STAT3 miR-146-b反饋回路抑制IL-6的NF-κB信號軸調控結直腸癌細胞的干性機制
2022-04-02 10:54:07韓嘉晟吳華星袁勝春
中國醫學創新 2022年6期
關鍵詞:檢測
韓嘉晟 吳華星 袁勝春
【摘要】 目的:探究STAT3 miR-146-b反饋回路抑制IL-6的NF-κB信號軸調控結直腸癌細胞的干性機制。方法:用Dulbecco改良的Eagle培養基(DMEM)培養結直腸癌細胞HCT116和正常人結腸上皮細胞CCD 841 CoN細胞。通過免疫熒光檢測IL-6和NF-κB信號軸。通過Western blot檢測p-STAT3、p-JAK2和miR-146-b蛋白表達水平。通過Transwell分析法檢測細胞遷移能力,通過TUNEL染色檢測細胞凋亡。通過CCK-8評估不同處理組細胞活力。通過Western blot檢測EMT間充質標志物波形蛋白(Vimentin,VIM),SNAIL和ZEB1及上皮標志物E-鈣粘蛋白(CDH1)水平。通過PCR檢測CRC標志物CD44、CD133、NANOG和Lgr5mRNA的表達。結果:與對照組相比,CRC組IL-6和NF-κB表達水平均升高(P<0.05),而IL-6抑制劑組較CRC組IL-6和NF-κB表達水平均降低(P<0.05)。與對照組相比,CRC組p-STAT3、p-JAK2和miR-146-b蛋白含量均升高(P<0.05),而IL-6抑制劑組較CRC組p-STAT3、p-JAK2和miR-146-b蛋白含量均降低(P<0.05)。與對照組相比,CRC組細胞遷移能力增加,細胞凋亡降低(P<0.05),而IL-6抑制劑組較CRC組細胞遷移能力降低,細胞凋亡增加(P<0.05)。與對照組相比,CRC組0 h細胞活力無差異(P>0.05),24、48和72 h細胞活力均增加(P<0.05),而IL-6抑制劑組較CRC組0 h細胞活力無差異(P>0.05),24、48和72 h細胞活力均降低(P<0.05)。與對照組相比,CRC組VIM、SNAIL和ZEB1蛋白含量均升高,CDH1蛋白含量降低(P<0.05),而IL-6抑制劑組較CRC組VIM、SNAIL和ZEB1蛋白含量均降低,CDH1蛋白含量升高(P<0.05)。與對照組相比,CRC組CD44、CD133、NANOG和Lgr5 mRNA的表達均升高(P<0.05),而IL-6抑制劑組較CRC組CD44、CD133、NANOG和Lgr5 mRNA的表達均降低(P<0.05)。結論:STAT3 miR-146-b反饋回路抑制IL-6的NF-κB信號軸調控結直腸癌細胞的干性。IL-6/STAT3/miR-146-b軸可以作為CRC的診斷標記,并為結腸癌治療提供治療靶標。
【關鍵詞】 結直腸癌 STAT3 miR-146-b IL-6 NF-κB 增殖 凋亡
Mechanism by STAT3 miR-146-b Feedback Loop Inhibits NF-κB Signal Axis of IL-6 in Regulating Stemness of Colorectal Cancer Cells/HAN Jiasheng, WU Huaxing, YUAN Shengchun. //Medical Innovation of China, 2022, 19(06): 0-029
[Abstract] Objective: To explore the mechanism by STAT3 miR-146-b feedback loop inhibits the NF-κB signal axis of IL-6 in regulating the stemness of colorectal cancer cells. Method: Colorectal cancer cell HCT116 and Normal human colonic epithelial cells CCD 841 CoN cells were cultured with Dulbecco’s modified Eagle medium (DMEM). The IL-6 and NF-κB signal axis were detected by immunofluorescence. The expression levels of p-STAT3, p-JAK2 and miR-146-b protein were detected by Western blot. The cell migration ability was detected by Transwell analysis method, and cell apoptosis was detected by TUNEL staining. The cell viabilities of different treatment groups were evaluated by CCK-8. The levels of EMT mesenchymal markers vimentin (Vimentin, VIM), SNAIL and ZEB1 and epithelial marker E-cadherin (CDH1) were detected by Western blot. The expressions of CRC markers CD44, CD133, NANOG and Lgr5 mRNA were detected by PCR. Result: Compared with the control group, the expression levels of IL-6 and NF-κB in the CRC group were increased (P<0.05), while the expression levels of IL-6 and NF-κB in the IL-6 inhibitor group were decreased compared with the CRC group (P<0.05). Compared with the control group, the protein contents of p-STAT3, p-JAK2 and miR-146-b in the CRC group were all increased (P<0.05), while the protein levels of p-STAT3, p-JAK2 and miR-146-b in the IL-6 inhibitor group were decreased compared with the CRC group (P<0.05). Compared with the control group, the CRC group had increased cell migration ability and decreased cell apoptosis (P<0.05), while the IL-6 inhibitor group had decreased cell migration ability and increased cell apoptosis compared with the CRC group (P<0.05). Compared with the control group, there was no difference in cell viability of the CRC group at 0 h (P>0.05), and increased cell viability at 24, 48 and 72 h (P<0.05), while the IL-6 inhibitor group had no difference in cell viability at 0 h compared with the CRC group (P>0.05), 24, 48 and 72 h cells reduced vitality (P<0.05). Compared with the control group, the protein contents of VIM, SNAIL and ZEB1 in the CRC group were increased, and the CDH1 protein content was decreased (P<0.05), while the IL-6 inhibitor group had decreased VIM, SNAIL and ZEB1 protein content and increased CDH1 protein content compared with the CRC group (P<0.05). Compared with the control group, the expression of CD44, CD133, NANOG and Lgr5 mRNA in the CRC group were all increased (P<0.05), while the expression of CD44, CD133, NANOG and Lgr5 mRNA in the IL-6 inhibitor group were decreased compared with the CRC group (P<0.05). Conclusion: STAT3 miR-146-b feedback loop inhibits NF-κB signal axis of IL-6 to regulate the stemness of colorectal cancer cells. IL-6/STAT3/miR-146-b axis can be used as a diagnostic marker for CRC and provide a therapeutic target for the treatment of colon cancer.
