FKBP38基因的表達對小鼠非酒精性脂肪肝發生和發展的影響
2022-04-07 02:48:04唐敏怡邢朝鳳劉傲璐趙子建穆云萍汪麗娜李芳紅
中國藥理學通報 2022年4期
關鍵詞:小鼠
唐敏怡,王 帥,邢朝鳳,劉傲璐,趙子建,穆云萍,汪麗娜,李芳紅
(廣東工業大學生物醫藥學院,廣東 廣州 510006)
非酒精性脂肪性肝病(nonalcoholic fatty liver disease,NAFLD)是一種世界范圍內最常見的慢性肝臟疾病,包括非酒精性單純性脂肪肝(nonalcoholic fatty liver,NAFL)和非酒精性脂肪性肝炎(nonalcoholic steatohepatitis,NASH)[1]。NAFLD與肥胖、血脂異常、Ⅱ 型糖尿病等代謝疾病密切相關,因此,有專家提議將其稱為代謝相關脂肪肝病[2]。因為目前尚沒有獲批的用于治療NAFLD的藥物,所以臨床上治療NAFLD的方法主要是治療其合并癥,如肥胖、心血管疾病、Ⅱ 型糖尿病等。因此,探尋NAFLD的有效的治療方法已成為目前研究的熱點[3]。
FKBP38結合蛋白(FK506-binding protein 38)由于具有FKBP-C結構域而屬于FK506結合蛋白家族(FKBPs)成員之一,是一類參與免疫調節、蛋白折疊和轉運功能的免疫親和蛋白[4]。FKBP38蛋白具有一個跨膜結構域(TM)使其定位于線粒體等細胞內膜上,能夠招募抗凋亡蛋白Bcl-2和Bcl-XL至線粒體,抑制由于線粒體外膜受損而釋放可溶性蛋白(如細胞色素C),從而改善線粒體功能[5]。
研究表明,在NAFLD的發展過程中,為了抑制脂肪積累,線粒體脂肪酸氧化的增加起到了重要作用,但這種反饋會導致氧化應激,降低線粒體呼吸鏈活性,導致線粒體功能障礙,因此,線粒體功能在NAFLD發展過程中起著重要的作用,所以我們假設FKBP38基因的表達可能在NAFLD的發生和發展過程中起重要調控作用[6-7]。
因此,本研究使用蛋氨酸膽堿缺失飼料(methionine-and choline-deficent diet,MCD)誘導NAFLD小鼠模型,觀察肝細胞特異性敲除FKBP38基因后NAFLD小鼠肝臟病理變化,闡明FKBP38基因的表達對肝臟脂肪酸代謝的作用,為進一步探討FKBP38基因的表達影響NAFLD的發生和發展的作用機制奠定基礎,并為NAFLD的治療提供理論依據。
1 材料與方法
1.1 材料
1.1.1實驗動物 肝細胞特異性缺失FKBP38突變小鼠在廣東藥科大學SPF級實驗動物中心飼養,實驗動物飼養設施合格證號為 SYXK(粵)2017-0125。本實驗所用動物保護指南是由廣東藥科大學動物護理委員會所制定。
