江西鉛山紅芽芋TIFY 9基因克隆與功能分析
2022-04-01 15:25:08趙紅鄧雨晴曾淑琴曾熒張彩霞張慧博張麗佳張玲張妮尹明華
江蘇農業科學 2022年5期
關鍵詞:煙草
趙紅 鄧雨晴 曾淑琴 曾熒 張彩霞 張慧博 張麗佳 張玲 張妮 尹明華







摘要:為了深入探究江西鉛山紅芽芋TIFY 9基因的功能,通過江西鉛山紅芽芋試管苗轉錄組數據庫篩選到江西鉛山紅芽芋TIFY 9基因的核心片段,利用RT-PCR 技術克隆該基因,并采用生物信息學方法、轉基因瞬時表達和實時定量 PCR進行序列分析、亞細胞定位、器官表達分析和功能分析。結果表明:江西鉛山紅芽芋TIFY 9基因cDNA總長度為522 bp,G+C 含量為58.62%;由174個氨基酸組成,分子量19 073.87 u,等電點9.30,為親水性蛋白;二級結構由α-螺旋(31.61%)、β-片層(12.64%)、無規則卷曲(55.75%) 構成,三級結構為單體;主要存在細胞核中;在進化上與Colocasia esculenta(芋)的親緣關系較近,與Colocasia esculenta(芋)hypothetical protein Taro_030839(MQL98131.1)在進化上具有最高的親緣關系。江西鉛山紅芽芋TIFY 9蛋白具有典型植物TIFY 9的結構特征,氨基酸序列及核酸序列與同源物種同源性高達100%,在進化上高度保守。煙草葉片亞細胞定位分析表明,TIFY 9定位于細胞質和細胞核中。實時定量PCR結果顯示,TIFY 9基因在江西鉛山紅芽芋中的表達存在器官特異性,在江西鉛山紅芽芋不同組織部位均有表達,且在葉中和球莖膨大初期表達量最高。在煙草花葉病毒脅迫下,與TIFY 9非轉基因煙草相比較,TIFY 9轉基因煙草的凈光合速率(Pn)、氣孔導度(Gs)、蒸騰速率(Tr)、最大光化學效率(Fv/Fm)、實際光化學量子效率(ΦPS Ⅱ)、光化學淬滅系數(qP)及電子傳遞效率(ETR)顯著增加,而非光化學淬滅系數(NPQ)顯著下降,表明TIFY 9基因響應病毒脅迫應答可能是依賴光合作用的提高。
關鍵詞:江西鉛山紅芽芋;TIFY 9基因;基因克隆;功能分析;轉基因表達
中圖分類號: S632.301文獻標志碼: A
文章編號:1002-1302(2022)05-0041-09
收稿日期:2021-04-27
基金項目:國家自然科學基金(編號:32060092、31960079);2021年度江西省教育廳科學技術研究項目(編號:GJJ211729);上饒市科技局平臺載體建設項目(編號:2020I001、2019I017)。
作者簡介:趙 紅(1966—),女,江西上饒人,碩士,高級實驗師,主要從事植物生物技術研究工作。E-mail:928807480@qq.com。
通信作者:尹明華,碩士,教授,主要從事植物生物技術研究工作。E-mail:yinminghua04@163.com。
江西鉛山紅芽芋(Colocasia esculenta L. Schoot var. cormosus‘Hongyayu’)屬于天南星科本草植物,藥食兼優,藥用可寬腸胃、補脾胃、腹中痞塊、消癆散結、調節免疫,食用營養豐富、清淡爽口、粉糯細膩,是老幼皆宜的美味佳肴[1]。2013年4月,江西鉛山紅芽芋核準為國家地理標志農產品,現已成為江西省上饒市鉛山縣“兩紅”(紅芽芋和河紅茶)產業之一[2]。TIFY 蛋白屬于植物特有GATA 轉錄因子家族,由于花序分生組織表達的鋅指蛋白(zinc-finger protein expressed in inflorescence meriste,ZIM)家族具有高度保守的 TIFY 功能結構域,因此TIFY 蛋白也被命名為 TIFY 基因家族。