[Key words] Colorectal cancer STAT3 miR-146-b IL-6 NF-κB Proliferation Apoptosis
First-author’s address: Liaoyang Central Hospital, Liaoning Province, Liaoyang 111000, China
doi:10.3969/j.issn.1674-4985.2022.06.006
癌癥是西方世界主要的健康問題之一,轉移是最關鍵的問題,它占癌癥死亡率的90%以上[1]。原發性結腸直腸癌(colorectal cancer,CRC)起源于胃腸道內的上皮細胞。在轉移過程中,癌細胞被認為具有間充質表型,從而使它們離開原發腫瘤的部位,侵入周圍組織,并遷移到遠處的器官。接種后,這些細胞轉換回上皮表型并增殖形成轉移灶。細胞在上皮和間充質表型之間轉換的過程稱為上皮-間充質轉化(epithelial-mesenchymal transition,EMT)及其對應的間充質-上皮轉化(mesenchymal-epithelial transition,MET)。炎癥是在受傷后開始的生物學過程,以去除有害刺激并開始愈合過程,越來越多的證據表明炎癥會促進EMT[2]。但是,如果炎癥延長,則可能對機體有害并促進包括癌癥在內的永久性疾病。大約25%的癌癥是由于慢性感染或其他類型的慢性炎癥而出現的。例如,炎癥性腸病或結腸炎是CRC的重要危險因素。IL-6是炎癥促腫瘤作用的重要介質,IL-6是由幾種類型的免疫細胞和癌產生的促炎細胞因子,尤其是在腫瘤發生的后期[3]。血清中IL-6水平的升高與晚期階段、腫瘤大小、轉移和CRC患者生存率降低相關,但對這些關聯的分子機制尚不甚了解。大量研究表明,微小RNA(microRNA,miRNA)在代表癌癥特征的生物過程中具有重要的調控功能,包括癌細胞的增殖、凋亡、侵襲和轉移。癌癥干細胞(CSC)的許多標志物取決于癌癥類型,例如在結腸癌中,據報道細胞表面標志物如CD133、CD44和ALDH是鑒定CSC的一類分子[4]。STAT3是STAT家族之一,參與細胞生長和存活、炎癥、免疫反應和癌癥進展。STAT3通過激活非受體蛋白酪氨酸激酶[如Janus激酶1(Janus kinase 1,JAK1)、JAK2、JAK3、EGFR和SRC信號傳導]與核輸入蛋白相互作用而二聚并進入細胞核。本文主要研究STAT3 miR-146-b反饋回路抑制IL-6的NF-κB信號軸調控結直腸癌細胞的干性的機制。現報道如下。