1.1.2主要試劑 蛋氨酸和膽堿缺失飼料(A02082002BR)購自江蘇省協同醫藥生物工程有限責任公司;蘇木精伊紅染色試劑盒(C0105)、RIPA裂解液(P0013B)購自上海碧云天生物技術有限公司;飽和油紅O染色劑(G1260)購自北京索萊寶科技有限公司;甘油三酯檢測試劑盒(10010303)購自美國Cayman公司;免疫組織化學-兔二步法試劑盒(PV-6001)購自北京中杉金橋生物技術有限公司;FKBP38抗體(AF3580)購自美國RD公司;F4/80抗體(70076)購自美國CST公司;TRIzol(T9424)購自美國Sigma公司;RNA逆轉錄試劑(R223-01)、ChamQ Universal SYBR qPCR Master Mix(Q711-02)、One Step Mouse Genotyping Kit(PD101-01)購自南京諾唯贊生物科技股份有限公司;引物由作者在NCBI數據庫中設計,并由上海生工生物有限公司合成。
1.1.3儀器 全自動生化分析儀(深圳雷杜生命科學股份有限公司);低溫離心機(美國Thermo);石蠟包埋機(德國SLEE);冷凍臺(中威電子儀器有限公司);石蠟切片機(美國Thermo);冰凍切片機(美國Thermo);正置顯微鏡(德國Leica);Cytation5多功能微孔檢測分析系統(美國Biotek);Chemi Doc+XRS化學發光凝膠成像系統(美國Bio-Rad);Cycler PCR儀(美國Bio-Rad);-80 ℃冰箱(中國美菱);qTOWER3G analytic(德國Jena)。
1.2 方法
1.2.1動物分組及NAFLD模型小鼠的建立 按照基因型將6周齡的小鼠分為野生型組(WT)以及純合敲除組(L-FKBP38-/-)2組,每組6只,所有小鼠(每籠6只)飼養在SPF可控環境內,恒溫22 ℃,相對濕度60%~70%,光照、黑暗周期為12 h,并自由進食蛋氨酸和膽堿缺失飼料和高壓滅菌水。小鼠喂食MCD 4周后,過夜禁食12 h,用三溴乙醇進行腹腔麻醉。從小鼠眼眶收集全血樣品,并在4 ℃下以12 000×g離心20 min來收集血清,用于檢測肝功能指標及血脂指標。最后,取小鼠最大肝葉分為兩份固定在4 %多聚甲醛中,用于肝臟病理分析,剩余的肝臟組織在液氮中快速冷凍,并儲存在-80 ℃,用于生化分析。使用以下引物對所有小鼠進行基因型鑒定,以區分野生型和純合敲除基因型。FKBP38-Flox上游引物1:5′-TCATGCAGGCAGAGTGCACATAG-3′,下游引物1:5′-CTGGAACTGGAGACTGGCTCCA-3′;上游引物2:5′-ACTGAGGCCACACAATCTAGGT-3′,下游引物2:5′-TGGAGACAGGCATTTCAGATCACA-3′; Alb-Cre引物1:5′-GCGGTCTGGCAGTAAAAACTATC-3′,引物2:5′-GTGAAACAGCATTGCTGTCACTT-3′,引物3:5′-CTAGGCCACAGAATTGAAAGATCT-3′,引物4:5′-GTAGGTGGAAATTCTAGCATCATCC-3′