根據保守結構域的組成,TIFY 家族蛋白可分為 4 個亞家族 (TIFY、JAZ、ZML 和 PPD)和2個亞群(亞群Ⅰ含 C2C2-GATA 結構域,亞群Ⅱ不含該結構域)[3]。TIFY 蛋白是控制植物中各種生物途徑的主要關鍵因子,在植物發育過程中參與防御和應激反應,以應對生物和非生物脅迫響應[4-5]。TIFY1是首個在擬南芥中被鑒定的TIFY 家族轉錄因子[6]。目前,TIFY家族已被克隆和鑒定[7-9]。植物病毒有“植物癌癥”之稱,可通過維管束侵染植物,造成生物脅迫逆境,經濟損失巨大,因此培育無病毒植株至關重要[10]。因此,對江西鉛山紅芽芋TIFY 9基因進行克隆和功能分析對培育江西鉛山紅芽芋的脫毒品種具有重要意義。近年來,TIFY 蛋白在植物抗逆脅迫中的研究成為熱點,多集中在植物應對一些非生物脅迫如干旱、鹽堿、滲透等逆境上,而對于生物脅迫相關研究較少。有研究表明,玉米的ZmJAZ基因對某些生物脅迫真菌(如絲黑穗、莖腐病和炭疽病)具有一定程度的脅迫響應[11]。目前,對紅芽芋的研究主要集中在脫毒快繁[2]、微芋誘導[1]、下游加工[12]、成分分析[13]等方面,關于紅芽芋TIFY 9蛋白的克隆及其功能分析尚未見報道。利用RT-PCR 技術克隆江西鉛山紅芽芋TIFY 9基因,并采用生物信息學方法、煙草轉基因技術和實時定量 PCR進行序列、功能和組織表達分析,旨在為揭示江西鉛山紅芽芋TIFY 9的生物學功能提供理論依據,為從分子水平選育江西鉛山紅芽芋脫毒品種提供新思路。
1 材料與方法
1.1 試驗材料
江西鉛山紅芽芋試管苗。試驗于2020年5月至2021年4月在上饒師范學院生命科學學院進行。
1.2 試驗方法
1.2.1 總RNA的提取和cDNA第1鏈的合成 用Trizol 試劑提取江西鉛山紅芽芋試管苗的總 RNA,提取步驟按說明書進行,使用紫外分光光度計和瓊脂糖凝膠電泳檢測 RNA 的濃度和完整性。以提取獲得的 RNA為模版,按照 M-MLV cDNA 第1鏈合成試劑盒說明書合成 cDNA 第1鏈。逆轉錄引物用 Oligo(dT) 18 Primer:5′-GGCCACGCGTCGACTAGTACTTTTTTTTTTTTTTTTTT-3′,具體步驟按說明書進行。
1.2.2 TIFY 9基因的克隆 利用轉錄組組裝的Unigene序列信息(TRINITY_DN3508_c0_g1),運用 Primer Premier 5.0 設計引物(F:GGGCGTGGTCCTCGGATAACA;R:AGACGGTGACGGTGCCGTTGT)。PCR擴增條件:95℃ 2 min;95 ℃ 30 s,51 ℃ 30 s,72 ℃ 30 s,35個循環;72 ℃ 10min。PCR 產物經 1%瓊脂糖凝膠電泳檢測后,將含有目的基因的條帶與 pMD19-T 載體連接并用熱激法轉化到感受態細胞 Escherichia coli DH5α,經鑒定正確的陽性轉化子提取質粒送往生工生物工程(上海)有限公司進行測序。
1.2.3 TIFY 9基因的生物信息學分析 使用 BioEdit軟件翻譯基因序列為氨基酸序列,用 ProtParam預測酶的理化性質,用 ProtScale預測酶的疏/親水性。使用GOR I軟件在線預測酶的二級結構。使用SWISS-MOLD在線預測酶的三級結構。采用 WoLFPsort在線預測基因的表達部位。通過軟件DNAMAN和 Bioedit進行氨基酸序列比對,利用MEGA5.0軟件進行系統進化樹的構建。
