Foxo6在高糖高脂應激誘導血管內皮細胞胰島素抵抗中的表達及與NF-κB通路的相互作用
2022-04-07 02:48:08涂以思熊小偉牛艾琳尹婷婷張雅楠黃起壬
中國藥理學通報 2022年4期
關鍵詞:檢測
夏 敏, 涂以思, 熊小偉, 牛艾琳, 尹婷婷, 張雅楠, 黃起壬
(1. 江西省基礎藥理學重點實驗室,江西 南昌 330006;南昌大學 2.藥學院藥理教研室、3.第三臨床醫學院,江西 南昌 330006)
近年來,在世界大范圍的慢性病的排名中,糖尿病的死亡率已升到第二,患者常由于心血管并發癥而死亡,給社會、家庭造成沉重負擔[1-2]。糖尿病有1型和2型之分,因為胰島β細胞天生胰島素分泌不足引起了1型糖尿病,而2型糖尿病(type 2 diabetes mellitus,T2DM)以慢性高血糖和胰島素抵抗(insulin resistance,IR)為基本特點[3],其發病率呈年輕化趨勢[4],大多數研究認為IR先于T2DM的發生,因此對于IR的研究尤為重要。
叉頭框(forkhead box,Fox)超級家族是繼homeodomain、bHLH和bZIP這3個家族之后的第四大轉錄因子超級家族,叉頭框o(forkhead box o,Foxo)是Fox超級家族的成員,其結構特征是存在叉頭域,這在Fox家族的所有成員中都普遍存在[5]。其中,哺乳動物中的o家族(叉頭框/盒)由Foxo1(FKHR)、Foxo3(FKHRL1)、Foxo4(AFX)和Foxo6組成。Foxos在多種不同的細胞中發揮著重要作用,不僅參與了眾多細胞的生理過程,而且與人類的多種疾病包括癌癥、糖尿病、不孕癥、神經退行性病變和免疫系統功能障礙等有關[6]。例如:肝臟中,胰島素通過抑制Foxos使葡萄糖的產生減少,同時使葡萄糖的利用率增加,而肝臟IR會激活Foxos,引起高血糖、高甘油三酯血癥,并且,Foxos可以誘導肝組織產生IL-1β等炎性細胞因子,因此其在促進肝臟IR和炎癥中發揮著重要作用[7]。據報道Foxos家族成員通過抑制PI3K/AKT信號下游通路與IR有關。但目前關于Foxos與內皮細胞IR的聯系尚未見報道,各亞型在HG/HF應激誘導內皮細胞IR中的表達如何以及具體哪一個亞型發揮了關鍵作用也不清楚,因此本文探討Foxos在HG/HF應激誘導內皮細胞IR中的表達改變并研究其具體的作用機制。
血管內皮細胞損傷、IR和炎癥標志物的增加是T2DM發生和發展的核心因素[8],但目前關于血管內皮細胞IR發生的分子機制還不十分明確。因此,闡明其發生的分子機制對于發現治療T2DM藥物的作用靶點,改善患者的生活質量具有重要意義。慢性炎癥反應在IR的發病機制中扮演著重要作用。NF-κB是典型的炎癥信號通路之一,參與了肥胖,且和糖尿病發病有關。其信號通路的激活不僅可以實現對下游基因的調控,而且與炎癥密切相關,并且可以從多種途徑誘發IR,在IR的形成中起著重要作用[9]。NF -κBp65或p65(也稱為RELA),是核轉錄因子,它可以增強巨噬細胞促炎細胞因子的產生,加重高脂飲食誘導的慢性炎癥和巨噬細胞IR[10];而抑制RELA核轉運蛋白的表達或抑制RELA核轉運可增加胰島素刺激的葡萄糖轉運和消耗,從而改善巨噬細胞IR。那HG/HF誘導的內皮細胞IR是否可以通過炎癥這條通路發揮作用,以及沉默Foxo6基因后是否可以通過此通路逆轉IR?以上就是我們所關注的問題。