1.2.2血清中肝臟損傷及血脂指標 取小鼠血清用全自動生化分析儀檢測血清中丙氨酸轉氨酶(alanine aminotransferase,ALT)、天冬轉氨酶(aspartate aminotransferase,AST)、甘油三酯(triglyceride,TG)和總膽固醇(cholesterol,CHO)水平。
1.2.3肝臟組織病理學染色
1.2.3.1 HE染色 肝臟組織用4%多聚甲醛固定后用乙醇溶液梯度脫水,石蠟包埋,制作成厚度為5 μm的石蠟切片。使用蘇木精伊紅染色試劑盒進行染色。
1.2.3.2 油紅染色 肝臟組織用4%多聚甲醛固定后用蔗糖溶液梯度脫水,OCT包埋溶膠包埋,制作成厚度為10 μm的冰凍切片。使用飽和油紅O染色劑進行染色。
1.2.3.3 免疫組織化學染色 肝臟組織用4%多聚甲醛固定后用乙醇溶液梯度脫水,石蠟包埋,制成厚度為5 μm的石蠟切片。使用F4/80抗體和免疫組織化學-兔二步法試劑盒進行染色。
1.2.4肝臟組織甘油三酯水平檢測 使用NP40裂解液裂解肝臟組織,用研磨機研磨后按照甘油三酯提取檢測試劑盒說明書檢測肝臟中甘油三酯的水平。
1.2.5肝臟組織基因表達水平檢測 使用TRIzol法提取肝組織提取總RNA,按照RNA逆轉錄試劑將RNA逆轉錄為cDNA,按照ChamQ Universal SYBR qPCR Master Mix說明書對FKBP38、過氧化物酶體增殖物激活受體α(peroxisome proliferators-activated receptors,PPAR-α)、過氧化物酶酰基輔酶A氧化酶1(Acyl Coenzyme A oxidase 1,ACOX-1)、肉毒堿棕櫚酰基轉移酶1α(carnitine palmitoyltransferase-1α,CPT-1α)、沉默調節蛋白3(sirtuin 3,SIRT3)基因進行實時熒光定量PCR (Quantitative Real-time PCR,qRT-PCR)實驗。
1.2.6肝臟組織蛋白表達水平檢測 使用RIPA裂解液裂解肝臟組織,在研磨機研磨后在4 ℃下以13 000×g離心15 min,取上清,于100 ℃金屬浴變性10 min,蛋白質免疫印跡法檢測組織中FKBP38蛋白的表達水平。
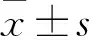
2 結果
