中國水稻百年育種的一些關鍵基因
2022-04-06 01:22:08劉定富尹合興應繼鋒
中國稻米 2022年2期
關鍵詞:水稻
劉定富 尹合興 應繼鋒
(1 武漢金玉良種科技有限公司,武漢 430064;2 華智生物技術有限公司,長沙 410125;3 中國水稻研究所,杭州 310006;*通訊作者:hexing.yin@higentec.com;yingjifeng@caas.cn)
水稻育種就是培養新的或者改良性狀的品種。傳統意義上的育種就是培育“具有一致性、優良性、穩定性的生物群體”。因此,育種的生物學本質就是聚合有利基因型或有利基因型集合。雖然水稻有近萬年的人工栽培歷史,但是真正意義上的育種只有100年左右時間,即使把人工介入的植物馴化包括在內,也不過2 000~3 000年的歷史,更多的則是自然進化[1]。人類賴以生存的主要農作物的萬年演變進化,從遺傳學本質上講,其實就是基因的被動負向淘汰到更多的基因正向選擇逐步轉變。
第一代(1 G=Generation)育種技術——馴化,大約從1 萬年前開始,早期的農民并不懂得遺傳多樣性的理論,但已經開始有意識或無意識對植物進行偶然的選擇,他們會選擇產量或其他性狀表現好的單株作為下一季栽培的種子,并不斷繁殖下去。第二代育種技術(2G)——雜交育種技術,始于19 世紀中后期,在孟德爾發現了植物遺傳定律(1865年左右)后,數量遺傳學理論被建立起來,育種家和專業的科學家通過人工雜交的手段,有目的地選配不同親本進行雜交、自交、回交等,結合雙親的優良性狀培育或改良作物品種。第三代育種技術(3G)——傳統育種技術,包括雜種優勢育種及主動誘變育種。19 世紀末至20 世紀初,英國的一些種子公司開始對植物進行復合雜交(主要是玉米),并從中選擇突破性品種。玉米雜種優勢和雙雜交種在商業化上突出的應用表現帶動了雜種優勢在水稻、高粱、油菜、棉花等其他作物上的運用。1940年,物理、化學或太空誘變等手段在作物育種上開始應用,主動的誘變育種可以創造全新的變異,還能促進遠緣雜交過程中染色體的變異,對于豐富遺傳性狀具有重要意義。第四代育種技術(4G)——分子育種技術,得益于現代分子生物學、基因工程的發展,自20 世紀80年代開始,以轉基因(GMO,Genetic Modified Organism)、分子標記輔助選擇(MAS,Marker Assisted Selection)、全基因組選擇(GS,Genome Selection)、等位基因挖掘等為代表的現代分子技術手段開始在作物育種上運用。GMO 已經發展成最快、應用效率最高的精準育種技術之一。目前在北美地區,90%以上的玉米、大豆作物的抗病、抗蟲、抗除草劑性狀得到應用并取得了商業化成功。MAS 是20 世紀80年代興起的DNA 標記技術,以QTL 作圖和RFLP、SSR、SNP 等分子標記為基礎,經歷了標記開發、遺傳圖譜、功能和比較基因組連鎖分析及基因組測序等不同發展時期。其基本原理是利用與目標基因緊密連鎖或表現共分離的分子標記對選擇個體進行目標以及全基因組篩選,從而減少連鎖累贅,獲得目標個體。GS 是MAS 的延伸,是近年來動、植物分子育種的全新策略,已成為分子技術育種的熱點和趨勢。分子技術育種是對傳統育種理論和技術的重大突破,實現了對基因的直接選擇和有效聚合,大幅度縮短了育種年限,極大地提高了育種效率。目前,各國對QTL、MAS、GS 和基因定位等精準育種的理論和試驗研究很多。許多主糧作物的關鍵性基因被定位、挖掘、克隆。第五代育種技術(5G)也是我們常說的智能育種[1]。基本定義為利用農作物基因型、表型、環境、遺傳資源(例如水稻上的品種系譜信息)等大數據為核心基礎,通過人工生物智能技術,在實驗室設計培育出一種適合于特定地理區域和環境下的品系品種。而傳統上的大田僅僅作為品種測試和驗證的場所,從而節省了大量的人力、物力、財力、環境等資源。智能育種是依托多層面生物技術和信息技術,跨學科、多交叉的一種育種方式[1-2]。