1 材料
1.1 細胞株及血清培養基實驗用人臍靜脈內皮細胞(human umbilical vein endothelial cells,HUVECs)株購自美國ATCC( Catalog No: CRL1730) ;胎牛血清購自天津TBD公司;DMEM培養基購自北京Solarbio公司。
1.2 抗體和試劑NO檢測試劑盒(20190722)、人內皮素-1(ET-1)酶聯免疫檢測試劑盒(2H-KMLJh311095)均購自南京建成;Foxo6兔多克隆抗體(19122-1-AP)、NF-kBp65兔多克隆抗體(96033)、GAPDH鼠多克隆抗體(Ls203561)均購自武漢三鷹;AKT-pS473兔單克隆抗體(#9271S)、AKT兔單克隆抗體(#9272S)均購自Abcam;逆轉錄試劑盒(#22106-01)購自Tolobio;SYBR Green(FP171206)購自天根;熒光素酶報告基因檢測試劑盒購自普洛麥格,腫瘤壞死因子α(tumor necrosis factor,TNF-α)ELISA試劑盒(2H-KMLJh311776)購自武漢博士德;白細胞介素-6(interleukin-6,IL-6)試劑盒(2H-KMLJ31201mm)購自南京卡米洛;ChIP試劑盒(#25268)購自Cell Signaling Technology;引物購自南昌臻善生物科技公司,小干擾RNA(small interfering RNA,siRNA)購自銳博生物有限公司,過表達質粒RELA(49607-11)購自上海吉凱基因有限公司。
1.3 主要儀器680型酶標儀(BIO-RAD,美國);SmartSpec Plus核酸蛋白分析儀(BIO-RAD,美國);PCR儀(BIO-RAD,美國);熒光倒置顯微鏡(Olympus,日本);流式細胞儀(BECKMAN COULTER,美國)。
2 方法
2.1Foxos在HG/HF處理的HUVECs中的差異性表達將HUVECs均勻接種于培養皿,隨機分為兩組,即Ctrl組和高糖高脂(High glucose and High fat,HG/HF)組,HG/HF組用22 mmol的葡萄糖和0.25 mmol的棕櫚酸(PA)處理細胞24 h后,兩組均用胰島素(100 nmol·L-1)處理30 