2.1 肝細胞特異性敲除FKBP38模型鼠的鑒定利用本課題組前期構建的肝細胞特異性缺失FKBP38突變小鼠進行合籠實驗[8],剪取子代小鼠的腳趾提取DNA,通過聚合酶鏈式反應(polymerase chain reaction,PCR)對小鼠基因型進行鑒定,并使用FKBP38-Flox和Alb-Cre特異性引物區分野生型、雜合敲除基因型和純合敲除基因型小鼠(Fig 1A)。將Alb-Cre/-FKBP38fl/fl小鼠作為野生型組(WT),Alb-Cre/+FKBP38fl/fl突變小鼠作為純合敲除組(L-FKBP38-/-),在MCD造模后,進一步利用qRT-PCR技術和Western blot技術來檢測小鼠肝臟中FKBP38基因及蛋白的表達水平。本研究結果顯示,與野生型組(WT)相比,純合敲除組(L-FKBP38-/-)肝臟中的FKBP38基因轉錄水平和蛋白表達水平明顯降低(P<0.01)(Fig 1B,C),表明L-FKBP38-/-組小鼠肝臟中無FKBP38基因的表達。

Fig 1 Generation of FKBP38 hepatocellular specific knockout
2.2 FKBP38基因的表達對MCD誘導的非酒精性脂肪肝小鼠肝臟的影響
2.2.1對小鼠肝臟損傷的影響 為了探討肝細胞特異性敲除FKBP38基因對小鼠肝臟損傷的影響,使用全自動生化分析儀檢測小鼠血清中ALT和AST的水平。發現與WT組相比,L-FKBP38-/-組小鼠血清中的肝功能指標ALT和AST水平明顯升高(P<0.05)(Fig 2A,B)。進一步對小鼠肝臟切片進行HE染色觀察,發現與WT組相比,L-FKBP38-/-組小鼠肝臟肝細胞氣球樣變、肝細胞脂肪空泡及炎癥細胞浸潤更為明顯(Fig 2C)。以上結果表明,肝細胞特異性敲除FKBP38基因加劇小鼠由于MCD誘導的肝臟損傷。
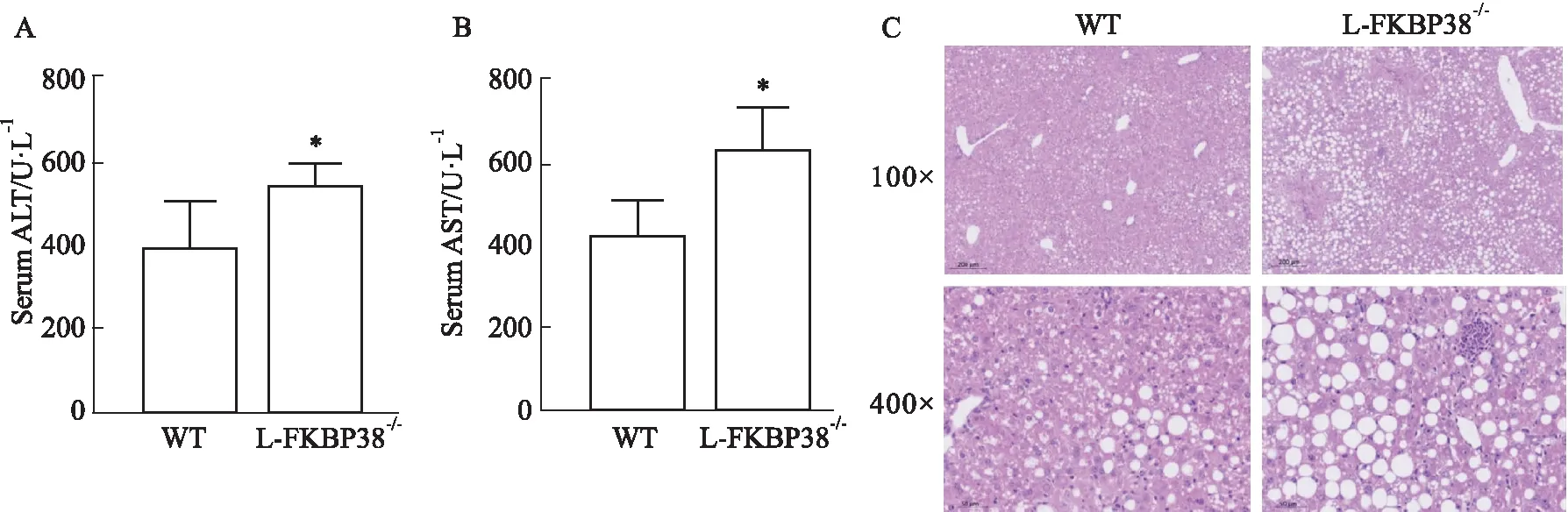
Fig 2 Effect of FKBP38 expression on liver damage in mice with
2.2.2對小鼠肝臟脂質積累的影響 為了探討肝細胞特異性敲除FKBP38基因對小鼠肝臟脂質積累的影響,使用全自動生化分析儀檢測小鼠血清中TG和CHO的水平。與WT組相比,L-FKBP38-/-組小鼠血清中的TG和CHO升高(P<0.05)(Fig 3A,B)。進一步對肝臟切片進行油紅染色觀察,發現與WT組相比,L-FKBP38-/-組小鼠肝臟脂滴數量更多而且面積更大(Fig 3C)。并且檢測肝臟中的TG含量,發現與WT組相比,L-FKBP38-/-組小鼠肝臟中的TG含量升高(P<0.05)(Fig 3D)。以上結果表明,肝細胞特異性敲除FKBP38基因加劇小鼠由于MCD誘導的脂肪積累。
Fig 3 Effect of FKBP38 expression on liver lipid accumulation in mice with