水稻進化研究表明,栽培稻(Oryza sativa)是由野生稻(Oryza rufipogon)馴化而來。馴化的主要性狀包括株型、落粒性、芒的長短和有無、異交率、產量、休眠性等。控制這些性狀的基因有:從匍匐生長到直立生長的基因PROG1[4],從長芒到短芒或無芒的基因LABA1 和GAD1[5],從極易落粒到不易落粒的基因sh4 和SHA1,從散穗到緊穗的基因OsLG1,控制粒色的基因Rc 和Rd,控制粒寬(chr1)、粒質量(chr6)、柱頭外露(chr8)等性狀的基因[6]。水稻數千上萬年的漫長進化,都只是少數幾個主效基因起著作用,而最近百余年的水稻育種,研究結果初步表明,也只有一些關鍵基因發揮了重要作用。
本綜述的目的是回顧百年水稻育種進程中的一些關鍵基因,評價和討論幾次水稻育種技術跨越式發展中起主導作用的一些關鍵性基因,并預測今后突破性水稻育種技術中的潛力基因。
1 已經發生的水稻育種三次跨越式突破
水稻育種的第一次跨越式突破“矮稈育種”的標志性成果雖然在國際上公認是國際水稻研究所1966年育成的IR8,但在1959年我國水稻育種家黃耀祥就通過雜交育種的方法育成了矮稈品種廣場矮,1956年廣東省農民洪春利、洪群英發現并育成了矮腳南特[3]。IR8、廣場矮、矮腳南特可以作為矮稈育種的代表性品種,來探討其基因的變化(表1)。
水稻育種的第二次跨越式突破“三系法雜交水稻”也是中國率先取得成功,并至今處于世界領導地位。水稻育種的第三次突破“兩系法雜交水稻”,從實踐到理論的初期開創是以湖北水稻育種家石明松為代表完成的。水稻育種的“三次跨越式突破”加上矮化育種之前的“高稈時代”,中國水稻育種或者種業的發展,基本經歷了高稈、矮稈、三系、兩系等四個階段[3]。
1.1 矮稈水稻育種(sd1)
我國自20 世紀20年代末開展水稻雜交育種,直至50年代仍然局限于高稈品種,后期倒伏是高稈品種高產的一個關鍵限制因子。20 世紀50年代中期開始的矮稈育種,其標志性事件有:
廣東省農業科學院黃耀祥團隊利用馬來西亞引進稻種矮仔占矮源開展育種工作,用系統育種方法選育出矮仔占4 號,并以此為親本與高稈品種廣場13 雜交,于1959年培育出第一個以雜交育種方法育成的矮稈品種廣場矮。
1955年洪群英、洪春利在南特號大田中發現株高僅70 cm 的矮稈變異株,穗大粒多高產,命名為矮腳南特并推廣應用。同期,臺灣學者也在開展矮稈水稻育種。1949年,臺灣臺中區農業試驗場洪秋增以低腳烏尖為母本、菜園種為父本配組,1956年育成臺中在來1號,并于1960年推廣。
這些事件引起了中國水稻生產的第一次革命,此后,水稻育種進入矮稈時代。同一時期,在國際水稻研究所,JENNINGS 用臺灣學者張德慈提供的矮源材料低腳烏尖,與印尼高稈高產的Peta 雜交,BEACHELL完成選育工作,1966年育成IR8,因為其高產優質,被稱為“奇跡稻”。此后連同矮稈小麥一起,被稱之為“綠色革命”[7]。
從高稈到矮稈,水稻進入2G 時代。該時代的品種,解決了不抗倒、不耐肥的問題,縮短了生育期,提高了產量,品種分蘗力也有所增強。
水稻2G 時代的品種類型主要是矮稈高產常規品種,育種方法主要是雜交選育,利用的變異主要是基因重組,育種理論主要是孟德爾遺傳學(基因獨立分配、自由組合和連鎖交換)。據統計,已發掘命名的水稻矮稈或半矮稈基因已有70個以上,不過有一些是等位的。主導矮稈革命的基因是半矮稈基因sd1。
sd-1 控制水稻的株高,在水稻矮化育種中得到了廣泛的應用。但是直到2002年sd1 才被克隆,被定位在水稻第1 染色體上,總長度2 742 bp,是由3個外顯子和2個內含子組成的開放閱讀框,該基因參與赤霉素的生物合成,編碼由389個氨基酸組成的GA20 氧化酶(GA20ox)。GA20ox 是赤霉素合成途徑中的關鍵酶,催化GA53 轉換為GA20。
通過對矮稈品種中sd1 的比較測序,發現sd1 的突變主要有7 種等位基因型,分別為IR8 的383 bp 缺失等位基因型、9311 外顯子提前終止等位基因型、矮腳南特中第一個外顯子中2 bp 缺失等位基因型,以及4 種SD1 氨基酸改變的等位基因型,它們的廣泛利用推動了矮稈育種的進程。sd1 不僅在“第一次綠色革命”中得到選擇應用,而且在更早的粳稻馴化過程中就被選擇;粳稻中2個功能性的FNPs(SD1-EQ) 導致GA20ox2 酶活性降低,內源GA 含量降低,最終株高降低,而秈稻和野生稻攜帶了強功能型的SD1(SD1-GR),內源GA 含量升高,株高增加。這8個突變的詳細情況歸納于表1。

