肝癌細胞侖伐替尼耐藥的基因篩選及其通路研究
2022-04-08 10:22:54陳家誠劉路政陳良陳騁許達峰林仕勛羅相相武金才
肝膽胰外科雜志 2022年3期
陳家誠,劉路政,陳良,陳騁,許達峰,林仕勛,羅相相,武金才
(海南省人民醫院/海南醫學院附屬海南醫院 肝膽胰外科,海南 海口 570311)
原發性肝細胞肝癌(以下簡稱為肝癌)在我國發病率和病死率均較高,雖然手術切除是肝癌治療的主要手段,但大部分肝癌患者確診時腫瘤已處于中晚期,失去手術機會[1]。近年來,靶向藥物及免疫治療逐漸被應用于中晚期或術后復發轉移的肝癌患者,為腫瘤降級轉化后外科治療創造了機會,也顯著延長了患者的總生存期[1-2]。侖伐替尼(Lenvatinib)作為新一線肝癌靶向治療藥物在中晚期肝癌治療中取得一系列亮眼的成績,但其耐藥問題也逐漸成為患者改善預后的主要障礙。然而,侖伐替尼耐藥的潛在分子機制尚不清楚。本實驗基于CRISPR/Cas9 sgRNA全基因組文庫結合Affymetrix表達譜芯片篩選出侖伐替尼耐藥相關基因THOC2,并探索其具體分子機制,現報道如下。
1 材料和方法
1.1 實驗材料
人SMMC-7721、MHCC-97H及MHCC-LM3肝癌細胞購于和元生物技術(上海)股份有限公司,細胞均無支原體等污染。侖伐替尼敏感與耐藥肝癌患者癌組織與癌旁組織樣本(各6例)取自海南省人民醫院肝膽胰外科標本庫,并遵從《赫爾辛基宣言》要求,簽署倫理知情同意書。樣本自離體30 min內液氮速凍,并在-80℃冰箱內保存。穩定細胞株構建中聚凝胺、嘌呤霉素購買于德國Sigma公司,殺稻瘟菌素購于上海翊圣生物科技有限公司。所有細胞均在含10%的胎牛血清(購于美國GIBCO公司)和1%的雙抗(青霉素和鏈霉素,購于美國Invitrogen公司)條件的杜氏改良培養基(購于美國GIBCO公司)中培養,細胞傳代胰酶購于美國Invitrogen公司。THOC2、BAX、cyclinD1、c-Myc(一抗)購于上海艾博抗公司,磷酸甘油醛脫氫酶GAPDP(一抗)、二抗、BCA蛋白濃度測定試劑盒、超敏ECL化學發光試劑盒購買于上海碧云天生物技術有限公司,IC50 CellTiter-Glo?試劑盒購于美國Promega公司,免疫共沉淀試劑盒購于美國賽默飛公司。侖伐替尼購買于日本衛材公司。
1.2 穩定細胞株的構建
構建對照慢病毒(空載體Empty)和目的病毒THOC2,然后將其感染SMMC-7721細胞。72 h以后,加入終濃度5 μg/mL的殺稻瘟菌素。每隔2~3 d更換1次含5 μg/mL殺稻瘟菌素的新鮮培養基,配合藥物篩選獲得穩定THOC2高表達的SMMC-7721-THOC2和對照SMMC-7721-Empty肝癌細胞。
1.3 CRISPR/Cas9 sgRNA全基因組文庫篩選