1 材料與方法
1.1 細胞培養 結直腸癌細胞HCT116(ATCC?HTB-247?)購自美國典型培養物保藏中心(ATCC公司,美國)。正常人結腸上皮細胞CCD 841 CoN購自賽默飛世爾科技(中國)有限公司(美國)。用Dulbecco改良的Eagle培養基(DMEM)培養HCT116和CCD 841 CoN細胞。此外,對于HCT116細胞系,應添加10%FBS和1%青霉素/鏈霉素(Gibco公司,美國)。正常結腸上皮細胞也在DMEM中生長。15%FBS,4 mM L-谷氨酰胺和1%青霉素/鏈霉素對于這些細胞的生長是必需的。具有5%CO2濕潤氣氛的細胞培養箱適用于所有細胞培養。
1.2 分組 對照組:正常結腸上皮細胞;CRC組:結直腸癌細胞;IL-6抑制劑組:經500 mg/mL的IL-6抑制劑(美國Sigma公司)處理結直腸癌細胞48 h。每組3個復孔。利用以上各組材料進行后續檢測分析。
1.3 免疫熒光 不同處理組結直腸癌細胞用4%甲醛固定,在0.1%Triton X-100中透化。將固定的結直腸癌細胞用1×PBS洗滌,并在室溫下用1×PBS中的2%牛血清白蛋白(BSA)封閉30 min。將固定的細胞與IL-6或NF-κB抗體的特異性一抗(Cell Signaling公司,美國)在4 ℃孵育過夜。洗滌后,將細胞與Alexa Fluor 546山羊兔IgG抗體(Life Technologies公司,美國)(1︰1 000)在室溫下孵育1 h。洗滌兩次后,用4,6-二mid基-2-苯基吲哚(DAPI,Sigma公司,美國)將細胞核染色。STAT3和DAPI染色的細胞圖像是通過Delta Vision成像系統(Applied Precision公司,美國)拍攝的。
1.4 蛋白質印跡 將HCT116細胞在含有磷酸酶抑制劑(Sigma公司,美國)的RIPA緩沖液(50 mM Tris-HCl,150 mM NaCl,2 mM EDTA和1%Triton X-100)中裂解。蛋白酶抑制劑(Roche公司,德國)。將蛋白質樣品在8%~15%的十二烷基硫酸鈉-聚丙烯酰胺凝膠(SDS-PAGE)上分離,并轉移到硝酸纖維素膜上。