表1 sd1 基因座上的等位變異
從表1 可見,sd1 基因座存在較多的等位變異,每個變異都影響赤霉素的生物合成,最終影響水稻的株高。
研究表明,除了影響赤霉素合成的基因(sd1,d18,d35 等)影響株高外,還有影響赤霉素信號傳導的基因(d1,GID2,GID1,OsSPY 等)、影響 BR 合成的基因(brd1,brd2,d2,dl1,OsDWARF4 等)、影響 BR 信號傳導的基因(d61 等)、多蘗矮稈基因(D3,HTD3,D27 等)影響水稻的株高,僅已知的就多達70 多個。不過,在生產上應用的主要是sd1 基因。可以說,sd1 實現了水稻育種的第一次突破[7]。
1.2 三系雜交稻育種(Rf3 和Rf4)
雜交育種的變異來源是有利基因的重組,其品種的優良表現依賴的是基因座位內等位基因的顯隱關系和基因座位之間基因的互作。有利基因數目的有限性,導致了產量等性狀改良出現了“瓶頸”,人們又開始尋求新的途徑實現突破,探索品種間雜種優勢利用成為一種途徑。
我國水稻雜種優勢理論始于1964年雜交水稻之父——袁隆平院士的開創性研究,但之后多年一直沒有實踐上的突破。直到1970年袁隆平助手李必湖在海南發現花粉敗育的野生稻,經全國大協作,三系法雜交水稻育種取得成功。1973年江西顏龍安最先選育出野敗秈型不育系珍汕97A,同年張先程最先發現野敗秈型不育系的強優勢恢復系IR24,幾乎同時,袁隆平團隊也培育出強優勢組合南優2 號(二九南1 號A/IR24)。我國野敗秈型雜交水稻“三系”完成配套,并選配出若干強優勢組合,導致了中國水稻育種的第2 次突破。1976年秈型雜交稻開始在全國大面積推廣,成為世界上第一個成功進行水稻雜種優勢商品化利用的國家。我國水稻育種進入3G 時代[7]。
水稻3G 時代的品種類型主要是矮稈高產三系雜交品種,育種方法主要是三系法育種,利用的變異主要是雜種優勢,育種理論主要是細胞質雄性不育理論和雜種優勢學說,主導三系革命的基因是水稻“野敗”細胞質基因和雄性不育恢復基因Rf3 和Rf4。水稻3G 時代品種大大增強了分蘗力,提高了耐肥能力,大幅提高了產量,缺點是降低了品質。
水稻野敗型細胞質雄性不育恢復基因Rf3 和Rf4能恢復水稻野敗型細胞質雄性不育系的育性。Rf3 和Rf4 定位研究的主要結果匯總于表2。野敗型細胞質雄性不育的分子機理得到解析。劉耀光教授課題組報道了一個野生稻線粒體中新近起源的基因WA352,與核編碼的線粒體蛋白COX11 互作,共同調控水稻野敗型細胞質雄性不育性(CMS-WA)。WA352 誘導雄性不育可以被 Rf3 和 Rf4 恢復,Rf3 暫時未被克隆,Rf4 編碼一個 PPR 蛋白,可以降低 WA352 的表達[7]。

表2 Rf3 和Rf4 定位研究結果
1.3 兩系雜交稻育種(pms3,tms5)
1973年石明松在晚粳農墾58 大田中發現3 株雄性不育株,第2年利用其自然結實的種子種植了48株,有雄性不育、可育兩種類型。此后6年,他對不育株進行測交和回交時發現,不育株的再生分蘗上能自交結實,分期播種的結果表明育性與光照長度有關。由此,他于1981年提出了水稻育種的全新設想:在長日高溫下制種,在短日低溫下繁殖,一系兩用,提出“兩用核不育系”概念,劃時代的“兩系法雜交水稻育種技術”由此開始[7]。
兩系法雜交稻的育性受核基因控制,沒有恢保關系,配組自由;稻種資源利用率高,選育出優良組合機率高;親本種子繁育程序簡單,成本低。1986年國家863 項目開展兩系法雜交水稻全國育種攻關,經過20多年的協作努力,建立了光溫敏不育系的兩系法雜種優勢有效利用的新途徑,解決了三系法雜交稻的主要限制因素,使水稻雜種優勢利用進入一個新階段。秈粳亞種間雜交,使得水稻產量、品質又上了一個新臺階,培育出了若干超級稻品種,這可謂是中國水稻研究和生產的第3 次大突破,中國水稻育種進入4G 時代。
水稻4G 時代的品種類型主要是矮稈高產兩系雜交品種,育種方法主要是兩系法育種,利用的變異也主要是雜種優勢,育種理論主要是光溫敏核不育理論和雜種優勢學說,主導兩系法育種的基因是光溫敏雄性核不育基因pms3 或溫敏雄性核不育基因tms5。水稻4G 時代的品種進一步提高了產量,增強了抗病能力,稻米品質有了一定改善。