min,去除細胞上清液,采用Trizol法提取各組細胞RNA,按照Tolobio試劑盒將RNA逆轉錄成cDNA,通過qRT-PCR檢測Foxo1、Foxo3、Foxo4、Foxo6基因的表達,PCR擴增所用引物由南昌臻善生物科技公司合成,Foxo1上游引物:5′-TTGCTGACTTCTGACTCTCCTC-3′,下游引物:5′-ATGTCCAGCGTGGGTATGG-3′;Foxo3上游引物:5′-GAACTCCCTACGCCAGTCTC-3′,下游引物:5′-GCAGCAAAGGACATCATCG-3′;Foxo4上游引物:5′-GCTCCGACTCTTCTGTTGCT-3′,下游引物:5′-AGGCATTCTGTCTTGGCTTG-3′;Foxo6上游引物:5′-GCGGAAAACTCACACCTACC-3′,下游引物:5′-CGGTTGAAGAGAGGGACTGA-3′。PCR擴增體系為20 μL,其中cDNA為2 μL,上下游引物各0.8 μL,PCR循環數為39,95 ℃預變性15 min,95 ℃變性10 s,60 ℃退火延伸32 s。
2.2Foxo6 siRNAs沉默效率的篩選實驗將HUVECs均勻接種于六孔板,隨機分為5組,即control組、control+NC組、control+Foxo6siRNA1組、control+Foxo6siRNA2組、control+Foxo6siRNA3組,除Ctrl組外,其余組分別轉染陰性對照NC、Foxo6siRNA1、Foxo6siRNA2、Foxo6siRNA3 24 h后,采用Trizol法提取各組細胞RNA,按照Tolobio試劑盒將各組RNA逆轉錄成cDNA,通過qRT-PCR進行檢測。也可以用裂解液裂解各組細胞,Western blot法檢測各組細胞的Foxo6蛋白表達,Foxo6的分子量為58,從而篩選出沉默Foxo6最顯著的siRNA。
2.3Foxo6 siRNA逆轉IR實驗將HUVECs均勻接種于培養皿,隨機分為4組,即control組、HG/HF組、HG/HF+NC組、HG/HF+Foxo6siRNA3組,除control組外,其余組分別加HG/HF處理細胞24 h,然后HG/HF+NC組轉染陰性對照,HG/HF+Foxo6siRNA3組轉染siRNA3 24 h,收集細胞上清液檢測NO和ET-1,收集細胞通過Western blot檢測AKT-pS473的表達。
將SD雄性大鼠用10%水合氯醛進行麻醉,沿大鼠的胸部和腹部縱向切開,剝離大鼠的主動脈,用PBS洗干凈,將其移入培養瓶中,放入孵箱孵育3~5 h,然后加入完全培養基(含10%FBS、肝素鈉和生長因子ECGS),待其生長狀態良好,傳代置培養皿。將原代培養的大鼠主動脈內皮細胞(RAECs)隨機分為4組,即control組、HG/HF組、HG/HF+NC組、HG/HF+Foxo6siRNA3組,收集細胞上清液檢測NO、ET-1,收集細胞通過Western blot檢測AKT-pS473的表達。
