CYP2C9、PTGS2、ADRA1A、CHRM3,見圖2。
2.2 HCC差異基因
從TCGA數據庫下載與HCC相關的424個轉錄組數據樣本,共包括13913個基因,篩選后獲得2703個差異基因,其中下調基因1680個(左側),上調基因1023個(右側),火山圖見圖3A。
從GEO數據庫下載的與HCC相關的20個腫瘤樣品和20個非腫瘤樣品,共包括21 655個基因,篩選后獲得1050個差異基因,其中下調基因621個(左側),上調基因429個(右側),火山圖見圖3B。

圖1 黃芩主要活性成分靶點的蛋白-蛋白相互作用網絡圖
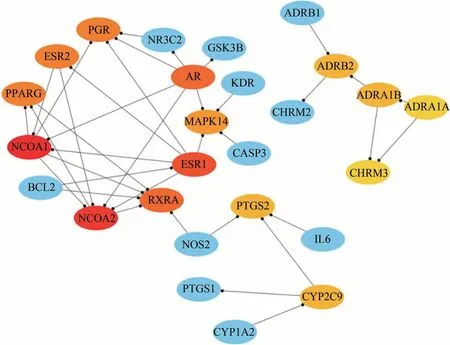
圖2 以MCC進行排序獲得的前15個候選基因靶點圖(黃芩)

圖3 HCC差異基因火山圖
2.3 WGCNA分析
本研究中,TCGA數據選擇軟閾值β=3(無尺度R2=0.9),動態樹切割共識別出9個基因模塊,見圖4A;GEO數據則選擇軟閾值β=2(無尺度R2=0.9),動態樹切割共識別出11個基因模塊,見圖4B。基因模塊與臨床性狀之間的相關性如下圖所示,可知TCGA數據中與正常樣本、腫瘤樣本正相關性最高的模塊分別是黃綠色模塊(MEgreenyellow,R=0.64,P=2e-50)、藍色模塊(MEblue,R=0.69,P=8e-62),見圖4C;GEO數據中與正常樣本、腫瘤樣本正相關性最高的模塊分別是藍色模塊(MEblue,R=0.44,P=0.005)、青綠色模塊(MEturquoise,R=0.75,P=2e-08),見圖4D。因此,我們選擇這四個模塊中的差異基因進行下一步分析。
2.4 HCC交集基因
如圖5所示,TCGA數據中黃綠色模塊和藍色模塊合并后共4701個基因,GEO數據中藍色模塊和青綠色模塊合并后共5928個基因,這些基因與TCGA數據中的2703個差異基因、GEO數據中的1050個差異基因取交集,共得到115個基因,而后將這115個交集基因與黃芩的15個候選基因靶點進行一一比對,獲得黃芩抗HCC的潛在基因靶點ADRA1A。

圖4 WGCNA聚類樹狀圖及聚類模塊特征熱圖
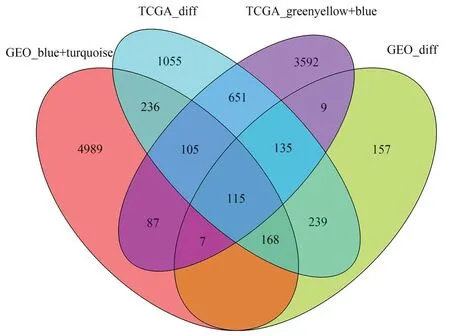
圖5 交集基因與差異基因Venn圖
2.5 GO與KEGG富集分析

圖6 基因富集分析圖
GO分析結果如圖6A所示,上述115個交集基因富集在體液免疫反應、細胞粘附、補體激活、呼吸系統發育、細胞過渡金屬離子穩態、BMP信號通路的調節、反射等生物學過程(BP);KEGG分析結果如圖6B所示,上述115個交集基因富集在膽固醇代謝、礦物質吸收、基底細胞癌、泛醌和其他萜類醌的生物合成、視黃醇代謝、細胞粘附、吞噬體等通路。如圖7所示,交集基因在HCC中主要涉及Wnt信號通路,即Wnt與細胞膜上的受體結合,激活第二信使Dvl,從而抑制由GSK-3β、Axin、APC組成的三元復合物的活性,使β-catenin不能被GSK-3β正常磷酸化,造成β-catenin在胞漿內大量積聚并移向細胞核內,進而與細胞核內的轉錄因子TCF/LEF結合,激活TCF轉錄活性,調節靶基因的表達。
2.6 hub基因識別
通過Cytoscape3.6.1的cytoHubba插件提取上述115個交集基因的中心子網絡,使用CytoHubba插件中的MCC算法計算網絡中每個節點的最大團中心性分數(MCC值),按MCC值的大小進行排序,MCC得分排名前10的關鍵基因依次為FCN2、COLEC11、CFP、COLEC10、FCN3、C7、LCN2、HAMP、B3GAT1、LYVE1。將這10個基因與黃芩抗HCC的潛在基因靶點ADRA1A整合,用于下一步分析。
2.7 OS與DFS分析
根據總生存期曲線和無病生存期曲線,觀察整合后的11個基因在總生存期與無病生存期的基因表達差異是否具有統計學意義(P<0.05),從而進一步篩選出與HCC密切相關的核心基因。OS分析結果顯示,具有統計學意義的基因有FCN3(P=0.033)、ADRA1A(P=0.012)、C7(P=0.03)、COLEC10(P=0.015),且這些基因高表達的患者前80個月生存率更高;DFS分析結果顯示,具有統計學意義的基因有CFP(P=0.029)、FCN3(P=0.0086),且這些基因高表達的患者同樣生存率更高,見圖8。因此,這5個基因可能是黃芩治療HCC的核心基因,分別為纖維膠凝蛋白3(FCN3)、腎上腺素能α1A受體(ADRA1A)、補體C7(C7)、凝集蛋白家族 10(COLEC10)、補體因子備解素(CFP)。其中,纖維膠凝蛋白3在OS、DFS分析中均具有統計學意義,可能是與HCC關系最密切的基因。