其后,很多學者對pmc3 和tms5 展開了深入的研究。pms3 和tms5 基因的定位研究結果匯總于表3。
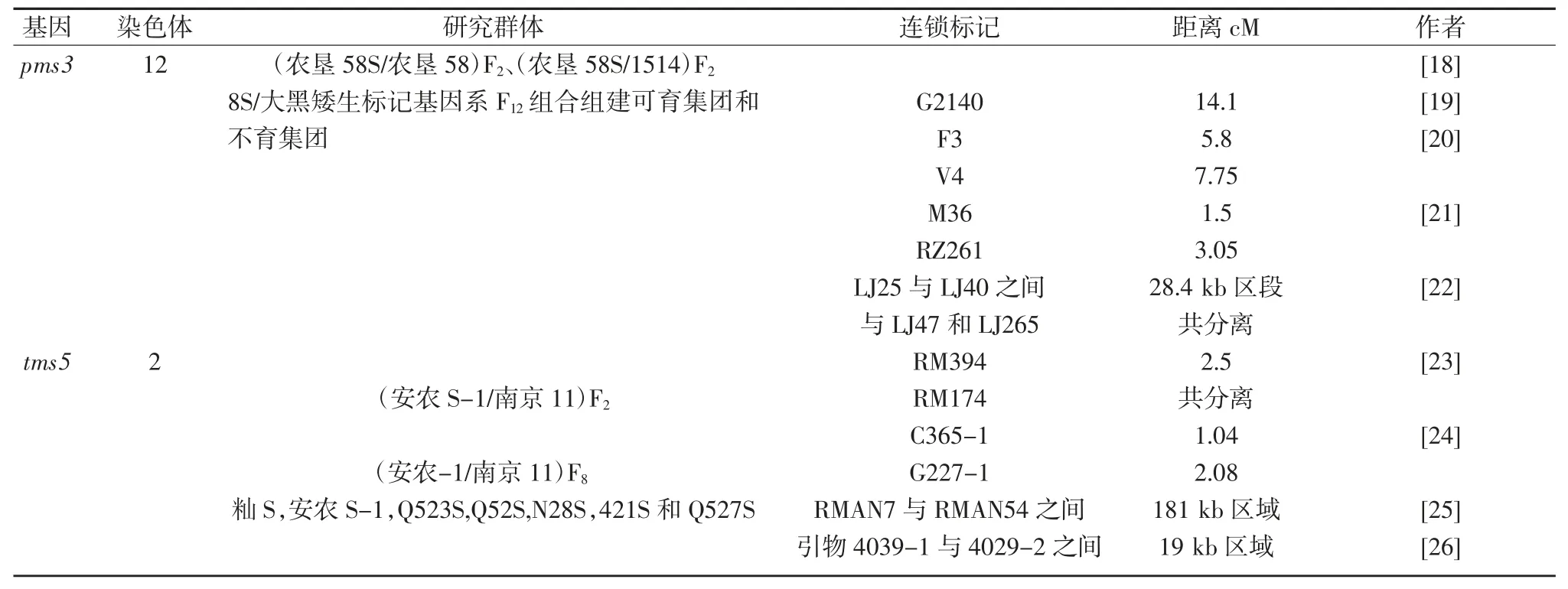
表3 pmc3 和tms5 定位研究結果
2 已基本應用的其他水稻重要基因
2.1 抗稻瘟病育種基因Pi
抗病育種是最為經濟、環保的植保方法。20 世紀60年代中期,日本率先開展了水稻品種抗稻瘟病基因的研究工作,鑒定了最初的8個抗性位點上的14個基因,并建立了一套抗稻瘟病基因分析用的鑒別體系。隨后,國際水稻研究所和中國等產稻國也逐漸開展了稻瘟病抗性遺傳的系統性研究[27]。
據funRiceGenes 數據庫記載,截至2022年1月,已至少報道了143個抗稻瘟病相關基因。這些基因成簇地分布于除第3 染色體外的所有水稻染色體上,尤其是第 6、11、12 染色體上,分別多達 15、24 和 17個。Pi2、Pi9、Pi50、Pigm、Piz、Piz-t 同為 Piz 位點上的復等位基因;Pi1、Pik-h/Pi54、Pik-m、Pik-p 同為 Pik 位點上的復等位基因;Pid3 與 Pi25 等位;Pia 與 PiCO39 等位[28]。
84個基因中,有 2個隱性基因(pi21,pi55(t)),82個顯性基因;2個穗瘟抗性基因(Pi26 和 Pib1),82個葉瘟抗性基因;8個數量抗性基因 (pi21、Pi34、Pi35(t)、Pib1、Pif、Pikur1、Pikur2、Pise1),76個質量抗性基因[28]。
現已有16個位點上的24個基因被成功克隆,它們是 Pib1、Pia、Pib、Pid2、Pid3、Pik、Pik-h/Pi54、Pik-m、Pik-p、Pish、Pit、Pita、Piz-t、Pi1、Pi2、Pi5、Pi9、Pi21、Pi25、Pi36、Pi37、Pi56、Pi63、PiCO39 等[29]。
隨著抗稻瘟病基因的發現,抗稻瘟病育種進展加快,2015—2020年我國每年審定的水稻品種中,抗病等級不斷提高,抗病品種比例不斷增加,基本能達到中抗或者高抗的水平。生產上推廣應用的品種,多數品種的稻瘟病較輕。
2.2 抗蟲育種基因cry1Ab/cry1Ac
我國水稻抗蟲育種明顯落后于抗稻瘟病育種,而且重視的是轉基因抗螟蟲育種。受北美玉米轉基因抗蟲品種的啟發,華中農業大學張啟發院士領導的團隊,從1997年起就開始抗螟蟲轉基因水稻研究。華中農業大學培育的轉cry1Ab/cry1Ac 融合基因的水稻品系明恢63 及其所配雜種Bt 汕優63 在大田種植全生育期不噴施殺蟲劑的條件下,抗螟蟲效果達到95%以上[30]。1999年開始進行轉基因生物安全性評價和申報工作,并于2003年至2004年完成生產性試驗,2009年12月首次獲得原農業部頒發的生產應用安全證書,有效期為 2009年8月17日—2014年8月17日,成為我國首張轉基因水稻生產應用安全證書。2021年3月農業農村部再次續發了該轉基因水稻生產應用安全證書,轉基因抗蟲水稻育種的突破,對控制螟蟲危害、減少農藥用量、保護環境作用巨大。但品種審定之前的區域試驗尚不能進行,所以產業化之路仍非常遙遠。
3 正在應用中的基因
3.1 品質育種基因 Wx、ALK、Chalk 5、fgr 等
水稻遺傳育種技術的顯著改進,加上中國社會經濟快速發展,水稻“第四次育種突破”已初現端倪,可以說中國水稻育種或種業與移動通訊產業同步,正在進入5G 時代。
水稻5G 時代的典型特征是優質香稻:在品種特性上有優質、高產、抗病、耐熱、耐冷、矮稈、抗倒和美味;在品種類型上有常規、三系和兩系;在育種技術上有雜交育種、三系育種、兩系育種、分子技術輔助育種和基因編輯;在理論基礎上有經典和分子遺傳學。優質香稻將是5G 水稻的最典型特征。主導優質突破的基因有Wx、Chalk、ALK、fgr 等。
水稻5G 時代的首要目標是米質優和食味好,產量高成為次要目標,抗病、抗倒、耐高溫、耐低溫、矮稈成為基本條件,外觀品質好也是追求目標。米質優正在成為水稻育種的首要目標。當人們解決了吃飽問題后,自然是轉向吃好,這是社會發展的必然。水稻育種或種業必然隨之轉型。