選擇人SMMC-7721 肝癌細胞,支原體檢測合格后,行細胞倍增時間測試、Puromycin篩選預實驗;IC50 實驗(共3 次)及藥物的最適濃度確定:確定侖伐替尼作用SMMC-7721 細胞株48 h或72 h的IC50,以不同藥物濃度(μg/mL)處理細胞14 d,得到藥物濃度為2 μg/mL時細胞抑制率為36.42%,并選擇以此濃度進行后續正式實驗。文庫慢病毒擴增包裝:行Cas9 sgRNA文庫擴增,包裝慢病毒;Cas9-sgRNA文庫(GeCKO library,含19 050個靶向基因和1 864個microRNA,且每個靶點設計6條sgRNA,共計12 萬種慢病毒載體)慢病毒感染:大規模擴增SMMC-7721細胞,感染慢病毒,并控制MOI,使大部分細胞只感染一個病毒,嘌呤霉素藥篩;細胞加藥培養:加入侖伐替尼(濃度為2 μg/mL)后共培養細胞,分別在第0天、7天、14天收耐藥細胞;sgRNA擴增:細胞基因組DNA純化后使用特異引物擴增sgRNA;高通量測序及數據分析:高通量測序定量分析sgRNA在3 個時間點的變化,篩選侖伐替尼敏感相關的候選基因,包括:測序數據質量分析;測序數據分布等。
1.4 Affymetrix基因表達譜芯片
將待檢測的12 例侖伐替尼耐藥組及敏感組肝癌組織進行研磨預處理,使用TRIzol法抽提組織總RNA,并使用天根RNeasy試劑盒進一步的純化。反轉錄合成雙鏈cDNA,準備IVT混合液,體外轉錄合成標記cRNA,提前在50~60 ℃預熱aRNA Elution Solution,10 min后進行aRNA純化。準備aRNA片斷化混合液并進行片斷化反應,室溫平衡芯片,同時加適量預雜交液至芯片,芯片預熱10 min。雜交液于99 ℃加熱5 min后,45 ℃放置5 min,最大速度離心雜交液5 min。將處理后雜交液加至芯片,45 ℃,60 rpm雜交16 h。最后,進行洗滌、染色和芯片掃描分析。
1.5 逆轉錄聚合酶鏈式反應(reverse transcription-PCR,RT-PCR)
采用兩步法進行RT-PCR。首先,將已抽提好的肝癌組織和細胞株RNA和稀釋處理后的Oligo(dT)15混勻離心,70 ℃凈置5 min,立即冰水浴,稍離心。根據M-MLV反轉錄說明書配制反應體系,混勻后離心,完成逆轉錄反應。將PCR反應試劑按照比例配制包含引物(引物信息:THOC2 引物正向序列:5’-ACATTGCCTTGAAACAGGCG-3’,反向序列:5’-A CTGCCCAGAGTAGCCCATA-3’;GAPDH引物正向序列:5’-GAAGGTGAAGGTCGGAGTC-3’,反向引物序列:5’-GAAGATGGTGATGGGATTTC-3’),混勻,97 ℃反應5 min,立即冰水浴,混勻。經預變性、變性、退火、延伸及終末延伸共28~36個循環,4 ℃保存。最終,加(1~10)μL PCR產物和溴芬蘭(1~2.5)μL混勻,加樣,電泳。