圖7 交集基因生成的HCC相關信號通路

圖8 HCC的5個密切相關基因OS、DFS分析

圖9 肝正常組織與HCC組織的免疫組化圖
2.8 免疫組化驗證
通過HPA數據庫分別檢索COLEC10、CFP、ADRA1A,免疫組化結果為空;FCN3、C7免疫組化結果如圖9所示,抗體CAB025945在正常肝組織中FCN3呈現中等強度染色,在HCC組織中則呈現低強度染色,染色細胞百分比為25%-75%;抗體HPA001465在正常肝組織中C7呈現高強度染色,在HCC組織中則呈現中等強度染色,染色細胞百分比均超過75%。這表明與正常肝組織相比,HCC組織中FCN3、C7基因表達量降低,該結果與OS、DFS分析所得結果一致。
3 結論與討論
本研究基于網絡藥理學的方法,發現黃芩可能是通過黃芩新素、欖核蓮黃酮、二氫黃芩苷等活性成分,降低ADRA1A甲基化頻率從而干預HCC的發生;通過對TCGA、GEO數據庫中HCC相關數據進行差異基因篩選、WGCNA分析、GO與KEGG富集分析,并經過OS、DFS、HPA 分析驗證,發現 FCN3、C7、COLEC10、CFP可能是HCC的核心基因,其作用機制可能涉及體液免疫、補體激活、吞噬作用調節和細胞凋亡等過程。
黃芩化學成分眾多,目前為止已被發現的約有40多種,包括黃酮類化合物、萜類、氨基酸、甾醇、揮發油和其他成分,其中黃酮類化合物是黃芩主要的有效成分和特征性成分。研究表明,黃酮類化合物中發揮抗肝癌作用的主要是黃芩素、黃芩苷、漢黃芩素和漢黃芩苷4種活性物質。杜忠良等[14]發現黃芩素能抑制肝癌移植瘤的生長、誘導腫瘤細胞凋亡,其作用機制可能與調控PI3K-AKT通路有關。魯興梅等[15]認為黃芩苷可抑制肝癌荷瘤小鼠瘤組織的增長,其作用機制可能與下調瘤組織PI3K、AKT和m TOR基因及蛋白表達有關。王曉東[16]證明漢黃芩素能通過線粒體信號通路促進人肝癌SMMC-7721細胞的凋亡、抑制人肝癌SMMC-7721細胞增殖。本研究綜合網絡藥理學、癌癥基因組圖譜和基因芯片技術,得到黃芩新素、欖核蓮黃酮、二氫木蝴蝶素、二氫黃芩苷等19種黃芩主要活性成分,其中黃芩新素的口服利用度高達104.34%,類藥性也達到了0.44,具有很高的研究價值。而且目前國內外對黃芩有效活性成分的研究多集中在黃芩苷、漢黃芩苷、黃芩素、漢黃芩素等含量較高的黃酮類成分,對于黃芩新素的研究非常少,而且多集中在提取及制備工藝方面。近年來研究發現,黃芩新素是誘導白血病細胞HL-60和K562凋亡的主要活性成分,其通過上調PP2A表達,下調miR-21表達,使PI3K/AKT/mTOR信號通路失活,從而降低肺癌發生率[17]。Chen等[18]研究發現,雖然黃芩新素在黃芩中的含量較低,但它能特異性結合在免疫抑制和腫瘤生長中起重要作用的靶蛋白STAT3。因此,我們猜測黃芩新素是否也能作為一個重要的活性物質發揮抗肝癌作用呢?這也將為黃芩有效成分及藥理作用的進一步挖掘和HCC的防治提供一個新思路。
GO功能富集分析和KEGG通路富集分析結果顯示,黃芩主要涉及與肝癌相關的信號通路是Wnt信號通路,這與文獻[19]報道一致,Wnt/β-catenin信號通路參與肝臟疾病進展的所有階段,該信號通路的異常將導致肝細胞癌、肝母細胞瘤和膽管癌等多種肝癌的發生,Wnt/β-catenin信號通路受到多種因子的調控,當細胞質中β-catenin不能正常降解時,Wnt/β-catenin信號通路活化增強,多種腫瘤疾病由此引發。目前已經有多項研究報道了調控該信號通路可以抑制肝癌的潛在價值[20-21],因此我們有理由相信黃芩的活性物質黃芩新素可能通過調控Wnt/β-catenin信號通路上下游相關基因的表達,作為防治肝細胞癌潛在藥物的出現是極有可能的。