2.4 免疫共沉淀法(CoIP)檢測Foxo6與NF-κBp65相互作用將HUVECs均勻接種于培養皿,按照總蛋白提取方法提取4組處理好的細胞蛋白,每個組又分為加入相應抗體的IP組、陰性對照IgG組、未經任何處理的input組,除input組外,IP組和IgG組分別加入1 μL NF-κBp65的抗體混懸過夜;次日IP組和IgG組再分別加入25 μL的瓊脂糖珠使蛋白沉淀,4 ℃混懸儀混勻1~3 h或者過夜;4 ℃ 2 500 r·min-1離心5 min,留沉淀,加入上樣緩沖液,上樣緩沖液的量根據加入珠子的量決定,渦旋混勻,煮沸10 min,最后加入適量上樣緩沖液混勻,-20 ℃保存備用,通過Western blot檢測Foxo6的蛋白表達。
2.5 免疫熒光檢測NF-κBp65亞細胞定位將細胞均勻接種于24孔板中,經前期處理后,加4%多聚甲醛固定細胞,隨后加入TritonX-100室溫通透3 min,洗凈后加入5%BSA室溫封閉2 h,然后加入由5%BSA新鮮配制的NF-κBp65的一抗(比例為1:100)4 ℃孵育過夜,次日加熒光二抗(1:100)室溫孵育2 h,然后DAPI染核,最后熒光顯微鏡下觀察。
2.6 染色質免疫共沉淀(ChIP)檢測NF-κBp65與其靶基因啟動子結合活性
2.6.1細胞交聯與樣品制備 根據實驗要求及分組,將細胞培養于培養皿,加入37%的甲醛溶液固定細胞,加入10×甘氨酸溶液終止上述固定反應,加入含PIC的1X PBS刮下細胞,收集細胞離心,去除上清。
2.6.2細胞核處理和染色質剪切 分別將離心完的細胞用1 mL 1×緩沖液A+DTT+PIC重懸,冰上靜置,3 000 r·min-1,離心5 min沉淀細胞核。將上清棄去,用緩沖液B+DTT 1 mL重懸細胞核沉淀,離心去除上清。用緩沖液B+DTT 100 μL繼續重懸,加微球菌核酸酶,消化DNA為150~900 bp的片段;加0.5 mol·L-1 EDTA使消化停止,離心收集細胞核沉淀,將其重懸于1×ChIP緩沖液+PIC中,冰上孵育,對細胞核樣品進行超聲破碎,離心,轉移上清,每管取2 μL測定染色質濃度,剩余樣品保存于-80 ℃進行后續實驗。
2.6.3染色質免疫沉淀 每管加等量的交聯染色質樣品,每個沉淀反應需要用1×ChIP緩沖液補齊到500 μL,并補加適量的200×PIC,將稀釋后的ChIP染色質,每個樣品吸取10 μL作為Input組,保存于-20 ℃;每個樣品管加3 μL的Foxo6抗體,每個陰性對照管加1.5 μL的IgG抗體,每個陽性對照管加10 μL H3抗體,封嚴各反應管,轉子上孵育4 h以上或過夜。取30 μL ChIP級蛋白G瓊脂糖微珠加入到每個免疫沉淀反應中,轉子上孵育2 h,沉淀蛋白G瓊脂糖微球,除上清;分別加低鹽漂洗液和高鹽漂洗液洗滌,吸除上清。
2.6.4將染色體從抗體/蛋白G微珠上洗脫并解交聯 每組對應的Input組分別加入150 μL 1×ChIP洗脫緩沖液,室溫放置;其它每份ChIP免疫沉淀樣品中分別加入150 μL 1×ChIP洗脫緩沖液,進行洗脫;沉淀蛋白G瓊脂糖微球,除上清;把每個樣品管中洗脫下來的染色質上清轉移到一個新的離心管中,并做好標記。在所有管中,包括Input組管,都加入6 μL 5 mol·L-1 NaCl和2 μL蛋白酶K,65 ℃孵育2 h。