2.2.3對小鼠肝臟巨噬細胞的影響 為了探討肝細胞特異性敲除FKBP38基因對小鼠肝臟炎癥浸潤的影響,通過免疫組織化學染色標記小鼠肝臟中成熟的巨噬細胞表面糖蛋白—小鼠含生長因子的粘蛋白樣激素受體1(mouse EGF-like module-containing mucin-like hormone receptor-like 1,F4/80)的表達情況。發現與WT組相比,L-FKBP38-/-組小鼠肝臟F4/80陽性率明顯增加,說明L-FKBP38-/-組小鼠肝臟中成熟的巨噬細胞數量更多(Fig 4)。以上結果表明,肝細胞特異性敲除FKBP38基因加劇小鼠由于MCD誘導的肝臟巨噬細胞激活。
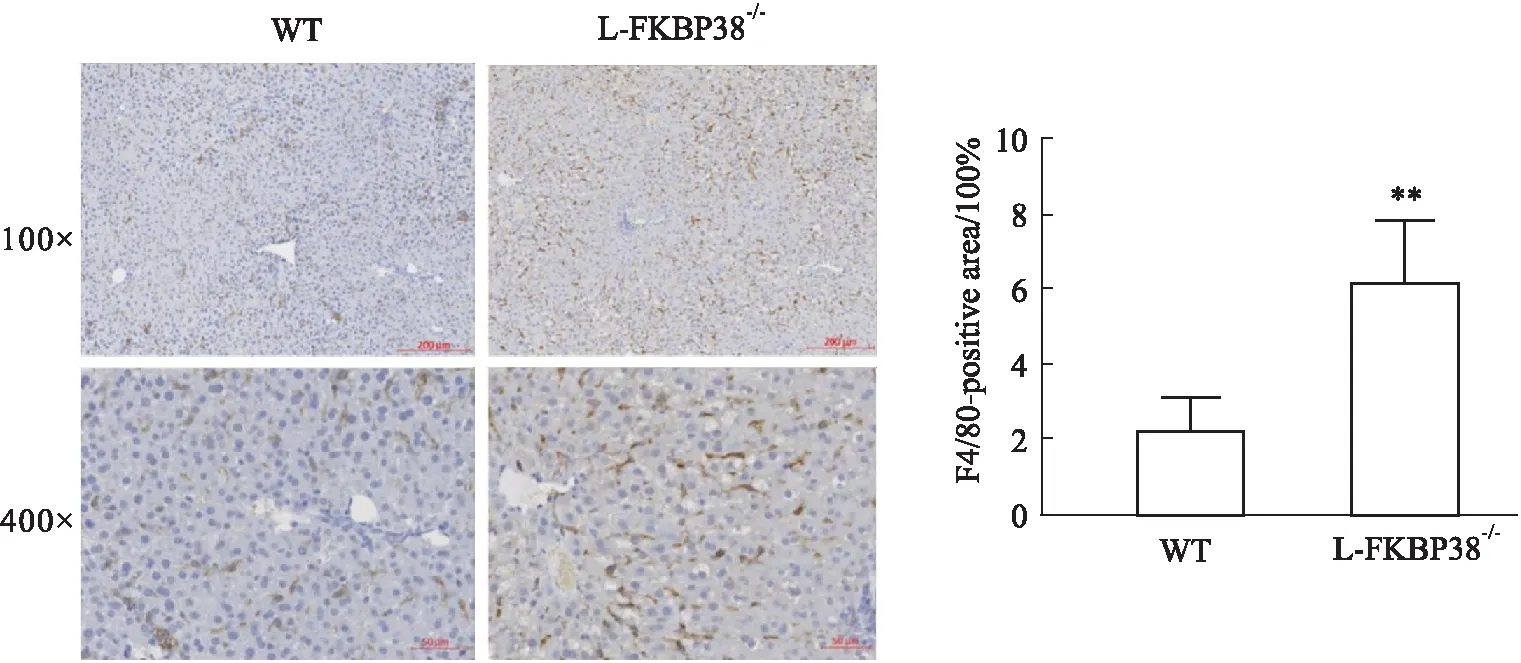
Fig 4 Effect of FKBP38 expression on liver F4/80
2.3 FKBP38基因的表達對MCD誘導的非酒精性脂肪肝小鼠肝臟脂肪酸氧化相關基因表達的影響為了探討肝細胞特異性敲除FKBP38基因如何加劇小鼠非酒精性脂肪肝的發生,通過qRT-PCR檢測小鼠肝臟中脂質代謝相關基因的表達情況。發現與WT組相比,L-FKBP38-/-組肝臟中PPARα的表達明顯下調(P<0.01)(Fig 5A)。并且與WT組相比,L-FKBP38-/-組肝臟脂肪酸β氧化相關基因ACOX-1、CPT-1α、SIRT3的表達明顯下調(P<0.05)(Fig 5B-D)。以上結果表明,肝細胞特異性敲除FKBP38基因抑制小鼠由于MCD誘導的肝細胞脂肪酸β氧化。
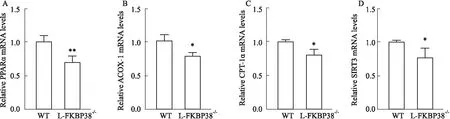
Fig 5 Effect of FKBP38 expression on expression of fatty acid oxidation gene in mice with
3 討論
NAFLD是指除了酒精和其他明確的損傷肝臟因素所導致的,以彌漫性肝細胞大泡性脂肪病變為主要病理特征的疾病,NASH為NAFLD的一種嚴重狀態,伴隨著肝細胞損傷、脂質積累、炎癥和纖維化,后期會發展為肝硬化和肝細胞癌[9]。在過去20年,NAFLD的全球患病率約為0.25,近年來,中國人的飲食結構向高脂高糖的西方飲食結構改變,生活水平的提高和不良生活方式的蔓延導致中國NAFLD患病率急劇上升,并呈年輕化趨勢,NAFLD已超過慢性乙型肝炎成為我國第一大肝病,據估計,到2030年,中國NAFLD患者將超過3億,中國是全球NAFLD患病率增長最快的國家[10]。大部分NAFLD患者對單純性脂肪肝不重視,隨著時間的推移,1/3的單純性脂肪肝可能會加劇為非酒精性脂肪肝肝炎,因而提高了臨床治療的難度。