1.6 Western blotting分析
提取已建立的SMMC-7721-THOC2 和SMMC-7721-Empty細胞的總蛋白并檢測濃度。十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳分離蛋白,濕轉法轉模。封閉,孵育一抗與GAPDH(5% BSA-PBST稀釋)4 ℃過夜。洗膜后孵育二抗。采用增強化學發光法顯色,全自動化學發光成像分析系統曝光,軟件Image J對圖像進行灰度分析。
1.7 免疫共沉淀(co-immunoprecipitation,Co-IP)分析
根據Co-IP試劑盒說明書,在上述穩定構建的SMMC-7721-THOC2細胞基礎上,使用雙琥珀酰亞胺辛二酸酯交聯抗體和磁珠,用含有蛋白酶抑制劑(Termo,Waltham,MA)的Pierce免疫共沉淀裂解緩沖洗裂解細胞。在4 ℃與細胞裂解液孵育過夜后,用細胞裂解緩沖液洗滌。從磁珠中分離出蛋白并進一步分析。
1.8 病理和免疫組織化學(immunohistoche-mical staining,IHC)染色分析
將臨床獲得的12例侖伐替尼敏感組與耐藥組肝癌患者癌組織分別制成石蠟切片,烘烤后經二甲苯、乙醇脫蠟,蘇木素染10 min,流水沖洗后70%和90%酒精中脫水各10 min,入酒精伊紅染色液染色2~3 min。再次脫水后經二甲苯透明后中性樹膠封片。上述對蠟片載玻片進行抗原回收處理。之后,組織切片冷卻和封閉,然后分別用相關抗體孵育。隨后,將腫瘤標本切片與酶標二抗混合。最后觀察載玻片。
1.9 生物信息學分析
采用腫瘤與癌癥基因組圖譜(The Cancer Genome Atlas,TCGA)數據集進行基因的表達和預后分析(http://starbase.sysu.edu.cn/panCancer.php),GCBI數據庫(https://www.gcbi.com.cn)預測基因相關轉錄因子。
1.10 統計學分析
使用Graphpad Prism 8軟件整理數據和繪圖,計量資料兩組比較采用t檢驗。生存分析采用Kaplan-Meier法,生存曲線采用Log-rank檢驗。P<0.05 表示差異有統計學意義。
2 結果
2.1 全基因組CRISPR/Cas9結合Affymetrix基因表達譜芯片篩選肝癌與侖伐替尼靶向耐藥相關基因