2.6.5離心柱純化DNA 將750 μL DNA結合緩沖液加到DNA樣品中混勻,將每組樣品450 μL轉移到離心柱中,14 000 r·min-1離心,棄廢液,加漂洗緩沖液,離心,加50 μL DNA洗脫緩沖液,離心。
2.6.6用PCR定量進行ChIP富集效率的分析 瓊脂糖凝膠電泳對IL-6和TNF-α進行定性分析,進行ChIP富集效率的PCR定量分析。
2.7 熒光素酶報告基因檢測NF-κB轉錄活性構建編碼RELA啟動子的熒光素酶報告基因質粒,用0.5 g空載體或RELA質粒分別轉染control組和HG/HF組的HUVECs細胞于24孔板,實驗分為4組,即control+NC組、control+RELA組、HG/HF+NC組、HG/HF+RELA組,根據24孔板的用量,按照說明書操作,根據公式:螢火蟲/海腎熒光素酶進行計算,并以control+NC組的比值為單位1,既可以得到不同處理組的相對熒光素酶活性。
2.8 ELISA法檢測NF-κBp65靶基因的表達按照ELISA檢測試劑盒說明書對IL-6、TNF-α進行檢測,繪制標準曲線并計算各組的含量。
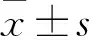
3 結果
3.1 IR模型的確立HUVECs IR指標常用培養基上清NO、ET-1水平以及Western blot法中AKT-pS473的表達水平表示,HG/HF組與control組比較,NO水平降低(P<0.05),而ET-1水平升高(P<0.05),AKT-pS473的表達水平明顯降低(P<0.05),表明IR模型的成功建立(Fig 1)。
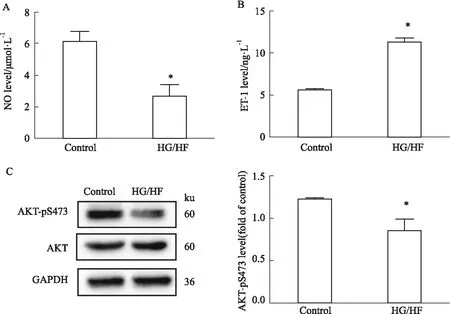
Fig 1 NO, ET-1 and AKT-pS473 levels of various treatment in cultured HUVECs
3.2Foxos在HG/HF應激誘導IR中的表達變化為了驗證Foxos在HG/HF誘導的內皮細胞IR中是否發生了變化,采用qRT-PCR和Western blot檢測其表達水平,qRT-PCR的結果顯示Foxo6在HG/HF組中變化最明顯,并且表達量明顯升高(P<0.01),Western blot的結果表明Foxo6在HG/HF組中的表達量也是明顯升高的(P<0.05)(Fig 2)。
3.3 沉默Foxo6 siRNA的篩選3.2中結果證明Foxo6在HG/HF組中變化最明顯并且表達量升高,說明Foxo6在HG/HF應激誘導IR中可能發揮了重要作用。為了進一步闡明Foxo6與IR間究竟是一種伴隨現象還是一種因果關系,通過構建Foxo6 siRNA,觀察Foxo6水平下調是否能逆轉HG/HF應激所誘導的IR效應。為此,針對Foxo6基因設計了3個不同靶位的siRNA序列并構建成質粒,目的是篩選出沉默效率最高的siRNA,用于后續的研究。qRT-PCR和Western blot均顯示,Foxo6 siRNA3序列沉默效果最明顯(P<0.05),因此選取Foxo6 siRNA3序列用來進行后續研究(Fig 3)。