NASH的發病機制尚不明確,但是目前較為公認的是二次打擊學說[11]。肝臟需要處理大量來自外周脂肪組織和胃腸道的脂質信號分子。當流入肝臟中的脂肪酸過多,而氧化和流出減少時會導致肝臟甘油三酯堆積,使肝臟脂質代謝紊亂,對肝臟進行第一次打擊,產生脂肪肝。代謝紊亂會激活巨噬細胞,使巨噬細胞從抗炎型M2亞型向促炎型M1亞型轉化,從而觸發炎癥反應,引起氧化應激、胰島素抵抗、線粒體功能障礙等,對肝臟進行第二次打擊,發展非酒精性脂肪性肝炎及纖維化[12]。
目前,治療NASH的藥物主要在Ⅱ期和Ⅲ期臨床試驗中,根據藥物在肝臟內的作用靶點進行分類,主要包括調節脂質和葡萄糖穩態、肝細胞內氧化應激、線粒體功能障礙、炎癥信號向肝細胞聚集的靶點等[3]。導致NAFLD發生和發展的分子途徑有很多,因此,探討NAFLD的發病機制及開發有效藥物已成為目前研究熱點。
研究表明[13],用MCD喂養C57BL/6小鼠能夠誘導小鼠肝臟脂肪變性,從而產生炎癥和纖維化,且其肝臟病理變化類似于人類NAFLD的組織學特征。因此,本研究采用MCD喂養小鼠4周產生NAFLD,以探討肝細胞FKBP38基因的表達對NAFLD模型小鼠肝臟的影響。本研究結果發現,在誘導小鼠產生脂肪肝病變后,與WT組相比,L-FKBP38-/-組小鼠血清中肝功能指標ALT、AST的水平顯著升高,肝細胞氣球樣變更嚴重,說明L-FKBP38-/-組小鼠肝臟受損更嚴重。同時,與WT組相比,L-FKBP38-/-組小鼠血清TG和CHO水平以及肝臟TG水平顯著升高,肝臟中脂滴數量顯著增多,說明肝細胞缺失FKBP38基因促進非酒精性脂肪肝的發生。我們進一步發現與WT組相比,L-FKBP38-/-組小鼠肝臟中F4/80免疫組織化學染色陽性比例增加,說明被激活的巨噬細胞顯著增加,可能意味著NAFLD發展為NASH。由于被激活的巨噬細胞中存在抗炎型M2亞型和促炎型M1亞型,兩者的比例在炎癥的發展過程中起著重要的作用,因此,本研究還需要進一步檢測小鼠肝臟中的被激活的M1亞型巨噬細胞的比例,以探討FKBP38基因的表達對NAFLD發展為NASH的過程中肝臟炎癥細胞浸潤的影響。此外,因為MCD誘導小鼠肝臟脂質積累的同時也會使小鼠因為缺乏營養而導致體重急劇下降,不能很好地模擬人類由于長期攝入高脂和高糖的食物而導致肥胖及胰島素抵抗等代謝疾病從而產生非酒精性脂肪肝的生理特征。因此本研究還需要進一步在小鼠營養過剩的條件下探討FKBP38基因的表達對NAFLD發生和發展的影響。