將膜與PARP的一抗,裂解的PARP,裂解的p-STAT3、p-JAK2、miR-146-b、VIM、SNAIL、ZEB1、CDH1蛋白和GAPDH一起孵育(Sigma公司,美國)。將它們在3%牛血清白蛋白(BSA)和PBS-Tween 20(1︰500~1︰2 000)中于4 ℃稀釋過夜,用PBS-Tween20洗滌3次,最后與HRP耦聯的二抗孵育(1︰2 000)。通過使用ECL Western印跡檢測試劑(GE Healthcare公司,英國)將表達可視化。
1.5 Transwell遷移測定 CRC細胞遷移的能力通過Transwell分析法(Thermo公司,美國)確定,將癌細胞鋪在帶有可滲透膜的細胞培養插入物的上層,將測試劑置于細胞滲透膜下方,將細胞孵育3~18 h并計數細胞。
1.6 TUNEL檢測 為了檢測細胞死亡,根據制造商的說明(Roche Molecular Biochemicals公司,德國)使用了DeadEnd?熒光TUNEL系統套件(Promega公司,美國)。將HCT116細胞用IL-6抑制劑處理24 h,用冷PBS洗滌。用4%多聚甲醛固定細胞30 min,并用PBS洗滌兩次,2 min/次。洗滌通透溶液(0.1%Triton X-100和0.1%檸檬酸鈉)中的固定細胞,并與TUNEL測定混合物孵育60 min。TUNEL染色的細胞通過Delta Vision成像系統(Applied Precision公司,美國)可視化。
1.7 CCK8分析 根據Cell Counting Kit-8分析(CCK-8)試劑盒(Dojindo公司,日本)評估細胞活力。將2×103細胞接種到96孔板中,并在37 ℃的含有5%CO2的潮濕室內孵育24、48或72 h。將CCK-8溶液(10 μL)加入每個孔中,并將板在37 ℃下孵育1 h。在酶標儀(Bio-Rad,美國)中測量450 nm處細胞的吸光度(OD450)。
1.8 RNA分離和定量實時聚合酶鏈反應(PCR) 使用ReliaPrep RNA Cell Miniprep System(Promega Corp.,Z6010,Madison,WI,United States)從HCT116細胞中提取總RNA 1 μg,以總RNA為模板,通過逆轉錄酶合成cDNA,再以cDNA為模板,采用Superscript One Step RT-PCR試劑盒(Invitrogen公司,美國)進行Platinum Taq聚合酶擴增。