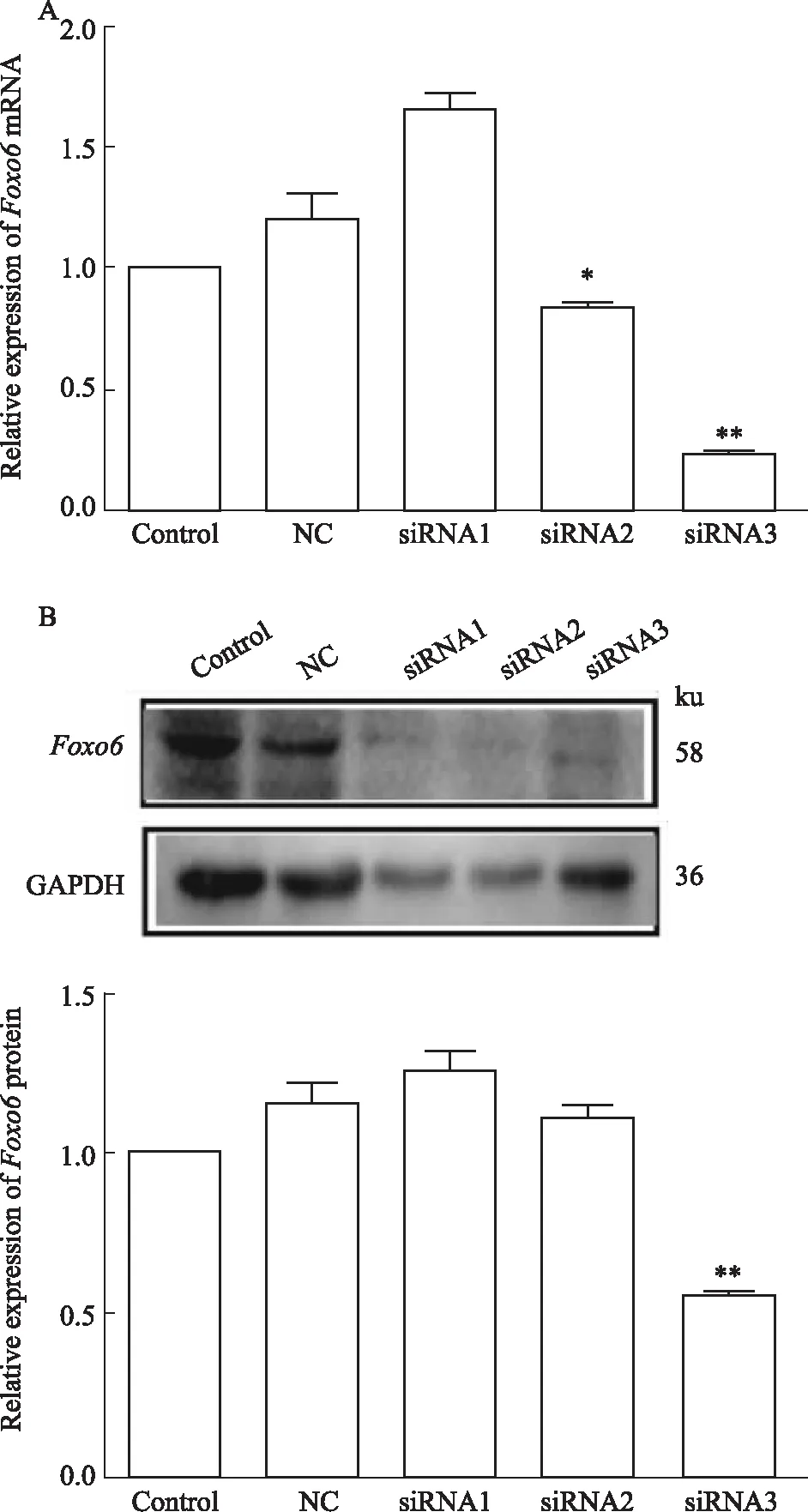
Fig 3 Foxo6 mRNA and protein expression levels of various treatment in cultured HUVECs n=3)
3.4 沉默Foxo6對HG/HF應激誘導IR的影響實驗分為control、HG/HF、HG/HF+NC、HG/HF+Foxo6siRNA3組。在HUVECs實驗中,與HG/HF組相比,沉默Foxo6后,NO水平明顯升高(P<0.01),ET-1水平降低(P<0.05),AKT-pS473的表達水平明顯升高(P<0.05),說明沉默Foxo6可明顯逆轉HG/HF誘導的血管內皮細胞IR(A-C);在大鼠原代主動脈內皮細胞(RAECs)實驗中也得到了類似的結果(D-F)(Fig 4)。
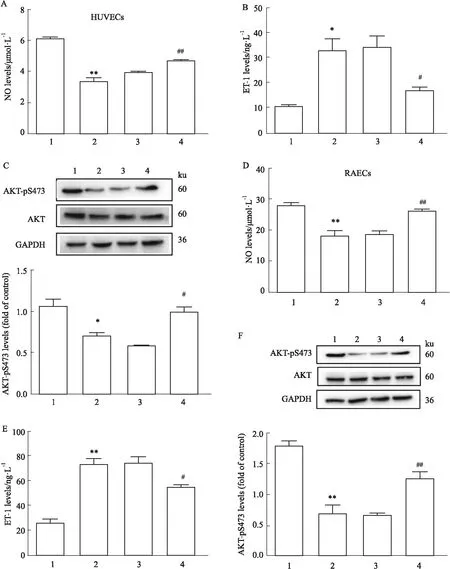
Fig 4 NO, ET-1 and AKT-pS473 levels of various treatment in cultured HUVECs and n=3)
3.5Foxo6沉默對HG/HF誘導的Foxo6與NF-κBp65相互作用的影響研究表明,IR也是一種慢性炎癥反應。NF-κB是炎癥反應中最為關鍵的轉錄因子。同為核轉錄因子的Foxo6是否與NF-κB通路產生相互作用從而調控HG/HF誘導的IR。為此,探究了Foxo6對NF-κB介導的炎癥通路的影響。CoIP結果顯示,與control組相比,HG/HF組和HG/HF+NC組中Foxo6的表達水平明顯升高(P<0.01),說明在HG/HF應激下Foxo6與NF-κBp65發生了相互作用促進IR;轉染Foxo6 siRNA3之后,Foxo6的表達水平降低(P<0.05),表明沉默Foxo6后使其相互作用減弱(Fig 5)。

Fig 5 Interaction between Foxo6 and NF-κBp65 of various treatment in cultured HUVECs n=3)
3.6Foxo6沉默對HG/HF誘導的NF-κBp65亞細胞定位的影響免疫熒光結果顯示,control組NF-κBp65主要位于胞質中,而HG/HF組NF-κBp65定位在核內,說明NF-κBp65在受到HG/HF應激后發生了核轉移;沉默Foxo6基因后,NF-κBp65主要定位于核外,揭示沉默Foxo6基因可抑制NF-κBp65的核移位(A);此外,通過提取細胞核蛋白和細胞質蛋白,分別進行Western blot檢測,其結果與免疫熒光結果相似(B、C)(Fig 6)。