1.2.4 基因克隆和載體構建 以提供的目的基因序列設計引物(去掉終止密碼子TAG/TAA/TGA),CZ-T9-BamHⅠ-JF:ATGTCTAGACTCGAGATGTCCAAGTCTTACGTCG;CZ-T9-BamHⅠ-GFPR:GGCCGCTGTACACATGCAGGGAGGAGAGGGAAG,粗體部分為載體同源重組序列,以轉錄組篩選的cDNA為模板,以所述引物進行PCR擴增(采用南京諾唯贊生物科技股份有限公司Phanta Max Super-Fidelity DNA Polymerase)。PCR反應體系(總體積50 μL)包括 17 μL ddH2O、25 μL 2×Phanta Max Buffer、1 μL dNTP Mix(均為10 mmol/L)、2 μL 模板DNA、2 μL引物1(10 μmol/L)、2 μL引物2(10 μmol/L)和 1 μL Phanta Max Super-Fidelity DNA Polymease(1 U/μL)。PCR反應程序為:95 ℃ 30 s;95 ℃ 15 s,51 ℃ 15 s,72 ℃ 1 min,39個循環;72 ℃ 5 min。將PCR產物進行凝膠電泳。將條帶切膠回收,即為目的基因片段。用BamHI酶切載體pCambia2301-KY-GFP線性化,回收后和目的基因片段重組反應(采用賽默飛世爾科技公司或TaKaRa公司相應限制性內切酶產品)。酶切反應體系包括2 μL 10×K Buffer、5 μL 載體質粒、2 μL BamHⅠ和ddH2O(補足 40 μL)。酶切產物純化后與上述PCR產物進行重組反應(重組反應試劑盒為南京諾唯贊生物科技股份有限公司的ClonExpress-Ⅱ One Step Cloning Kit)。重組連接反應體系(總體積10 μL)包括4 μL 線性化載體、1 μL 插入片段、2 μL 5×CE Ⅱ Buffer、1 μL Exnase Ⅱ和ddH2O(補足10 μL)。上述反應液使用移液器輕輕吸打混勻,短暫離心將反應液收集至管底。置于37 ℃反應30 min,隨后立即置于冰上冷卻。重組產物轉化大腸桿菌DH5α細胞。挑取PCR陽性的轉化子搖菌培養提取質粒,同時擴增產物送測序。擴增和測序引物為插入目的基因兩側的載體序列(目的基因較短則只進行一側測序),分別為35S-F:5′-GACGCACAATCCCACTATCC-3′;GFP-JR:5′-GGGTGAGCTTGCCGTAGGTG-3′。
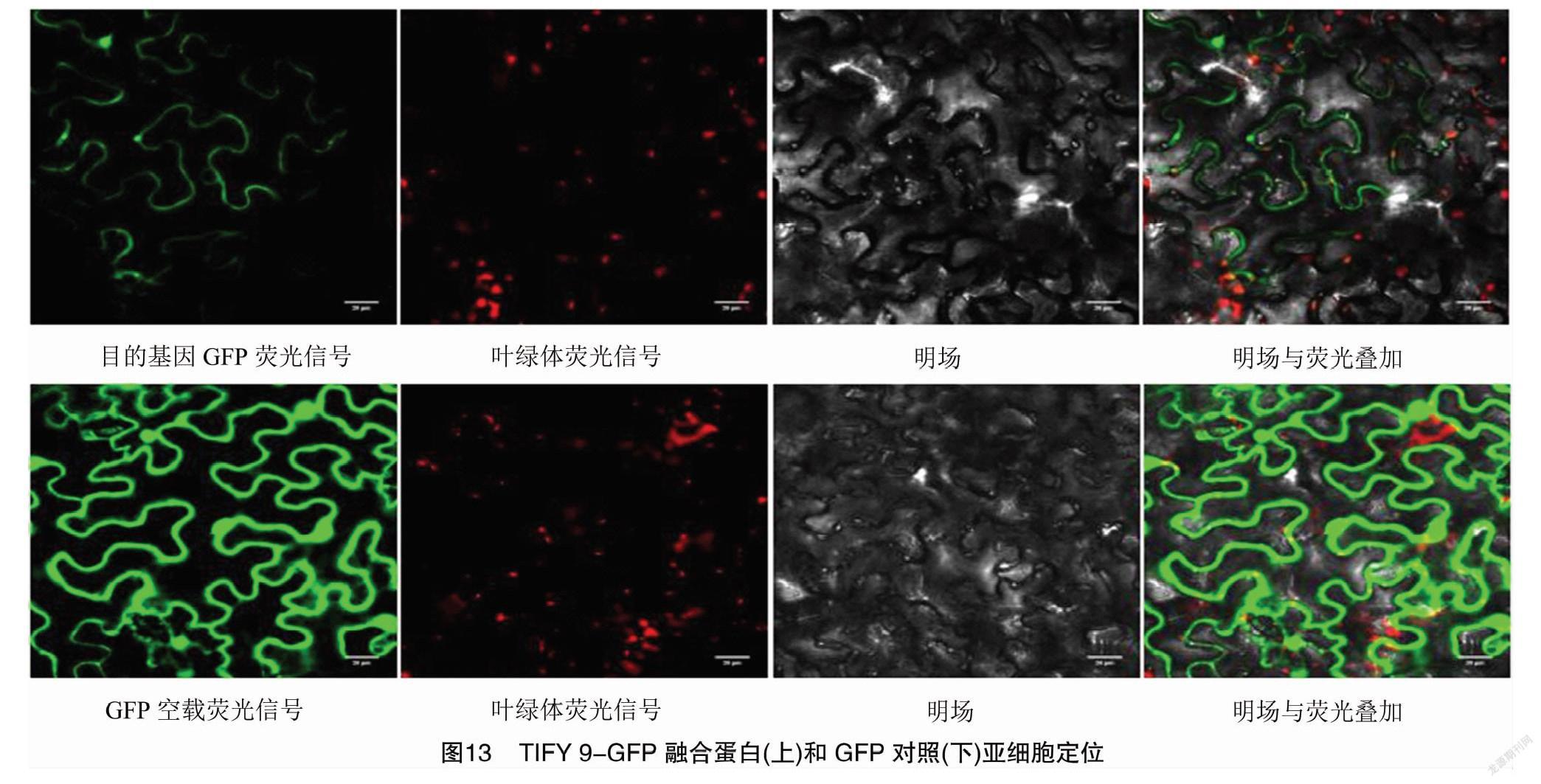
1.2.5 煙草葉片亞細胞定位 播種煙草,培養約1個月用于試驗;將構建好的載體質粒轉入農桿菌GV3101,涂布卡那霉素抗性平板,挑取單克隆于YEB液體培養基在28 ℃搖床內小搖、大搖培養 2 d,菌體4 000 r/min離心4 min,去上清后菌體用用10 mmol/L MgCl2(含120 μmol/L乙酰丁香酮)懸浮液重懸菌體,調整D600 nm值為0.6左右;用無針頭的1 mL注射器吸取農桿菌液,從煙草葉片下表皮(背面)壓迫注射;將注射完成的煙草植株弱光培養2 d,即可觀察;接種2 d后取注射區域的煙草葉片,制作玻片,在激光共聚焦顯微鏡(FluoView FV1Oi,OLYMPUS公司)下觀察、拍照;同時以空載體轉化的農桿菌作為對照重復上述操作。葉綠體熒光信號說明:葉綠體熒光信號激發波長640 nm,發射波長675 nm;綠色熒光蛋白(GFP)信號說明:綠色熒光蛋白GFP激發光488 nm,發射光510 nm。