稻米品質是決定一個水稻品種特性的重要指標之一。稻米品質主要包括5個方面:外觀品質,包括堊白度、透明度、氣味等;加工品質,包括出糙率、整精米率等;蒸煮品質,包括糊化溫度、膠稠度等;營養品質,包括蛋白質含量等;食味品質,包括直鏈淀粉含量、堿消值等。當然,食味方面還有許多亟待研究的問題。
稻米品質性狀的遺傳,現在研究公認主要與Wx(直鏈淀粉含量)、ALK(糊化溫度)、Chalk5(堊白)等基因有關。
3.1.1 Wx(直鏈淀粉含量)
Wx 基因是食味品質的主要決定因子,位于第6 染色體的短臂,該基因編碼顆粒淀粉合成酶,控制胚乳直鏈淀粉含量。研究表明,Wx 基因座至少有10個不同功能 的 等 位 基 因 , 它 們 是 Wxlv、Wxa、Wxin、Wxop、Wxb、Wxla、Wxmq、Wxmp、Wxhp 和 wx。Wxlv 野生型,源自野生稻;wx 是隱性突變基因。wx 是Wx 基因第2 外顯子第112 堿基處有23 bp 的重復,引起翻譯停止,導致顆粒淀粉合成酶缺失。其他等位基因也是由Wxlv 突變分化而來。一般認為,Wxlv 在秈稻中自然突變為Wxa,Wxlv 在秈稻中經人工誘變突變為Wxin;Wxlv 在粳稻中自然突變為Wxb,Wxb經人工誘變突變為Wxmq[31]。
3.1.2 ALK(糊化溫度)
糊化溫度是僅次于直鏈淀粉含量的稻米蒸煮品質評價的重要指標,許多文獻報道,該性狀受一主效基因控制,且在第6 染色體短臂的Wx 基因附近[32]。因此對水稻糊化溫度基因的克隆,有助于水稻品質的分子改良。中國水稻研究所的科研人員采用圖位克隆法已成功分離到水稻糊化溫度基因ALK,序列分析表明其編碼可溶性淀粉合酶Ⅱ。進一步比較不同品種間該基因的DNA 序列以及堿消法分析結果,推測出ALK 基因編碼區內的堿基替換可能引起了支鏈淀粉晶體層結構的改變,從而導致糊化溫度的變化。基因克隆得到了功能互補實驗的證實,從而為淀粉合成基因的表達調控提供了理想的材料,為淀粉合成代謝機理的闡明打下了基礎。
SSIIa/SSIIIa 的雙抑制株系呈現堊白、直鏈淀粉含量增加、糊化溫度升高、黏度減小。雙突變還降低了短(聚合度DP 5~6)和長(DP 12~23)支鏈淀粉鏈的含量、增加了中長型(DP 7~11)支鏈淀粉的含量。雙突變并不表現出加性特征,表明在淀粉合成過程中SSIIa 和SSIIIa 基因間存在相互作用。參與淀粉合成的主要酶間的成對的酵母雙雜交實驗表明,SSIIa 和SSIIIa 基因間互作可能是通過淀粉磷酸化酶發生物理作用。這些結果表明,SSIIa 和SSIIIa 在水稻籽粒淀粉合成中發揮著不同的但是又有相互重疊的作用[32]。
3.1.3 Chalk 5(堊白)
堊白是灌漿期胚乳淀粉粒和蛋白質顆粒排列疏松而充氣所形成的白色不透明部分,不僅影響了稻米可食用性產量(整精米率),同時對稻米外觀品質(透明度)、蒸煮食味和營養品質(直鏈淀粉含量、膠稠度和蛋白質含量)等也都有大的影響。因此,堊白是評價稻米品質的重要指標之一,也是水稻優質和高產的重要限制因子[31,33]。
研究表明,Chalk 5 位于第5 染色體上,起始密碼子前第721 位堿基由C 突變為T、在起始密碼子前第485 位堿基由A 突變為T,且這2個突變點均為功能突變。進一步研究表明,Chalk 5 編碼1個液泡膜質子轉運焦磷酸酶,具有無機焦磷酸水解活性和質子轉運活性,影響水稻籽粒堊白的形成和精米率等品質性狀。提高Chalk 5 表達增加胚乳堊白,可能是通過干擾發育中種子的內膜轉運系統pH 穩態來實現,這個過程影響了蛋白體的形成并且與囊泡類結構顯著增加相耦連,因此在胚乳儲存物質中形成了氣體空間,從而導致了籽粒堊白的形成[33]。
3.1.4 fgr/Badh2(香味)
香稻或香米是水稻育種家和企業的追求目標之一,也是消費者的追求。香稻或香米是指自身含有能有效被識別的特殊香味的稻谷或稻米。
稻米的香味受核基因遺傳控制,與細胞質基因無關。香味為隱性,非香味為顯性。葉片的香味受植株基因型控制,米粒的香味受胚乳基因型控制。有學者認為,稻米香味受1個隱性主效基因座位控制;也有人認為受多個基因座位上的隱性基因控制;還有人認為受主效基因和微效基因共同控制;也有人認為除主效基因外還受多個QTL 控制[34]。