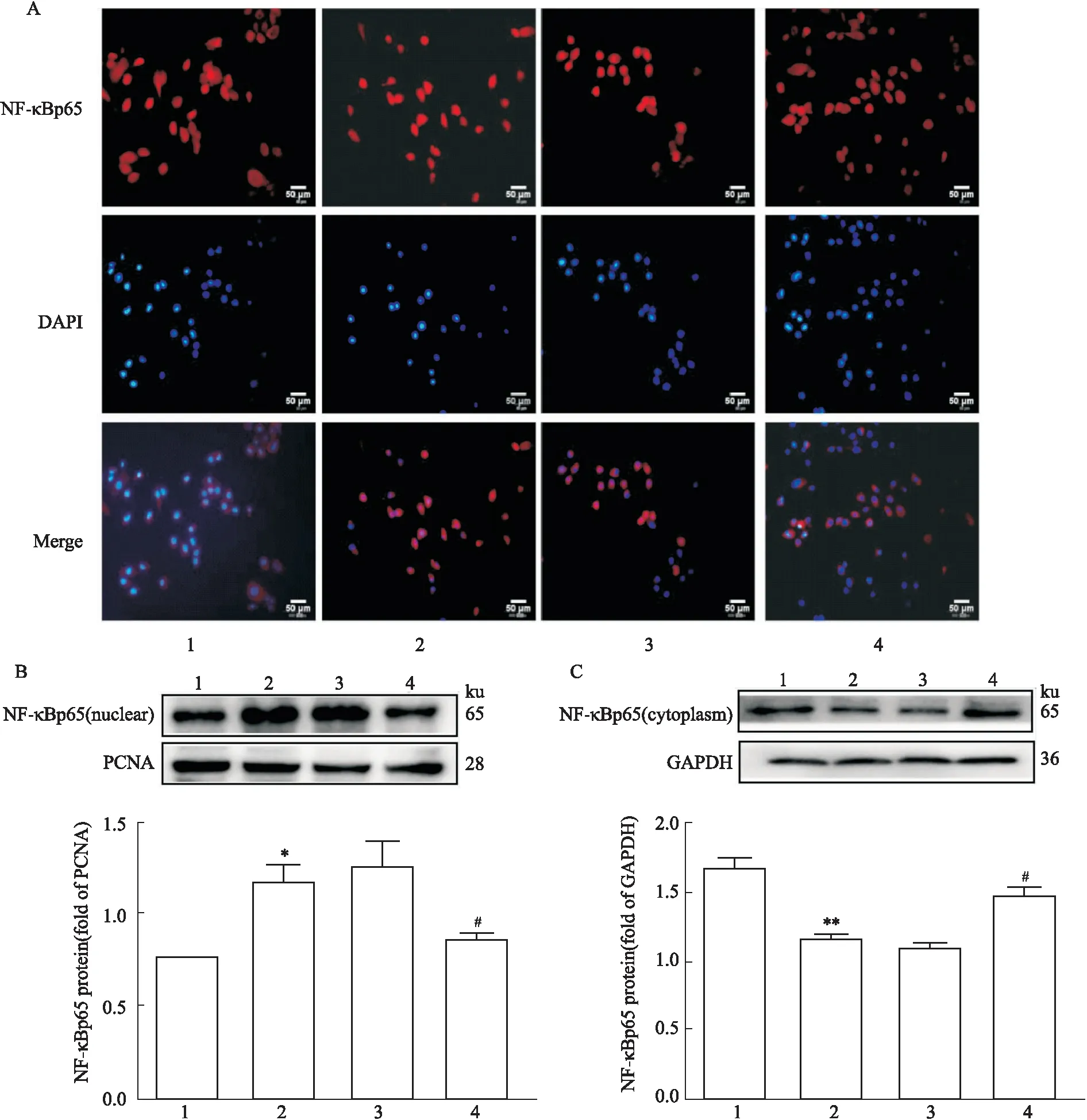
Fig 6 Expression and location of Foxo6 in nucleus and cytoplasm after Foxo6 was silenced in various treatment in cultured HUVECs n=3)
3.7Foxo6增強NF-κBp65與靶基因啟動子的結合活性IL-6、TNF-α是NF-κB的下游靶標分子,瓊脂糖凝膠電泳結果表明HG/HF和HG/HF+NC組中IL-6和TNF-α基因表達增加(P<0.01,P<0.05),沉默Foxo6基因后IL-6和TNF-α基因表達降低(P<0.05),qRT-PCR結果表明Foxo6增強NF-κBp65與IL-6和TNF-α的富集效率(Fig 7)。
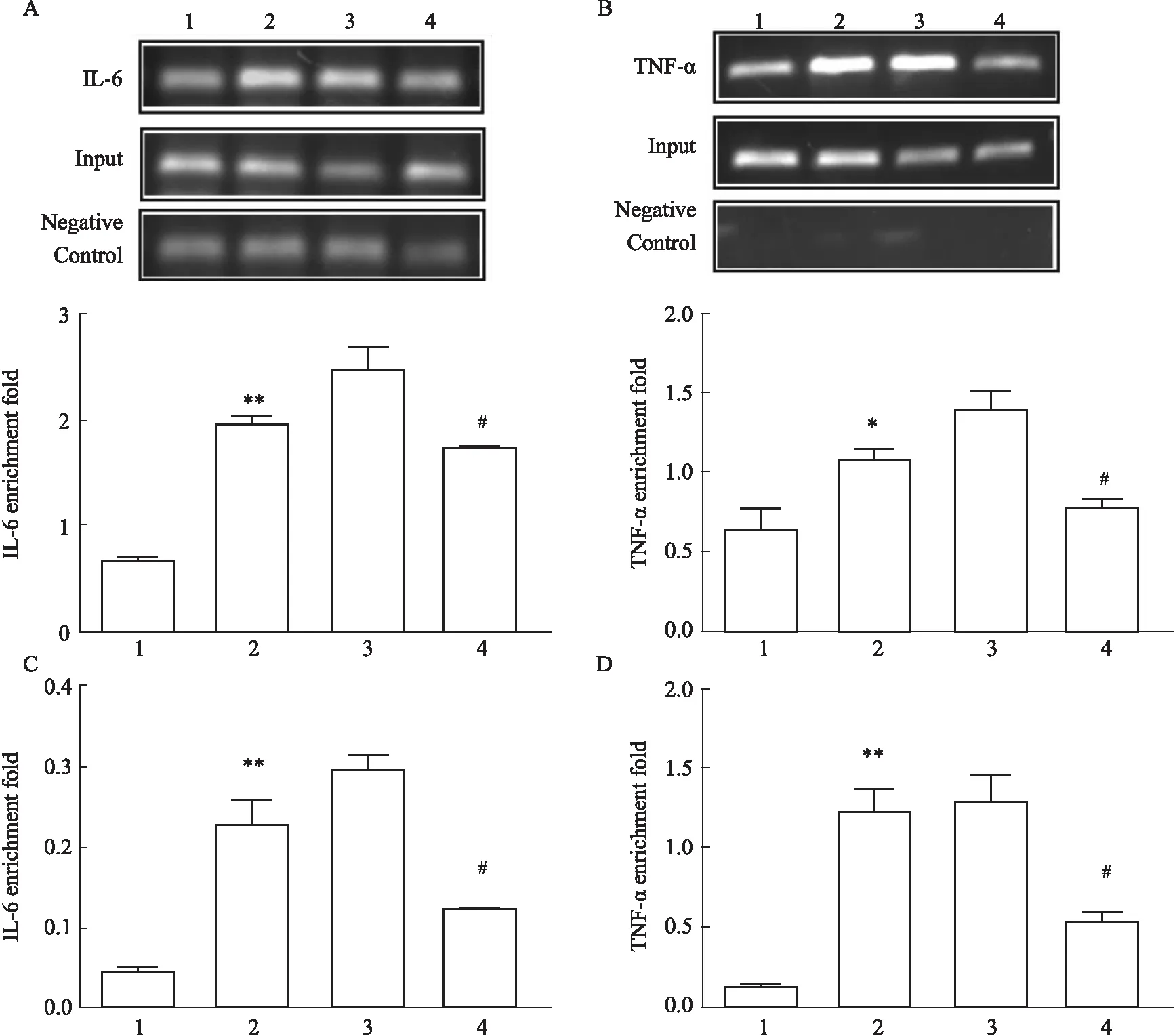
Fig 7 Silencing Foxo6 inhibited binding activity of NF-κBp65 to its target gene promoter in various treatment in cultured HUVECs n=3)
3.8Foxo6增強NF-κB轉錄活性以及促進NF-κBp65靶基因的表達實驗分為4組,分別是control+NC組,control+RELA過表達質粒組,HG/HF+NC組、HG/HF+RELA過表達質粒組。熒光素酶報告基因結果顯示,與control+RELA或HG/HF+NC組相比,HG/HF+RELA組中的NF-κBp65的轉錄活性明顯增強(P<0.01),表明Foxo6影響NF-κBp65的轉錄活性(A);IL-6、TNF-α是NF-κB的下游靶標分子,炎癥狀態下IL-6和TNF-α的表達活性增強。ELISA結果顯示,與Ctrl組相比,HG/HF組中IL-6和TNF-α的表達增加(P<0.01),表明Foxo6促進NF-κBp65下游靶基因的表達;沉默Foxo6后IL-6和TNF-α的表達降低(P<0.05,P<0.01),表明Foxo6沉默后可能下調IL-6和TNF-α的表達(B、C)(Fig 8)。
Foxos是一類轉錄因子家族,重要作用包括其可以激活或抑制基因轉錄、調節下游靶基因的表達,異常的Foxos功能與神經退行性疾病、癌癥和糖尿病[14]等疾病有關,研究表明Foxos與IR有關[15],但其在內皮細胞IR中的作用如何以及具體哪一個亞型起了關鍵作用尚不明確,而且Foxos在不同的細胞和不同的狀態下發揮的作用也各不相同。先前的研究認為,Foxo6只在大腦中表達,因此對Foxo6的研究相對較少,后來Foxo6也被證明在多個組織和器官中表達,但Foxo6在內皮細胞IR中的表達和作用以及其具體的作用機制目前也無相關報道。尤其是近幾年來,有很多關于內皮細胞IR的研究,但具體作用機制還不完全清楚[16]。