1.2.6 煙草TIFY 9轉基因PCR檢測和GUS組織化學染色 煙草葉片用農桿菌侵染后均經過4次篩選/繼代,誘導出抗性芽,再轉入伸長培養基中。采用PCR對其再生苗進行鑒定,得到TIFY 9基因的煙草轉基因株系。PCR檢測引物:載體上游引物35S-F:GACGCACAATCCCACTATCC;目的基因下游引物:GCAGGGAGGAGAGGGAAG。使用南京諾唯贊公司Taq 酶擴增體系和條件。2×Taq Master Mix 擴增體系(共20 μL)包括7μL ddH2O、10 μL 2×Taq Master Mix、1 μL 模板DNA、1 μL引物1(10 μmol/L)、1 μL 引物2 (10 μmol/L)。PCR反應程序:95 ℃ 5 min;95 ℃ 30 s,55 ℃ 30 s,72 ℃ 60 s,35個循環;72 ℃ 5 min。GUS組織化學染色:采集煙草TIFY 9轉基因株系和非轉基因株系移栽苗的葉片組織,加GUS染液于37 ℃過夜染色;染色后的葉片再用75%的乙醇脫色(去除葉綠素),也于37 ℃過夜。倒掉脫色乙醇后再用75%乙醇清洗后觀察并拍照。
1.2.7 煙草TIFY 9轉基因株系和非轉基因株系移栽苗的病毒接種和光合指標檢測 培養3個月后,取含煙草花葉病毒的煙草試管苗葉片放在研缽中并加入適量磷酸緩沖液研磨成糊狀病樣汁液;然后在煙草TIFY 9轉基因株系和非轉基因株系移栽苗幼嫩的葉面上灑少許等量金剛砂,蘸取含煙草花葉病毒的煙草試管苗等量病樣汁液在灑了金剛砂的葉面上輕抹 3次(模擬蚜蟲的病毒田間傳播),之后用無菌水清洗葉面;然后放入智能人工氣候室內澆灌MS液體培養基培養;2個月后采用Li-6400便攜式光合測定儀(LI-COR公司,美國)儀測定煙草TIFY 9轉基因株系和非轉基因株系移栽苗的光合參數和葉綠素熒光參數。所有數據表示為平均值±標準差,并使用SPSS 19.0軟件進行統計分析,應用單因素方差分析(One-way ANOVA)檢驗TIFY 9基因組織表達的差異顯著性(α=0.05)。
1.2.8 TIFY 9基因的組織表達分析 分別取江西鉛山紅芽芋試管苗的根、莖、葉、試管球莖(初期、中期和末期)RNA 500 ng,反轉錄為cDNA。熒光定量 PCR(qRT-PCR,SYBR Green Ⅰ)檢測內參基因為GAPDH。設計引物(F:TGCCCTGAGCGTTCCCTAC;R:ACCCGCCTCCACTTCTTCC;長度:178 bp;Tm:583 ℃)。qRT-PCR 檢測采用20 μL反應體系,PCR反應程序:95 ℃ 10 min;95 ℃ 10 s,60 ℃ 34 s,95 ℃ 15 s,40個循環。使用 2-ΔΔCT 法計算基因表達水平。試驗重復3次。所有數據表示為平均值±標準差,并使用SPSS 19.0軟件進行統計分析,應用單因素方差分析(One-way ANOVA)檢驗TIFY 9基因組織表達的差異顯著性(α=0.05)。
2 結果與分析
2.1 cDNA序列
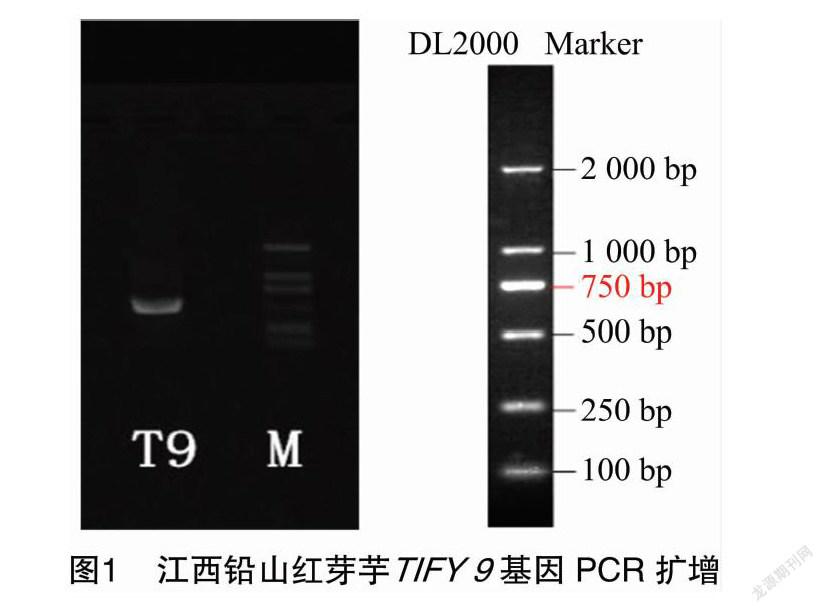
通過PCR擴增技術(圖1),江西鉛山紅芽芋TIFY 9基因cDNA總長度為522 bp(圖2),G+C含量為58.62%。
2.2 氨基酸序列