水稻香味的遺傳基礎比較復雜,控制水稻香味的基因座位少則1個,多則4個以上。其原因可能有:(1)香味類型的差異,如爆米花型、茉莉花型、紫羅蘭型、山核桃香型等。(2)香味基因與多種環境因素(光、溫、肥等)相互作用。(3)香味成分的多樣性,目前檢測方法不完善。
香味Badh2 基因全長1 509 bp,包含15個外顯子和14個內含子,編碼503個氨基酸。有學者推斷,水稻中的Badh2 基因與控制水稻香味性狀的隱性基因fgr是同一個基因[34]。
國際機構從20 世紀70年代起就對香米的香氣成分進行廣泛深入的研究。目前最被廣為接受的共識是一種叫乙酰基吡咯啉(2-acetyl-1-pyrroline,2-AP)的化合物是香米的主要致香原因。盡管稻米的香味是由200多種不穩定的化合物形成,但2-AP 是這種香味形成的主要成分。而且,在其他谷類也發現了這種化合物。
3.2 粒型控制基因 GW5、qSW5、GSE5 育種
稻米的粒型是一個重要的外觀指標。粒長、粒寬、長寬比是比較復雜的數量性狀,遺傳復雜,受多個QTL控制。其中,研究人員發現了顯著影響水稻粒寬和粒質量兩個性狀連鎖的主效基因GW5、qSW5、GSE5 等。
GW5 精細定位于含有1 212 bp 缺失的21 kb 基因組區域,在1 212 bp 缺失下游約5 kb 的區域,有一個編碼鈣調素結合蛋白的基因,即GW5。存在于寬粒品種的1 212 bp 缺失通過調控GW5 的表達量進而調控籽粒大小。利用CRISPR 技術將GW5 基因敲除,可以增加其他不含1 212 bp 缺失的水稻品種籽粒的粒寬和粒質量,達到增產效果[35]。
利用粳稻品種日本晴和秈稻品種Kasalath 雜交構建的F2群體,在第5 染色體定位到一個控制粒寬的主效QTL,命名為qSW5。通過圖位克隆將qSW5 精細定位在2 263 bp 內,最后通過基因表達分析,以及互補實驗確定其中一個ORF(開放閱讀框)為qSW5[18]。GSE5編碼一個鈣調素結合蛋白,可以與水稻鈣調素蛋白OsCaM1-1 在體內相互作用。GSE5 啟動子區域的缺失是導致水稻品種粒寬變異的遺傳基礎。細胞學分析表明,GSE5 通過影響種子的細胞數目來控制籽粒寬度和粒型[36]。
3.3 秈粳交雜種優勢利用基因S5-n
秈粳交雜交水稻現在已經有了多方面的突破。一是秈粳交雜交稻新品種選育快速發展,至2020年僅甬優系列品種就多達40個,還有春優系列、浙優系列、中嘉優6 號和中嘉優9 號等。二是農業農村部在確認超級稻品種和國家水稻數據中心在劃分品種類型時,均把秈粳交雜交稻作為一個新類型,甬優12、甬優15、浙優 18、春優 84、甬優 538、甬優 2640、甬優 1540、甬優7850 等8個秈粳交雜交稻品種先后被確定為超級稻品種,在2021年有效的135個超級稻中占有不小的比例。三是“秈不粳恢”模式取得較大進展,如湖北省農業科學院選育的“秈不粳恢”雜交稻品種旱優79 已通過審定。
廣親和材料的發掘以及秈粳雜種不育基因的克隆和應用是有效解決秈粳雜種不育的根本途徑。秈粳雜交品種的育性主要受控于水稻第6 染色體的S5 位點,由 3個緊密連鎖的基因 ORF3、ORF4 和 ORF5 組成,形成“殺手-保護者”系統。在雌孢子形成過程中,“殺手”ORF5+和“搭檔”ORF4+的行為會引起內質網脅迫,“保護者”ORF3+能阻止內質網發生脅迫反應,促進細胞產生正常的配子,但是ORF3-不具這一功能,結果導致提前發生程序性細胞死亡與胚囊敗育。典型的秈稻攜帶ORF3+ORF4-ORF5+組合,而典型的粳稻攜帶ORF3-ORF4+ORF5-組合;在秈粳雜種中,ORF5+和ORF4+合作殺死雌配子,而攜帶護衛基因ORF3+的雌配子免于敗育[37]。
S5 的三等位基因系統是水稻秈粳雜種生殖隔離和雜交親和性的主要調節因子。日本學者池橋等(1984)根據不同生態型水稻品種間的雜交研究,提出了廣親和基因的假說,認為秈、粳和廣親和品種之間的雜種一代育性受控于水稻第6 染色體位點一組復等位基因的相互作用。廣親和基因可有效地克服秈粳之間雜種一代的不育現象。