HB Nucleic mix(Heimbiotek公司,韓國)合成的引物序列。為了進行PCR擴增,進行以下步驟。最初的步驟是在50 ℃下進行30 min,在94 ℃下進行2 min,在94 ℃下進行30次循環15 s,在55 ℃下進行30 s,在72 ℃下進行1 min,最后一步在72 ℃下進行10 min。使用HB Real-Time PCR預混液試劑盒(Heimbiotek公司,韓國)進行RT-qPCR。
1.9 統計學處理 采用SPSS 20.0軟件對所得數據進行統計分析,計量資料用(x±s)表示,組間比較采用單因素方差分析或者重復測量的方差分析,組間兩兩比較采用LSD-t檢驗;計數資料以率(%)表示,比較采用字2檢驗。以P<0.05為差異有統計學意義。
2 結果
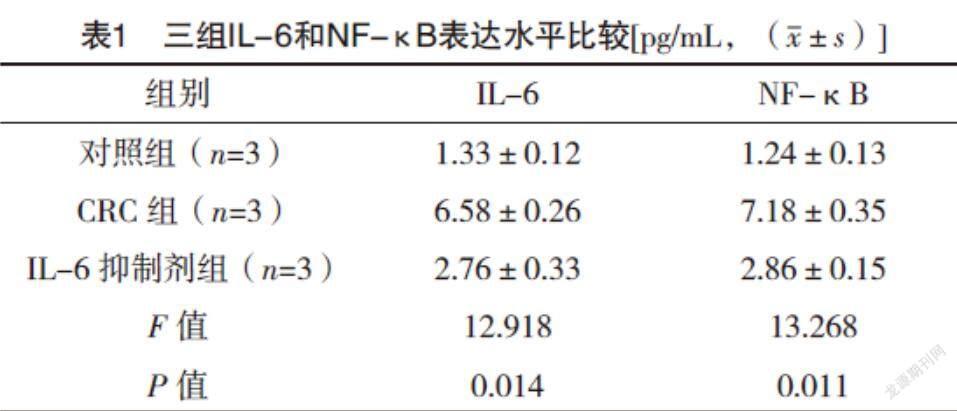
2.1 免疫熒光檢測IL-6和NF-κB信號軸 與對照組相比,CRC組IL-6和NF-κB表達水平均升高(P<0.05),而IL-6抑制劑組較CRC組IL-6和NF-κB表達水平均降低(P<0.05),見表1。
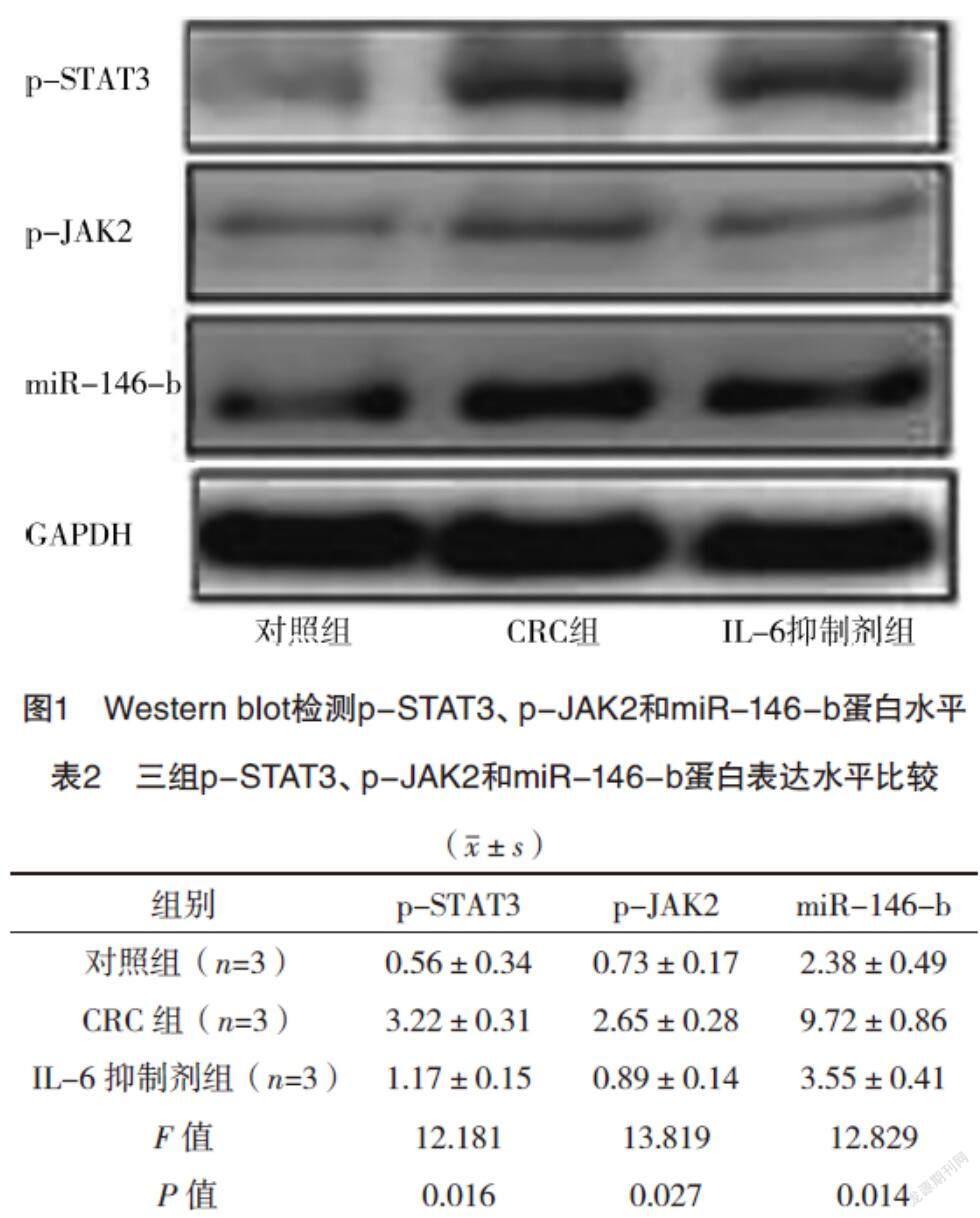
2.2 Western blot檢測p-STAT3、p-JAK2和miR-146-b蛋白水平 與對照組相比,CRC組p-STAT3、p-JAK2和miR-146-b蛋白含量均升高(P<0.05),而IL-6抑制劑組較CRC組p-STAT3、p-JAK2和miR-146-b蛋白含量均降低(P<0.05),見圖1、表2。
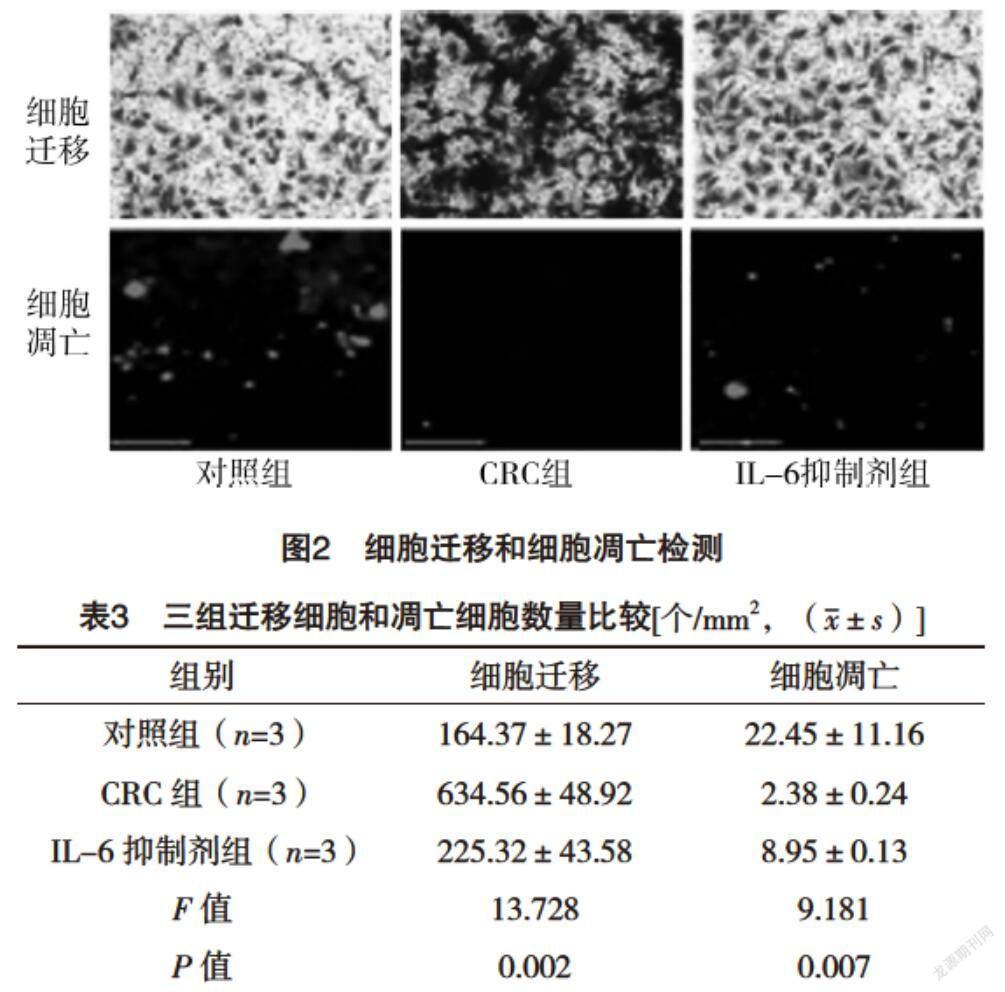
2.3 細胞遷移和細胞凋亡測定 與對照組相比,CRC組細胞遷移能力增加,細胞凋亡降低(P<0.05),而IL-6抑制劑組較CRC組細胞遷移能力降低,細胞凋亡增加(P<0.05),見圖2、表3。
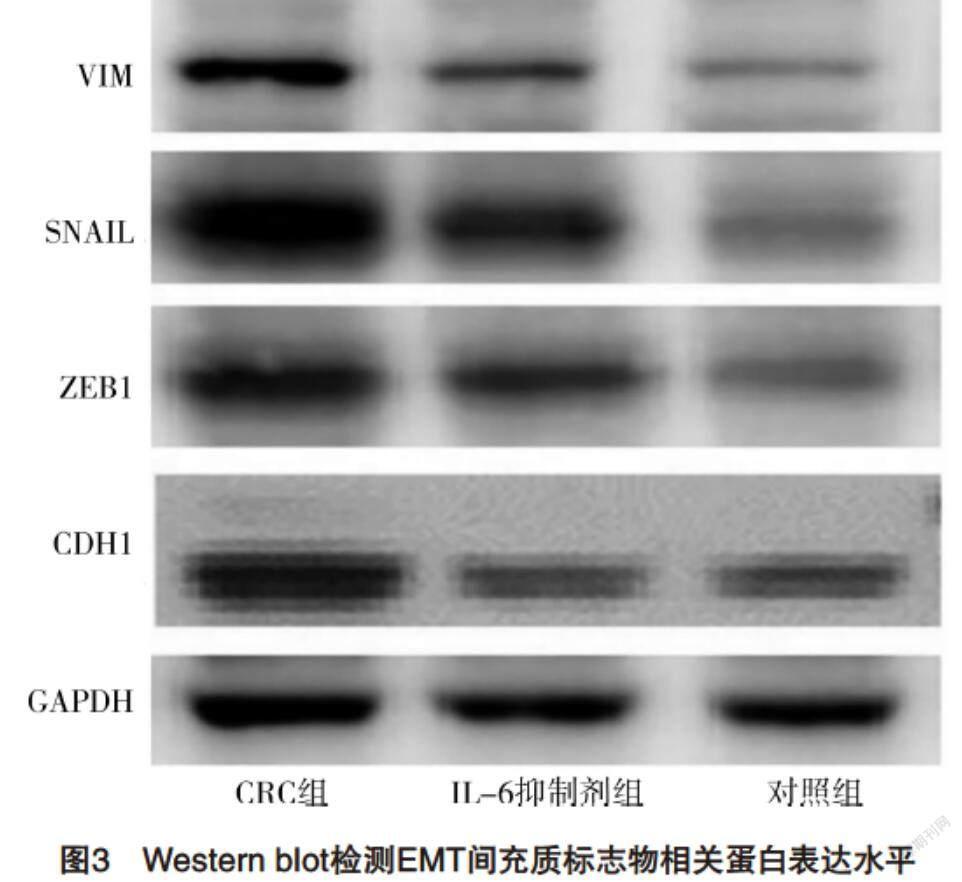
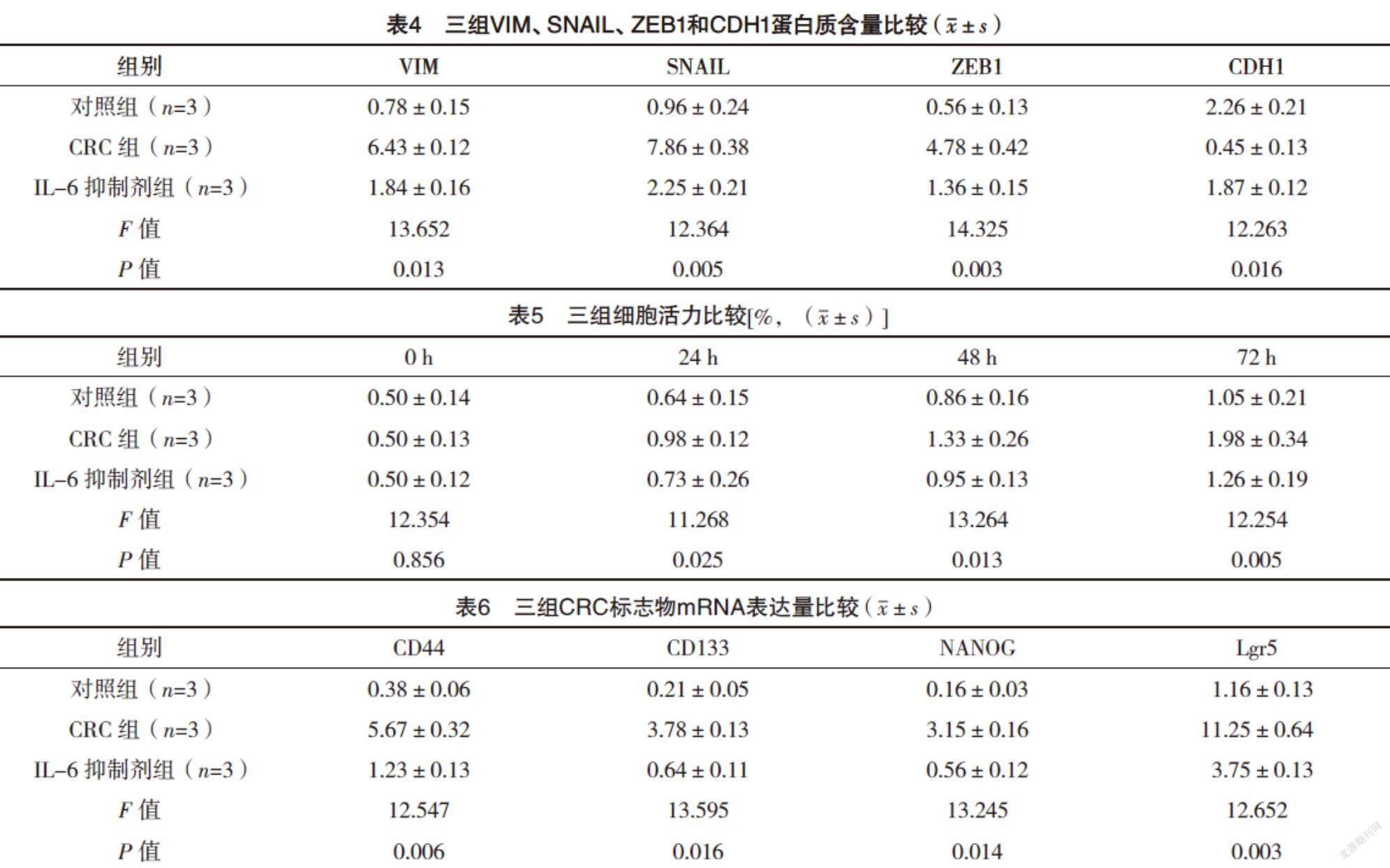
2.4 Western blot檢測EMT間充質標志物相關蛋白表達水平 與對照組相比,CRC組VIM、SNAIL和ZEB1蛋白含量均升高,CDH1蛋白含量降低(P<0.05),而IL-6抑制劑組較CRC組VIM、SNAIL和ZEB1蛋白含量均降低,CDH1蛋白含量升高(P<0.05),見圖3、表4。