Protparam預測顯示江西鉛山紅芽芋TIFY 9氨基酸序列見圖3。該基因由174個氨基酸組成,分子量19 073.87 u,等電點9.30,為親水性蛋白。各氨基酸的數量和比例為Ala(A)(18,10.3%)、Arg(R)(13,7.5%)、Asn(N)(7,4.0%)、Asp(D)(8,4.6%)、Cys(C)(3,1.7%)、Gln(Q)(3,1.7%)、Glu(E)(9,5.2%)、Gly(G)(6,3.4%)、His(H)(1,06%)、Ile(I)(8,4.6%)、Leu(L)(13,7.5%)、Lys(K)(9,5.2%)、Met(M)(6,3.4%)、Phe(F)(7,40%)、Pro(P)(17,9.8%)、Ser(S)(21,12.1%)、Thr(T)(10,5.7%)、Trp(W)(2,1.1%)、Tyr(Y)(3,1.7%)、Val(V)(10,5.7%)。帶負電殘基總數(Asp+Glu)為17,正電荷殘基總數(Arg+Lys)為22。估計半衰期為30 h(哺乳動物網織紅細胞,體外);>20 h(酵母,體內);>10 h(大腸桿菌,體內)。不穩定指數:失穩指數(Ⅱ)計算為5469,這將蛋白質分類為不穩定的。
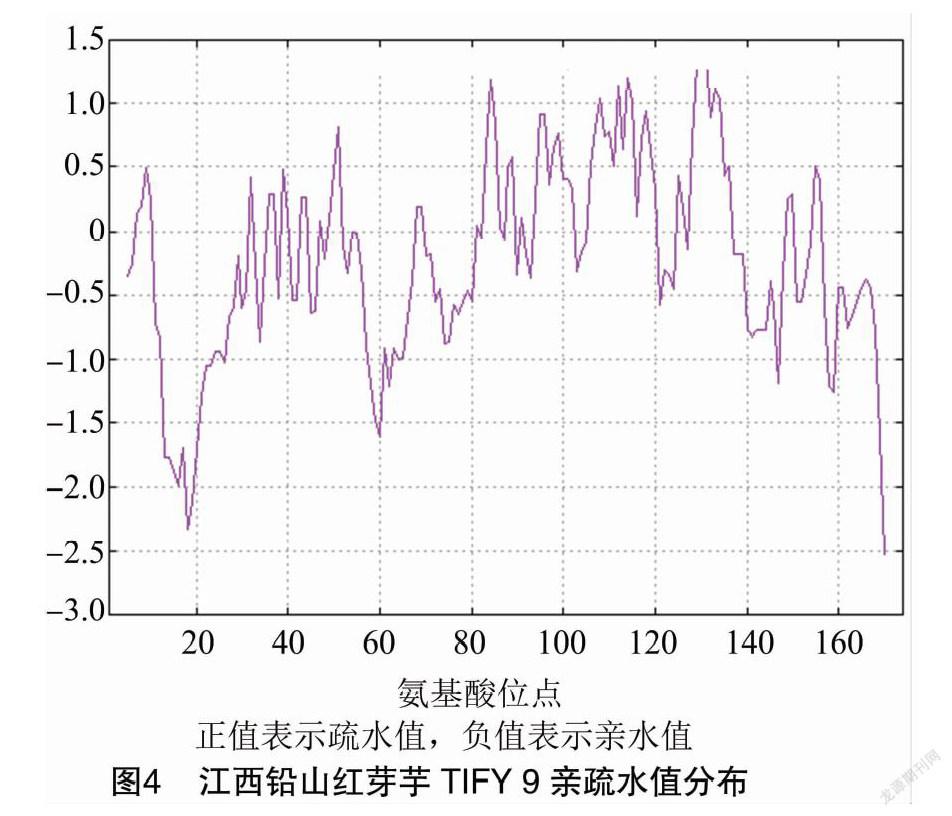
2.3 親疏水性分析
從圖4可知,高峰值(正值)的區域表示疏水的區域,而低谷值(負值)的區域是親水區域。其最大疏水值為1.5左右,說明在該多肽中該處的疏水性最強;親水峰最大值為-2.5左右,整個蛋白質表現出高度的親水性,說明該蛋白為親水性蛋白質。
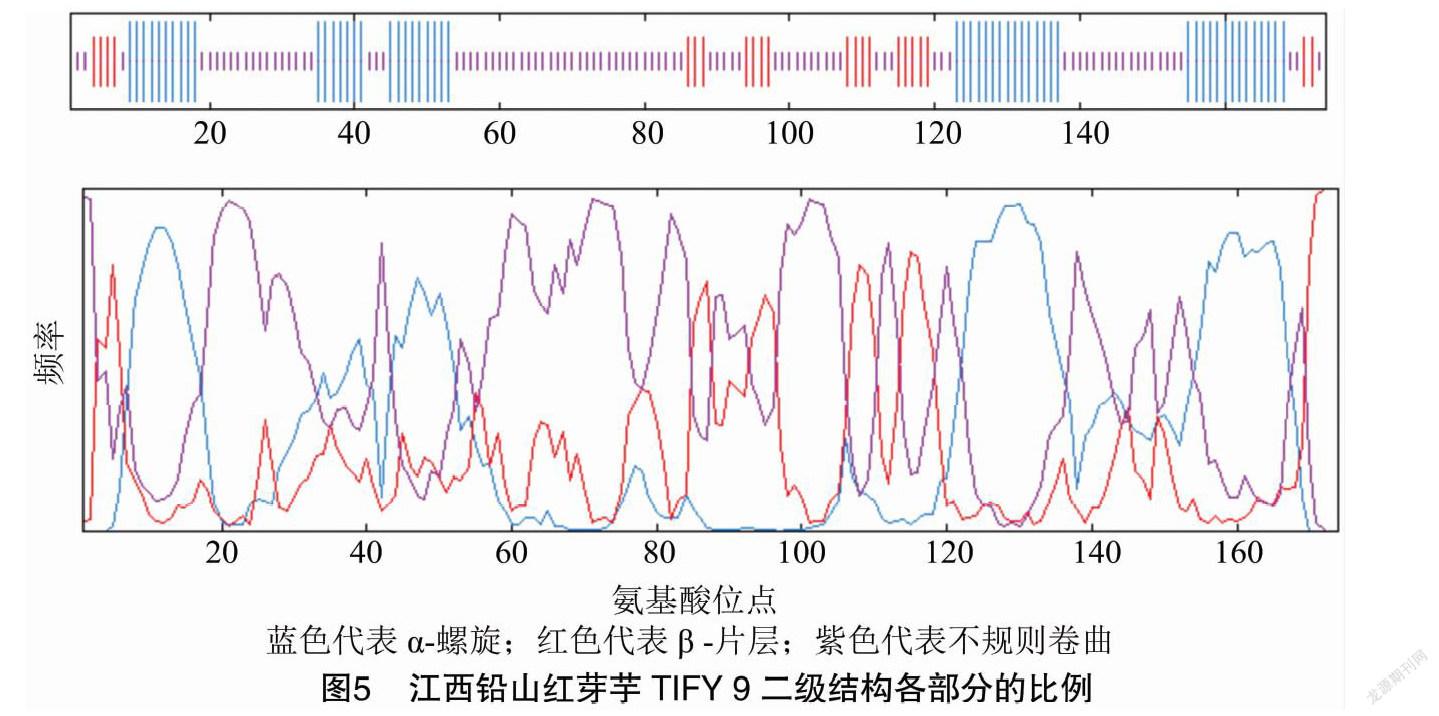
2.4 二級結構分析
GOR 預測顯示江西鉛山紅芽芋TIFY 9二級結構由α螺旋(Hh,31.61%)、β-片層(Ee,12.64%)、 無規則卷曲(Cc,55.75%)構成(圖5)。從分布位點上來看,C端和N端主要含α-螺旋、無規則卷曲和β-片層,且無規則卷曲、β-片層和 α-螺旋則散布于整個蛋白質中。
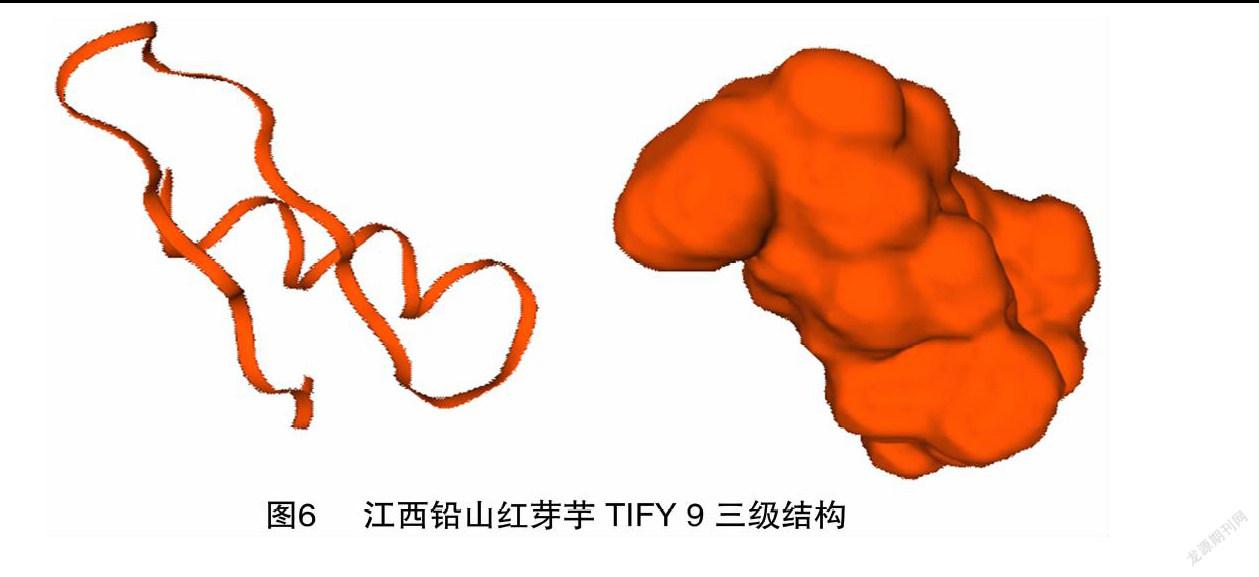
2.5 三級結構分析
SWISS-MODEL預測顯示江西鉛山紅芽芋TIFY 9的三級結構為同源三聚體(圖6)。
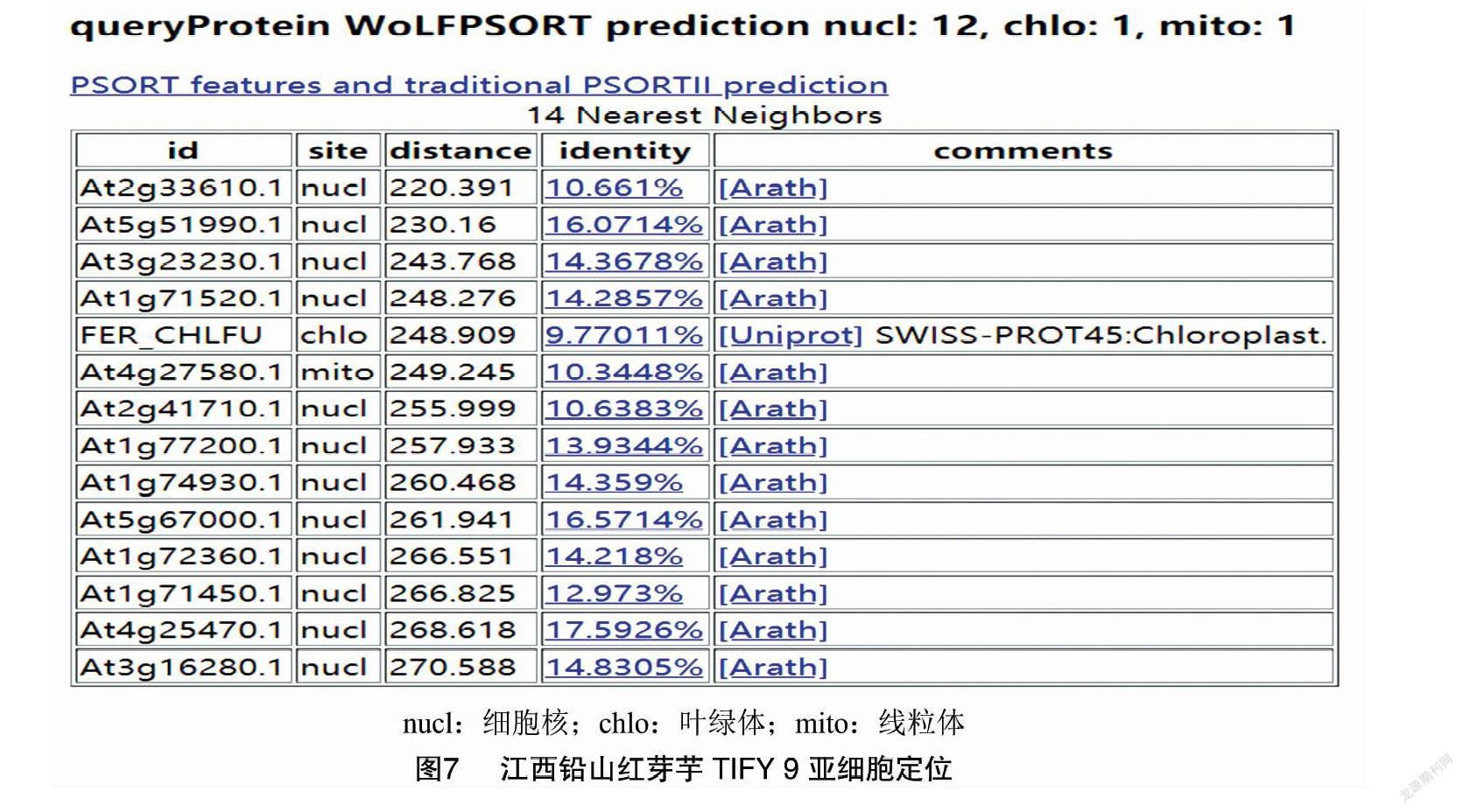
2.6 亞細胞定位
采用 WoLFPsort在線軟件對江西鉛山紅芽芋TIFY 9基因的表達部位進行預測。圖7顯示,定位于細胞核中的數量為12,線粒體中的數量為1,葉綠體中的數量為1。表明江西鉛山紅芽芋TIFY 9基因主要存在細胞核中。
2.7 系統進化分析
從構建的進化樹(圖8)可見,江西鉛山紅芽芋與Colocasiaesculenta(芋)在一個大分支下,這說明江西鉛山紅芽芋TIFY 9在進化上與Colocasia esculenta(芋)的親緣關系較近,與Colocasia esculenta hypothetical peotein Taro_030839(MQL98131.1)在進化上具有最高的親緣關系。
2.8 同源蛋白的序列比對信息
同源蛋白的序列比對信息見圖9,其中※號區域是該蛋白家族的保守結構域。
2.9 目的片段擴增與瞬時表達載體構建
以提供的目的基因序列設計引物(去掉終止密碼子TAG/TAA/TGA)進行PCR擴增,對PCR產物凝膠電泳結果顯示,在目標位置擴增到單一條帶,大小正確。將條帶切膠回收,即為目的基因片段(圖10)。用BamHⅠ酶切載體pRI101-GFP線性化,回收后和目的基因片段重組反應。重組產物轉化大腸桿菌DH5α細胞。挑取PCR陽性的轉化子搖菌培養提取質粒,對重組質粒進行酶切檢測(圖11),同時擴增產物送測序。測序比對結果顯示,目的基因已插入載體,表達載體構建準確(圖12)。
2.10 亞細胞定位分析