線粒體是脂肪酸氧化的主要場所,線粒體的功能對NASH的發生和發展起著重要的調控作用[6-7]。研究表明,當線粒體外膜完整性受損時,受Bcl-2家族蛋白調控,線粒體通過釋放可溶性蛋白(如細胞色素C)促進細胞凋亡[5]。而shirane等[14]的研究發現,在使用移動性離子載體A23187誘導HeLa細胞胞漿Ca2+濃度升高誘導細胞凋亡時,FKBP38基因的表達能減少發生凋亡的細胞數量,這種抑制細胞凋亡的作用是由FKBP38蛋白通過招募抗凋亡蛋白Bcl-2和Bcl-XL至線粒體來實現的。并且FKBP38蛋白能與自噬核心蛋白Atg8蛋白相互作用,將脂質化的自噬標記物LC3A募集到受損的線粒體從而誘導線粒體自噬,清除受損的線粒體[15]。因此,FKBP38蛋白可能在線粒體發揮正常功能的過程中起著重要的作用。
脂肪酸在線粒體中進行β氧化,產生三磷酸腺苷(ATP)為機體提供能量,而這個活動主要由過氧化物酶體增殖物激活受體PPARα調控[7]。在我們探討FKBP38基因的表達如何調控肝臟脂肪酸代謝的過程中,發現小鼠肝臟中PPAR-α的轉錄水平在L-FKBP38-/-組小鼠中明顯下調,我們猜測FKBP38基因的表達在線粒體脂肪酸β氧化的過程中發揮著重要的作用,于是我們進一步檢測脂肪酸β氧化中關鍵基因的表達。研究表明,極長鏈脂肪酸在ACOX-1的催化下脫氫形成脂酰CoA,再通過位于線粒體外膜的CPT-1進入線粒體,在脂肪酸β氧化酶系催化下,進行脫氫、加水,再脫氫和硫解連續反應,最后生成乙酰CoA用于三羧酸循環提供能量[16]。在這一系列的反應中,一些脂肪酸β-氧化代謝酶如長鏈酰基CoA脫氫酶(LCAD)容易發生過度乙酰化修飾而失去活性,而SIRT3是肝臟線粒體中重要的去乙酰化酶,SIRT3基因表達下調會使線粒體中一些代謝酶過度乙酰化,從而抑制脂肪酸β氧化[17]。我們發現小鼠肝臟中ACOX-1、CPT-1α和SIRT3的轉錄水平在L-FKBP38-/-組小鼠中顯著下調。說明FKBP38基因的表達在線粒體脂肪酸β氧化的過程中發揮著重要的作用,而FKBP38基因的表達如何影響線粒體脂肪酸β氧化仍需進一步深入研究。
此外,FKBP38蛋白作為一種能夠調節線粒體功能和抑制細胞凋亡的蛋白,其與癌癥發生和發展有著密切的關系,近年來受到研究人員廣泛關注。研究表明,FKBP38基因的表達能降低AM3Ca細胞的增殖、遷移和侵襲能力,與子宮內膜癌的發生顯著相關[18]。在既往的研究中,研究者大多關注FKBP38蛋白在腫瘤發生和發展過程中的作用,而沒有研究針對FKBP38蛋白在脂質代謝中的作用機制,本研究為FKBP38基因的表達在脂質代謝中發揮的功能提供新思路。
綜上所述,本研究結果表明,肝細胞特異性缺失FKBP38突變小鼠通過抑制肝臟脂肪酸氧化加劇脂質積累,損傷肝細胞,從而加劇非酒精性脂肪肝。通過本研究,我們首次發現FKBP38基因的表達在脂質代謝中發揮著重要作用,并為NAFLD的治療提供理論依據。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34