2.5 CCK8分析細胞活力 CRC組0 h細胞活力與對照組相比,差異無統計學意義(P>0.05),24、48和72 h細胞活力較對照組均增加(P<0.05);而IL-6抑制劑組與CRC組0 h細胞活力比較,差異無統計學意義(P>0.05),24、48和72 h細胞活力較CRC組均降低(P<0.05)。見表5。
2.6 三組CRC標志物mRNA表達量比較 與對照組相比,CRC組CD44、CD133、NANOG和Lgr5 mRNA的表達均升高(P<0.05),而IL-6抑制劑組較CRC組CD44、CD133、NANOG和Lgr5 mRNA的表達均降低(P<0.05),見表6。
3 討論
CRC是第三大常見癌癥,約占所有癌癥病例的10%,全世界每年診斷出的新病例超過100萬[5]。手術是結直腸癌和化學療法的主要治療方法,分子靶向治療是輔助治療。越來越多的證據表明,在早期發現的CRC患者中大約有90%可以通過手術治愈,但是不幸的是,大多數患者被診斷為晚期,導致預后不良[6]。因此,迫切需要找到新的診斷和預后標志物。
miRNA是一種小型非編碼RNA,它們通過與3’非翻譯區(UTR)結合而與其靶基因mRNA相互作用,可減少蛋白質豐度并調節多種信號通路,與血管生成、增殖、骨重塑和癌癥進展有關[7]。據報道,許多miRNA可以調節多種生物學過程,例如增殖、分化、遷移和凋亡。此外新出現的證據表明,miRNA是用于癌癥診斷和預后的有前途的生物標志物。miRNA經常形成反饋環,因為它們本身受轉錄因子調控,它們直接或間接靶向[8]。這種自我穩定的電路可以是表觀遺傳開關的核心組成部分,其中細胞表型和表達模式從一種穩定的表觀遺傳狀態轉換為另一種,而DNA序列沒有變化[9]。表觀遺傳開關需要啟動事件(炎癥),但是新細胞類型的表型在沒有啟動信號的情況下遺傳并通過自我維持的反饋回路維持[10]。
大腸癌是全世界與癌癥相關的死亡的主要原因之一[11]。幾種致癌分子與大腸癌的發展有關,包括信號轉導子和STAT3[12]。本研究中,確定了一個IL-6觸發的反饋環,該環涉及STAT3介導的miR-146-b抑制,并控制EMT和結腸腫瘤的進展。筆者分析顯示,IL-6/STAT3/miR-146-b環存在于所有具有間充質并因此具有轉移性狀的癌細胞中,這暗示它可能代表了一種新的致癌統一機制。