通過煙草葉片瞬時表達,將TIFY 9融合綠色熒光蛋白(GFP)的載體質粒TIFY 9-GFP轉入農桿菌,吸取農桿菌液,壓迫注入煙草葉片下表皮(背面)進行煙草病毒增殖蛋白1亞細胞定位分析(圖13)。利用激光共聚焦顯微鏡觀察,融合GFP的TIFY 9轉化煙草葉片只在細胞質和細胞核膜觀察到綠色熒光,表明TIFY 9定位于細胞質和細胞核。
2.10 轉基因煙草鑒定、GUS組織化學染色和光合特征
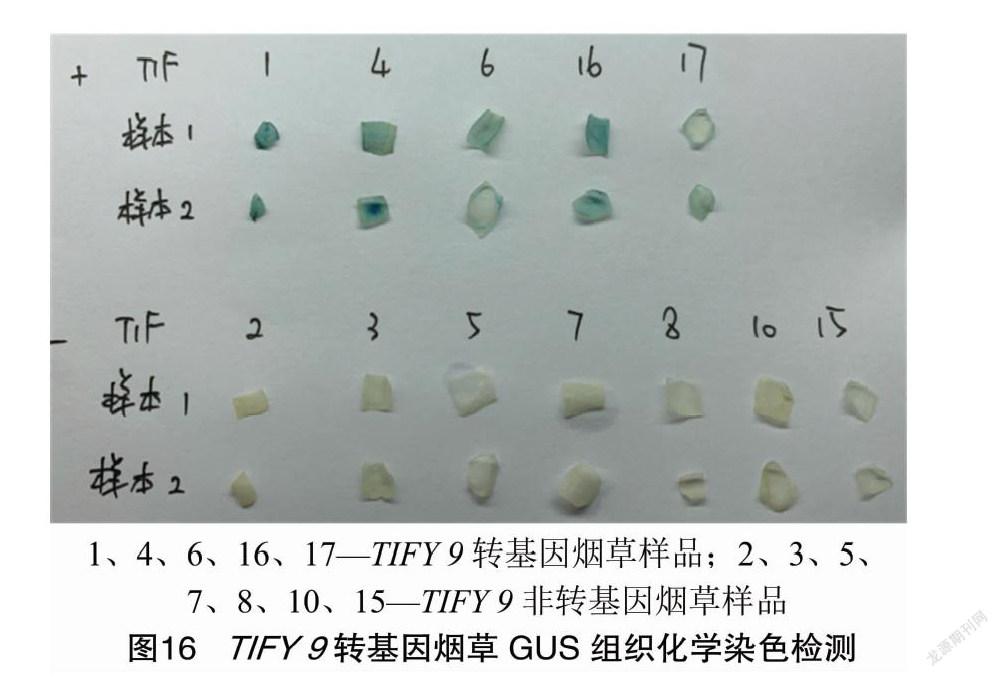
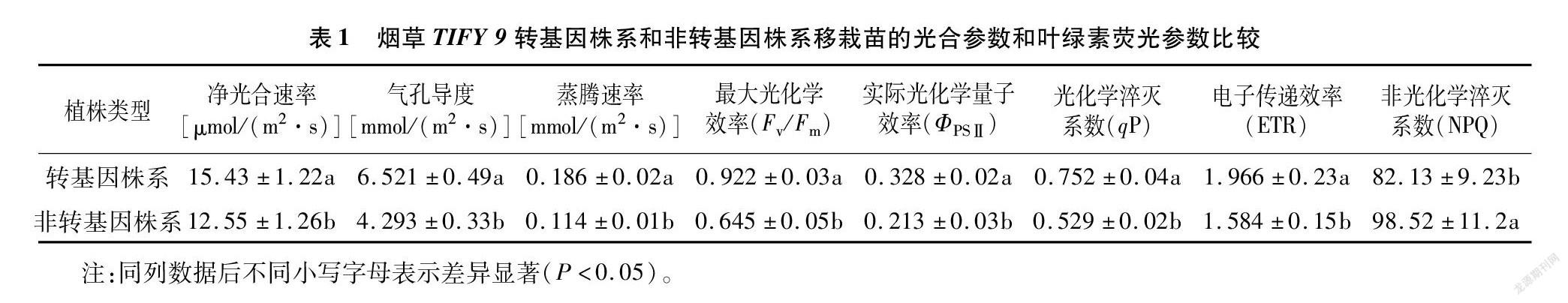
煙草葉片用農桿菌侵染后均經過3次篩選/繼代,誘導出抗性芽,再轉入伸長培養基,最后將其移栽成活(圖14)。經PCR檢測(圖15),獲得TIFY 9轉基因煙草和TIFY 9非轉基因煙草。在PCR檢測基礎上,對2種煙草進行GUS染色驗證(圖16)。結果表明,PCR檢測和GUS檢測結果一致。然后模擬蚜蟲的病毒田間傳播,在煙草TIFY 9轉基因株系和非轉基因株系移栽苗涂抹含煙草花葉病毒的煙草試管苗等量病樣汁液,2個月后測定煙草TIFY 9轉基因株系和非轉基因株系移栽苗的光合參數和葉綠素熒光參數。結果表明,與TIFY 9非轉基因煙草相比較,TIFY 9轉基因煙草的凈光合速率(Pn)、氣孔導度(Gs)、蒸騰速率(Tr)、最大光化學效率(Fv/Fm)、實際光化學量子效率(ΦPSⅡ)、光化學淬滅系數(qP)及電子傳遞效率(ETR)顯著增加,而非光化學淬滅系數(NPQ)顯著下降(表1)。
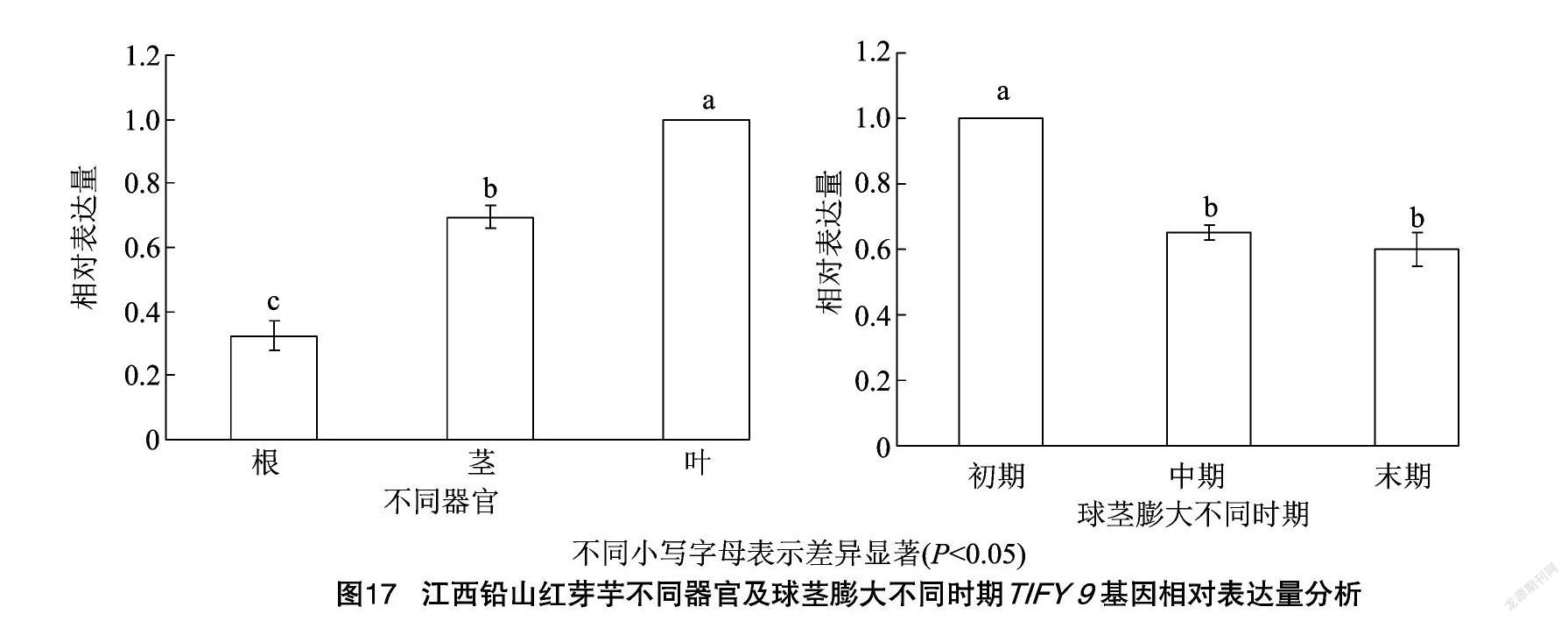
2.11 不同器官及球莖膨大不同時期TIFY 9基因的表達分析
以江西鉛山紅芽芋的GAPDH為內參,利用實時熒光定量PCR分析江西鉛山紅芽芋TIFY 9基因在江西鉛山紅芽芋不同器官中的表達情況。結果顯示,江西鉛山紅芽芋TIFY 9基因在根、莖、葉中均有表達,但在不同組織器官的表達情況差異顯著(圖17),其中江西鉛山紅芽芋TIFY 9基因在葉和球莖膨大初期中表達量最高。
3 結論與討論
TIFY蛋白是植物特有的轉錄因子,在植物的生長發育、響應逆境脅迫和信號轉導等過程中發揮重要作用[14-15]。前期研究中,筆者所在課題組發現1個江西鉛山紅芽芋TIFY9基因在超低溫療法脫毒處理的脫毒苗葉片中的表達量下調,說明它是一個芋花葉病毒(DsMV)、黃瓜花葉病毒(CMV)、芋羽狀斑駁病毒(TFMoV)等病毒侵染后誘導的基因[1]。在本試驗中,亞細胞定位預測結果顯示江西鉛山紅芽芋TIFY 9主要定位在細胞核、線粒體和葉綠體中,與煙草葉片TIFY 9轉基因瞬時過表達的亞細胞定位分析結果一致,也與轉錄因子核定位特征一致[16]。有研究表明,龍血樹DcTIFY基因在根和莖中的表達量最高[17],而在本試驗中,江西鉛山紅芽芋TIFY 9基因在葉和球莖膨大初期中表達量最高。江西鉛山紅芽芋TIFY 9基因在葉中的表達水平顯著高于其他部位(根和莖),且在球莖膨大初期的表達水平
顯著高于其他時期(中期和晚期),說明它可能在葉片中和球莖膨大初期發揮著重要的作用,推測江西鉛山紅芽芋TIFY 9基因可能參與了調控球莖膨大成熟過程。在本試驗中,江西鉛山紅芽芋TIFY9 轉錄因子與Colocasia esculenta(芋)hypothetical peotein Taro_030839(MQL98131.1)在進化上具有最高的親緣關系(100%),說明江西鉛山紅芽芋TIFY 9相對保守。有研究表明,TIFY基因在病原體反應中起著重要作用[8,18]。在本試驗中,江西鉛山紅芽芋TIFY 9蛋白編碼174個氨基酸,為親水性不穩定蛋白,因此分析判斷江西鉛山紅芽芋TIFY 9基因可能與病毒脅迫逆境相關。