本研究發現黃芩抗HCC的潛在基因靶點ADRA1A,篩選出的HCC核心基因為FCN3、C7、COLEC10、CFP。腎上腺素能α1A受體(ADRA1A)是一類通過結合兒茶酚胺刺激交感神經系統(SNS)的G蛋白偶聯受體,Chen等[22]的研究表明,與癌旁組織相比,在HCC組織中,ADRA1A mRNA和蛋白的表達水平顯著降低,其啟動子區的平均甲基化水平顯著增加(17.0% vs.25.2%,P<0.0001),ADRA1A基因高甲基化可能導致HCC的發生,這與本研究中ADRA1A的OS分析結果一致(ADRA1A高表達的患者前80個月生存率更高),因此,ADRA1A是黃芩治療HCC的有前景的基因靶點。纖維膠凝蛋白3(FCN3)是機體天然免疫的關鍵性分子,主要參與補體激活、吞噬作用調節和細胞凋亡過程[23],研究發現,FCN3功能障礙或表達異常可能造成人類某些感染性疾病、慢性疾病及自身免疫性疾病的發生[24],與KEGG富集分析結果一致。補體C7是補體級聯反應的下游,通過補體經典途徑或者凝集素途徑激活,補體途徑還可激活天然免疫系統,在抵抗細菌等病原體感染中起重要作用。張鑫等[25]報道,通過供肝組織SNP檢測分型發現,供肝C7 rs6876739基因型與肝移植術后的早期細菌感染顯著相關,這可能與移植肝中C7合成減少有關;Würzner等[26]報道,移植肝合成高達50%以上的C7,說明C7可作為HCC的預后因子。凝集蛋白家族10(COLEC10)是HCC中miR-452-5p的直接靶標,miR-452-5p過表達能夠顯著加速細胞增殖,誘導細胞周期從G1轉變,抑制癌細胞的凋亡[27-28],說明COLEC10是HCC的核心基因之一。OS與DFS分析、以及免疫組化驗證也已證實,FCN3、C7、COLEC10、CFP的異常表達與HCC的發生發展密切相關。
綜上所述,本研究初步揭示了黃芩抗HCC的潛在基因靶點和HCC核心基因,探討了其相關分子機制,為后期開展黃芩治療HCC的實驗和臨床研究提供理論參考。但本研究仍然存在不足和局限性:
(1)數據來源比較單一
網絡藥理學的重要基礎是對不同來源數據進行整合分析,不同數據庫如常用的中藥成分數據庫(ETCM、TCMSP、TCMID、TCM)數據收錄差異大、且數據信息之間無有效關聯相對比較獨立,因此數據收集的完整性和整合分析對研究結果的可靠性、可重現性有較大影響。本研究黃芩活性成分及其靶點的篩選選用的是TCMSP,來源比較單一,一定程度上可能影響研究結果的重復性。以后我們可以盡量通過整合多個來源的數據、同時結合臨床或實驗方法進行數據采集、檢測和補充[29]。
(2)缺乏更加有效、科學的驗證
本研究遵守2021年世界中醫藥學會聯合會發布的《網絡藥理學評價方法指南》[30-31],并按要求對其內容進行可靠性評價、規范性評價和合理性評價。但在結果驗證方面,本研究目前還停留在文獻驗證和計算機實驗驗證階段,缺乏更加科學有效和更具說服力的驗證方式,如體外細胞模型、動物模型實驗驗證,甚至臨床試驗驗證等方式。
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
電子制作(2018年18期)2018-11-14 01:48:24
財經(2017年2期)2017-03-10 14:35:35
山東工業技術(2016年15期)2016-12-01 05:31:22
財經(2016年15期)2016-06-03 07:38:02
財經(2016年3期)2016-03-07 07:44:46
財經(2016年6期)2016-02-24 07:41:51
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