在實驗中用HG/HF處理HUVECs 24 h成功構建IR模型,觀察Foxos在其中的表達和變化,結果發現Foxo6基因變化最顯著,并且在HG/HF組中表達量升高,因此,選擇Foxo6這一個變化最明顯的亞型來進行后續研究。隨后,構建了沉默Foxo6的siRNA,并從中篩選出沉默效果最明顯的一條siRNA3鏈進行后續實驗,通過檢測NO、ET-1、AKT-pS473等指標證明沉默Foxo6可以逆轉IR。接下來進一步探討Foxo6促進IR的具體作用機制。造成IR的原因有多種,一是微血管損傷,二是胰島素信號轉導異常、三是線粒體功能障礙、四是炎癥,其中炎癥是一個重要方面,炎癥信號已經成為IR的驅動因素,同時大量基礎研究表明血管內皮的IR在心腦血管疾病的進展中起著重要作用,并與炎癥反應之間相互影響、相互促進[17]。NF-κB作為經典的炎癥通路,在調節IR中起著重要作用,但NF-κB在內皮細胞IR中起著什么作用,其是否和Foxo6存在相互作用呢?因此,又對Foxo6是否可以通過炎癥這條通路促進IR進行了探索?通過研究Foxo6是否與NF-κBp65相互作用以及Foxo6是否影響NF-κB的轉錄活性以及與其下游靶基因的表達,從而闡明Foxo6促進IR的具體作用機制。
4 討論
IR與T2DM密切相關,并加速著T2DM的進展,同時血管功能障礙也誘發了心腦血管疾病[11]。血管IR表現為血管內皮功能障礙,其原因主要是由于血管舒張異常所導致[12],血管內皮功能障礙不僅是血管疾病的最初階段,還是代謝性疾病的預測因子[13],此與機體長期高水平的血糖濃度和炎癥狀態密不可分,IR型的糖尿病患者常伴隨高血糖和高血脂,糖和脂又是我們日常生活中所必需的營養物質,攝入過多對人類的健康產生極大危害。近幾年來,有很多關于內皮細胞IR的研究,但具體的機制還不完全清楚。對于前期的研究工作已經證明HG/HF可以誘導內皮細胞IR,因此,弄清內皮細胞IR的作用機制對防治包括糖尿病血管并發癥和動脈粥樣硬化等疾病有重要意義。
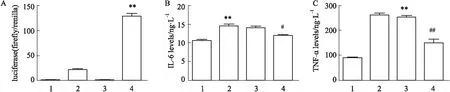
Fig 8 Effects of Foxo6 silencing on NF-κB transcriptional activity (A) and expression of target genes including IL-6 (B) and TNF-ɑ (C)
綜上所述,Foxo6在HG/HF應激誘導內皮細胞IR中的表達最顯著,并且表達量升高,其作用機制可能是Foxo6與NF-κBp65相互作用從而促進IR,Foxo6基因有可能作為T2DM治療的新靶點,同時為IR的治療提供一定的理論依據。但目前只是在細胞水平證明了Foxo6低表達可以逆轉IR,其在動物水平是否有同樣的結果還需要進一步驗證。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48