盡管如此,IL-6/STAT3/miR-146-b環的活化可能存在組織和細胞類型特異性差異,因為STAT3活化可以通過多種其他信號途徑來實現,例如在癌癥中失活的受體酪氨酸激酶[13]。最近的觀察表明,在轉移過程中,腫瘤細胞獲得了一種間充質表型,從而使它們離開原發腫瘤的部位,侵入周圍組織,并遷移到遠處的器官,而在播種后,這些細胞又轉換回上皮表型,從而有效地轉移[14-15]。IL-6對IL-6/STAT3/miR-146-b環的激活會誘導EMT,并可能使細胞表型向間質狀態轉移,這有利于轉移級聯的浸潤,內滲和外滲步驟[16]。相反,IL-6抑制劑對環的干擾會誘導MET,并可能使細胞轉換回上皮狀態,從而造成轉移的集落和向外生長[17]。IL-6/STAT3/miR-146-b環的可逆特性有望實現旨在阻止其前轉移活性的多種治療干預。
IL-6/STAT3/miR-146-b環成分的幾個已知靶標可能對癌癥進展很重要,例如STAT3可以直接誘導EMT激活劑ZEB1。此外,文獻[18]表明,另一種公認的EMT誘導物SNAIL是miR-146-b的直接靶標。其他研究表明,SNAIL誘導IL-6,從而提供可能增強循環的其他反饋[19]。總的來說,這些結果表明SNAIL是IL-6/STAT3/miR-146-b環的重要效應子。IL-6還可以通過STAT3/SNAIL信號通路促進頭頸部腫瘤轉移和EMT。
CSC取決于癌癥類型,占據不同的標志物,例如SOx2、CD133、EpCAM、CD166、CD24、CD29、Lgr5、Klf4和ALDH[20]。本研究結果顯示,與對照組相比,CRC組CD44、CD133、NANOG和Lgr5 mRNA的表達均升高,而IL-6抑制劑組較CRC組CD44、CD133、NANOG和Lgr5 mRNA的表達均降低。
綜上所述,STAT3 miR-146-b反饋回路抑制IL-6的NF-κB信號軸調控結直腸癌細胞的干性。IL-6/STAT3/miR-146-b軸可以作為CRC的診斷標記,并為結腸癌治療提供治療靶標。
參考文獻
[1] ABDULRIDHA M K,AL-MARZOQI A H,GHASEMIAN A.The Anticancer Efficiency of Citrullus colocynthis Toward the Colorectal Cancer Therapy[J].J Gastrointest Cancer,2020,51(2):439-444.