S5-j 位于第 6 染色體,cDNA 全長 2 495 bp,包含3個外顯子,編碼一個由472個氨基酸組成的蛋白產物。產物包含信號肽、中央結構域、N 端小葉和C 端小葉。粳稻品種日本晴和Balilla 完全一樣,第273 位為亮氨酸,第471 位為頡氨酸;秈稻品種南京11 第273 位為苯丙氨酸,第471 位為丙氨酸,終止密碼子下游172 bp 處缺失1個A;廣親和品種02428 轉錄起始位點ATG 前 67 bp 以及 ATG 后 69 bp 總共 136 bp 缺失,造成包含信號肽的N 端115個氨基酸缺失,導致不能定位到細胞壁上[37]。
綜上所述,秈粳雜交稻育種成功的必要條件是克服亞種間雜交不育,其有效途徑是廣親和基因S5-n 的利用。在秈粳交育種時代,借鑒“玉米雜種優勢群”的理論,嚴格建立父母本各自的雜種優勢群,親本育種在群內進行,配組在群間進行,避免遺傳血緣混雜。
4 未來可能被利用的基因
4.1 抗蟲育種
4.1.1 抗飛虱育種基因Bph
水稻抗病育種特別是抗稻瘟病育種取得明顯突破之后,抗蟲水稻育種就在期待中。水稻害蟲主要有兩類,一類是吃葉或吃莖的螟蟲,一類是吸汁的針吸式害蟲,如蚜蟲、薊馬、飛虱等。抗螟蟲育種,曾借助轉基因技術獲得了突破,由于學術界對轉基因風險爭論的原因,尚未能商業化應用。
褐飛虱是危害水稻生產的重要害蟲,利用寄主抗性培育抗褐飛虱品種被認為是防治其危害的有效途徑。自20 世紀70年代起,各國相繼開展了水稻褐飛虱抗性基因的發掘工作,到目前為止,已經鑒定和報道了至少34個抗褐飛虱基因位點,其中顯性基因19個、隱性基因15個,已經定位的主效抗褐飛虱基因達43個,有9個位點的抗性基因被克隆,分別是Bph14、Bph15、Bph3、Bph2/26、Bph9/1/7/10/21、Bph18、Bph29、Bph32、Bph6。此外還鑒定了一些重要的抗性QTL[38]。
4.1.2 抗螟蟲育種
螟蟲是水稻的大敵。轉抗螟蟲基因水稻的抗蟲效果已被實踐廣為證實,轉抗蟲基因玉米、棉花、大豆等在全球已有廣泛應用,也是一種佐證。對于我國而言,水稻產量穩定始終是糧食安全的重要組成部分,當稻米產不足需、螟蟲危害風險加重的情況出現時,中國轉抗螟蟲基因水稻的商業化應用將有可能。
4.2 抗除草劑育種
在水稻生產中,草害是影響高產、穩產、優質的重要因素之一。隨著直播栽培面積的不斷擴大,草害問題也越來越嚴重。據統計,由于稻田雜草的侵害,水稻產量的損失可達40%以上。近年來,除草劑在農業生產上得到大面積推廣應用,有效提高了我國水稻的產量,大大減輕了稻作勞動量,降低了水稻生產成本。
除草劑通過抑制雜草代謝過程中的酶系統,使其代謝過程受阻,達到除草的目的。根據此原理,通過物理化學誘變或基因工程的方法對水稻體內靶酶進行修飾,使其對除草劑不敏感,或通過基因工程的方法轉入與除草劑有高親和力的酶,降低除草劑對水稻內源靶酶的影響,減少除草劑對水稻內部代謝作用的干擾,達到水稻具有抗除草劑特性的目的。
通過誘變技術獲得抗除草劑水稻在美國早已應用。例如,美國Rice Tec 推出了Clearfield XL729、Clearfield XL745、Clearfield XP756、Clearfield Xp534 等抗咪草煙除草劑雜交水稻,市場份額高達75%。
近年來,我國不少學者對抗除草劑水稻種質資源進行了廣泛研究創制,并取得了許多成果。我國最早通過審定的抗除草劑水稻品種是天津市水稻研究所培育的金粳818。北京大學鄧興旺團隊用EMS 誘導黃華占獲得的抗除草劑突變體潔田稻001,2019年獲得植物新品種權授權(授權號:CNA20150959.0),2021年通過安徽省審定(審定編號:皖審稻20212004)。在2020年,荃優潔田 1 號(荃 9331A×潔田稻 001,國審稻20206078)、興農絲占(粵農絲苗/潔田稻 001,國審稻20206104)、仲旺絲苗(五山絲苗/潔田稻 001,國審稻20206105)已通過國家審定。這預示著潔田稻系列品種可以在市場上應用,成為中國抗除草劑水稻的一個開端。表4 是人工誘變和基因編輯獲得的一些突變體,其中黃華占EMS 突變體已經通過審定,命名為潔田稻001,進入了商業化應用。經過20 余年的研究,已知EPSPS、aroA、pat、Bar、protox、P450、bxn、csrl 等基因,通過轉基因技術可以獲得抗特定除草劑的轉基因水稻。