經禾甲安處理后的番木瓜感病植株中CpTIFY10A-like基因表達量顯著升高,而番木瓜環斑病毒(PRSV)基因表達量顯著降低,推測禾甲安可誘導CpTIFY10A-like基因高效表達,致使PRSV基因表達量大幅度降低[19]。本試驗模擬蚜蟲的病毒田間傳播,在煙草TIFY 9轉基因株系和非轉基因株系移栽上苗涂抹含煙草花葉病毒的煙草試管苗等量病樣汁液,與TIFY 9非轉基因煙草相比較,TIFY 9轉基因煙草的光合參數(Pn)、氣孔導度(Gs)、蒸騰速率(Tr)、最大光化學效率(Fv/Fm)、實際光化學量子效率(ΦPSⅡ)、光化學淬滅系數(qP)及電子傳遞效率(ETR)顯著增加,而非光化學淬滅系數(NPQ)顯著下降,說明江西鉛山紅芽芋TIFY 9基因響應病毒脅迫應答可能是依賴光合作用的提高。本研究通過克隆江西鉛山紅芽芋TIFY 9基因并對其進行生物學信息、組織表達量、亞細胞定位和功能分析,為今后深入進一步探索江西鉛山紅芽芋TIFY 9基因的功能及表達蛋白互作提供理論參考,同時為研究江西鉛山紅芽芋TIFY 9基因在江西鉛山紅芽芋抗芋花葉病毒(DsMV)、黃瓜花葉病毒(CMV)、芋羽狀斑駁病毒(TFMoV)等病毒侵染過程的分子調控機理打下基礎,對江西鉛山紅芽芋脫毒育種具有重要意義。
參考文獻:
[1]劉星月,朱強龍,李慧英,等. 紅芽芋脫毒試管芋誘導及植株再生[J]. 園藝學報,2020,47(12):2427-2438.
[2]周慶紅,劉星月,王葡萄,等. 脫毒紅芽芋不同世代生長特性及產量分析[J]. 種子,2020,39(2):96-98.
[3]楊銳佳,張中保,吳忠義. 植物轉錄因子TIFY家族蛋白結構和功能的研究進展[J]. 生物技術通報,2020,36(12):121-128.
[4]Thireault C,Shyu C,Yoshida Y,et al. Repression of jasmonate signaling by a non-TIFY JAZ protein in Arabidopsis[J]. The Plant Journal,2015,82(4):669-679.
[5]Ebel C,BenFeki A,Hanin M,et al. Characterization of wheat (Triticum aestivum) TIFY family and role of Triticum Durum TdTIFY11a in salt stress tolerance[J]. PLoS One,2018,13(7):e0200566.
[6]Nishii A,Takemura M,Fujita H,et al. Characterization of a novel gene encoding a putative single zinc-finger protein,ZIM,expressed during the reproductive phase in Arabidopsis thaliana[J]. Bioscience,Biotechnology,and Biochemistry,2000,64(7):1402-1409.
[7]溫 東,王夢月,米要磊,等. 中藥火麻仁基原植物大麻的TIFY基因家族鑒定及功能分析[J]. 中國實驗方劑學雜志,2020,26(24):134-143.
[8]Ye H Y,Du H,Tang N,et al. Identification and expression profiling