[2] PERIASAMY S,WU W H,CHIEN S P,et al.Dietary Ziziphus jujuba Fruit Attenuates Colitis-Associated Tumorigenesis:A Pivotal Role of the NF-κB/IL-6/JAK1/STAT3 Pathway[J].Nutr Cancer,2020,72(1):120-132.
[3] LI Z W,SUN B,GONG T,et al.GNAI1 and GNAI3 Reduce Colitis-Associated Tumorigenesis in Mice by Blocking IL6 Signaling and Down-regulating Expression of GNAI2[J].Gastroenterology,2019,156(8):2297-2312.
[4] LEE J W,STONE M L,PORRETT P M,et al.Hepatocytes direct the formation of a pro-metastatic niche in the liver[J].Nature,2019,567(7747):249-252.
[5] XU K,ZHAN Y,YUAN Z,et al.Hypoxia Induces Drug Resistance in Colorectal Cancer through the HIF-1α/miR-338-5p/IL-6 Feedback Loop[J].Mol Ther,2019,27(10):1810-1824.
[6] WANG T,SONG P,ZHONG T,et al.The inflammatory cytokine IL-6 induces FRA1 deacetylation promoting colorectal cancer stem-like properties[J].Oncogene, 2019, 38(25):4932-4947.
[7] WANG S,DONG W,LIU L,et al.Interplay between bile acids and the gut microbiota promotes intestinal carcinogenesis[J].Mol Carcinog,2019,58(7):1155-1167.
[8] KUANG Y,GUO W,LING J,et al.Iron-dependent CDK1 activity promotes lung carcinogenesis via activation of the GP130/STAT3 signaling pathway[J].Cell Death Dis,2019,10(4):297.
[9] WANG J,ZHOU J,JIANG C,et al.LNRRIL6,a novel long noncoding RNA,protects colorectal cancer cells by activating the IL-6-STAT3 pathway[J].Mol Oncol,2019,13(11):2344-2360.
[10] XU J,QIAN J,ZHANG W,et al.LYPD8 regulates the proliferation and migration of colorectal cancer cells through inhibiting the secretion of IL-6 and TNF-α[J].Oncol Rep,2019,41(5):2389-2395.
[11] FANG W,ZHU S,NIU Z,et al.The protective effect of syringic acid on dextran sulfate sodium-induced experimental colitis in BALB/c mice[J].Drug Dev Res,2019,80(6):731-740.
[12] WEI J,MA L,LAI Y H,et al.Bazedoxifene as a novel GP130 inhibitor for Colon Cancer therapy[J].J Exp Clin Cancer Res,2019,38(1):63.
[13] ZHANG B,XU Y,LIU S,et al.Dietary Supplementation of Foxtail Millet Ameliorates Colitis-Associated Colorectal Cancer in Mice via Activation of Gut Receptors and Suppression of the STAT3 Pathway[J].Nutrients,2020,12(8):2367.
[14] TIAN Y,XU J,LI Y,et al.MicroRNA-31 Reduces Inflammatory Signaling and Promotes Regeneration in Colon Epithelium,and Delivery of Mimics in Microspheres Reduces Colitis in Mice[J].Gastroenterology,2019,156(8):2281-2296.
[15] WANG A,DENG S,CHEN X,et al.miR-29a-5p/STAT3 Positive Feedback Loop Regulates TETs in Colitis-Associated Colorectal Cancer[J].Inflamm Bowel Dis,2020,26(4):524-533.
[16] HUANG H,NIU J,WANG F,et al.A natural compound derivative P-13 inhibits STAT3 signaling by covalently inhibiting Janus kinase 2[J].Invest New Drugs,2019,37(3):452-460.
[17] RAJ V,BHADAURIA A S,SINGH A K,et al.Novel 1,3,4-thiadiazoles inhibit colorectal cancer via blockade of IL-6/COX-2 mediated JAK2/STAT3 signals as evidenced through data-based mathematical modeling[J].Cytokine,2019,118(23):144-159.
[18] FUJITA J,SAKURAI T.The Oncoprotein Gankyrin/PSMD10 as a Target of Cancer Therapy[J].Adv Exp Med Biol,2019,1164(5):63-71.
[19] KE Z,WANG C,WU T,et al.PAR2 deficiency enhances myeloid cell-mediated immunosuppression and promotes colitis-associated tumorigenesis[J].Cancer Lett,2020,469(15):437-446.
[20] MA H,WANG J,ZHAO X,et al.Periostin Promotes Colorectal Tumorigenesis through Integrin-FAK-Src Pathway-Mediated YAP/TAZ Activation[J].Cell Rep,2020,30(3):793-806.
(收稿日期:2021-06-24) (本文編輯:程旭然)
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