表4 人工誘變和基因編輯獲得的抗除草劑水稻突變體
鑒于水稻草害的嚴重性和水稻輕簡化(免耕、直播、機拋)栽培條件下化學除草的必要性,抗除草劑水稻育種商業化應用已經迫在眉睫,諸多基因通過EMS誘變而非轉基因途徑實現,規避了轉基因安全的學術爭論,即將引起又一次水稻育種突破。
4.3 多抗性育種技術
隨著商業化育種體系的完善,抗倒、耐熱、耐冷、耐旱、抗病(稻瘟病以外)等多種抗性育種需求,都會通過育種效率的提高而完成,只不過這種突破不一定能夠引起業內直觀上的感知而已。
5 未來育種技術新方向——基因編輯
育種就是聚合控制目標性狀的基因座位上能產生最佳表現型的有利基因型或有利基因型的集合。因而育種的主要工作就是創造變異、固定或穩定有利變異、選擇有利變異。基因編輯技術就是一種創造或創新變異的技術。
基因編輯依賴于經過基因工程改造的核酸酶(分子剪刀),在基因組中特定位置產生位點特異性雙鏈斷裂,誘導生物體通過非同源末端連接或同源重組來修復特異性雙鏈斷裂,因為這個修復過程容易出錯,從而導致靶向突變,這種靶向突變就是基因編輯。基因編輯以其能夠高效率地進行定點基因組編輯,被作物育種界寄予厚望。很顯然,基因編輯的前提是知道要編輯什么基因。因此,要編輯的基因都是主效基因,所不同的是,基因編輯的基因一般是育種不需要的非有利基因,期望它產生有利突變。迄今為止,分子遺傳學家對水稻基因編輯的主效基因列于表5。

表5 編輯過的水稻主效基因(截至2021年)
可以預見,隨著技術突破,越來越多的水稻基因將被編輯。不過,至今為止,還沒有一個基因編輯的水稻品種通過審定。
6 討論與展望
無論是玉米和水稻的萬年自然進化,還是數千年人類參與的人工馴化,或者是人工直接干預的百余年水稻育種,我們都可以清楚看出,水稻從野生稻(Oryza rufipogon)進化為栽培稻(Oryza sativa),變化或馴化的主要性狀包括株型、落粒性、芒的長短和有無、異交率、產量、休眠性等,主效基因的改變主導了水稻的進化、馴化和育種突破,其他的基因只是起輔助作用。
回顧百余年的水稻育種,半矮稈基因sd1、野敗細胞質雄性不育基因WA352 及恢復基因Rf3 和Rf4、光溫敏核不育基因pms3 或溫敏核不育基因tms5 分別導致了中國水稻育種的三次跨越式突破——矮稈水稻、三系法雜交水稻和兩系法雜交水稻。抗稻瘟病基因Pi也可以說引起了中國水稻育種又一次較大突破,只是時間跨度長、抗病基因多、效果沒有前三次明顯。現在回過頭來看,抗稻瘟病品種的普及,對水稻生產的貢獻不亞于前面的任何一次跨越式突破,殺菌劑用量減少、穩產性增加、產量損失降低甚至提高,抗稻瘟病育種功不可沒。
中國的水稻育種,當前正在發生的似乎有兩場變革,一是追求品質的優質育種,包括香稻育種以及實際意義存疑的追求長粒的粒型育種,都可看作優質育種的一部分;二是在保持優質的前提下追求超高產的秈粳交雜種優勢利用育種。
可以預見,在不遠的將來,“豐(產)優(質)抗(病)、長(粒)香(米)美(味)”將成為水稻育種的主攻方向。水稻的進化從1G 到5G,不難發現栽培生產上所要求水稻的主要農藝性狀的綜合進化,所圍繞的也是若干主效基因的聚合。
作者據此推論,未來水稻育種的跨越式突破,無疑仍將由關鍵主效基因的變化所驅動。最可能發生的水稻育種革命大概率將由抗除草劑、抗飛虱、抗螟蟲、耐熱、耐冷、抗倒、抗其他病害等主效功能基因所推動。本文闡述和預測的推進水稻育種突破的主效基因歸納于表6。

表6 已導致和可能導致水稻育種革命的主效基因
除此之外,水稻育種技術也一直在呼喚變革,基因編輯正在被廣泛探索。一般認為,基因編輯仍然是建立在已知的主效基因之上,所不同是,它主要是基于對眾多不利等位基因的研究。本文的論述表明,水稻育種一直是在主效基因上做文章。一旦新的主效基因被發掘,該基因所控制的相關農藝性狀改良就會突破。這告誡我們,水稻育種創新,就是抓住新的主效基因,加以攻克,取得突破。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