analysis of TIFY family genes involved in stress and phytohormone responses in rice[J]. Plant Molecular Biology,2009,71(3):291-305.
[9]Zhu D,Bai X,Luo X A,et al. Identification of wild soybean (Glycine soja) TIFY family genes and their expression profiling analysis under bicarbonate stress[J]. Plant Cell Reports,2013,32(2):263-272.
[10]Llave C. Virus-derived small interfering RNAs at the core of plant-virus interactions[J]. Trends in Plant Science,2010,15(12):701-707.
[11]Zhang Z B,Li X L,Yu R,et al. Isolation,structural analysis,and expression characteristics of the maize TIFY gene family[J]. Molecular Genetics and Genomics,2015,290(5):1849-1858.
[12]李 云,牛麗亞,涂 瑾,等. 親水膠體對紅芽芋全粉理化特性和消化特性的影響[J]. 中國糧油學報,2020,35(2):12-17.
[13]姜紹通,程元珍,鄭 志,等. 紅芽芋營養成分分析及評價[J]. 食品科學,2012,33(11):269-272.
[14]Clarke J D,Volko S M,Ledford H,et al. Roles of salicylic acid,jasmonic acid,and ethylene in cpr-induced resistance in Arabidopsis[J]. The Plant Cell,2000,12(11):2175-2190.
[15]Vanholme B,Grunewald W,Bateman A,et al. The tify family previously known as ZIM[J]. Trends in Plant Science,2007,12(6):239-244.
[16]羅冬蘭,巴良杰,陳建業,等. 香蕉MaTIFY1轉錄因子特性及其在成熟過程中基因表達分析[J]. 園藝學報,2017,44(1):43-52.
[17]丁旭坡,馬納納,龐寬壯,等. 海南龍血樹4個DcTIFY10轉錄因子的鑒定和表達分析[J]. 分子植物育種,2019,17(19):6287-6294.
[18]Song S S,Qi T C,Fan M,et al. The bHLH subgroup IIId factors negatively regulate jasmonate-mediated plant defense and development[J]. PLoS Genetics,2013,9(7):e1003653.
[19]呂金慧,王府潤,陳 萍. 番木瓜CpTIFY10A-like基因克隆及表達分析[J]. 南方農業學報,2020,51(6):1308-1315.
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