我們在Cas9藥篩第7天分析(Day7vsDay0)共發現339個具有敲除后藥效增敏作用的關鍵基因,即肝癌細胞對侖伐替尼不敏感或耐藥的相關基因;在給藥第14天收集分析(Day14vsDay0)共發現1 438個上述基因;且其中共53個基因出現在第7和14天的交集(Day7 vs Day14)中,提示這些基因可能具有更重要的研究價值。進一步,我們應用Affymetrix表達譜芯片分析了12 例肝癌組織樣本(其中侖伐替尼敏感組6例,耐藥組6例),結果提示共有1 233個差異表達基因(Fold change≥2)(見圖1A),其中THOC2、CTNNB1(β-catenin)、MET(c-Met/HGFR)、HTATIP2、C10orf11、PRDX3、MAST4等7個基因存在于上述(Day7vsDay14)53 個交集基因中(見圖1B、1C),其在侖伐替尼耐藥組中的表達顯著高于敏感組;且聚類分析發現該7個基因可以較好地將兩組區分開來(見圖1D)。
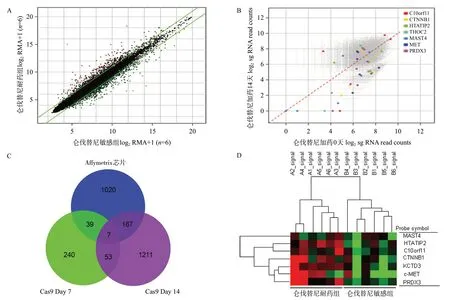
圖1 全基因組CRISPR/Cas9結合Affymetrix基因表達譜芯片篩選肝癌與侖伐替尼靶向耐藥相關基因
2.2 差異基因的生物信息學分析
我們使用TCGA 數據庫中LIHC(Liver Hepatocellular Carcinoma,肝細胞癌)的數據(http://starbase.sysu.edu.cn/panCancer.php)分析提示,上述7個耐藥基因中3個基因(C10orf11、PRDX3、MAST4)在癌組織與癌旁組織中表達水平均未見統計學差異,且除PRDX3外均無生存相關;其余4 個基因(β-catenin、c-Met、THOC2和HTATIP2)中,THOC2在癌與癌旁中表達存在顯著統計學差異(見圖2A),且與肝癌臨床分期及預后呈負相關(見圖2B、2C)。進一步對我院臨床肝癌及癌旁組織IHC檢測也提示THOC2蛋白在癌組織的表達(主要在細胞核)高于癌旁組織(見圖2D、2E、2F、2G)。
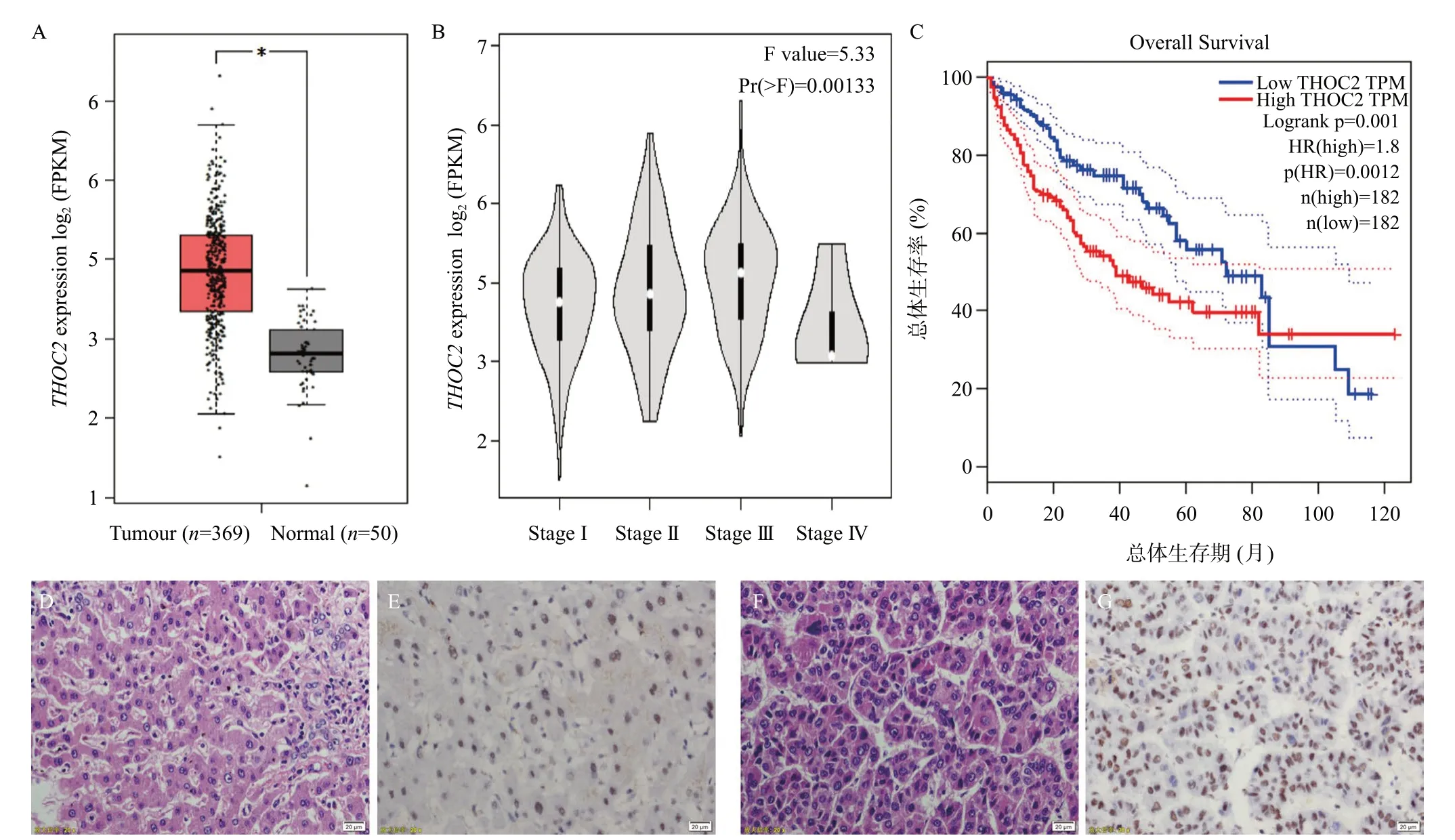
圖2 差異基因THOC2在TCGA數據庫及THOC2蛋白肝癌組織中的表達差異
2.3 TCF家族可作為THOC2核轉錄的潛在調控因子
進一步我們預測調控THOC2的轉錄因子發現:Wnt通路關鍵轉錄因子TCF蛋白家族可能調控THOC2進而影響下游Wnt通路(https://www.gcbi.com.cn/gcanalyze/html/generadar/search/singlegene/regregulat/THOC2),預測提示THOC2與TCF(TCF 1、3、4、11)存在結合位點(且存在SNP),進而參與調控Wnt/β-catenin通路(見圖3,表1)。
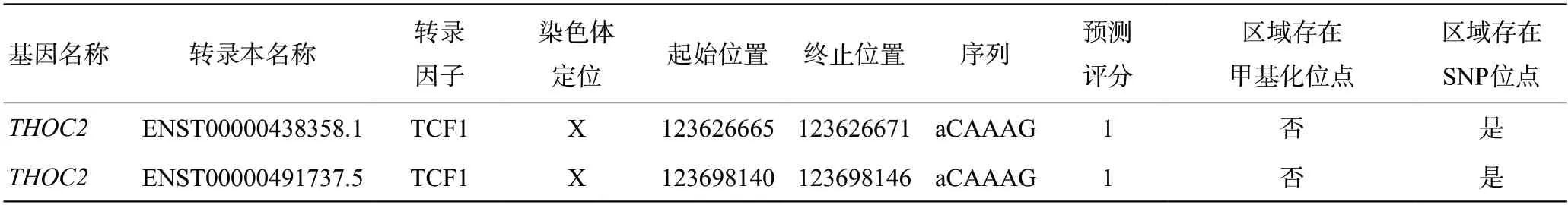
表1 THOC2可能結合位點
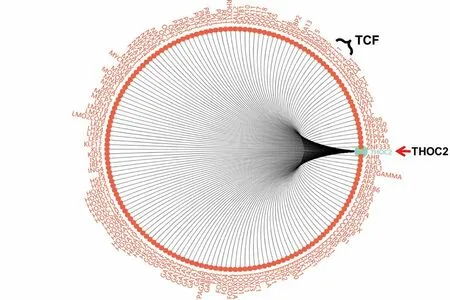
圖3 THOC2核轉錄的潛在調控因子預測分析顯示Wnt通路關鍵轉錄因子TCF蛋白家族與THOC2密切相關
2.4 THOC2與Wnt/β-catenin通路在肝癌侖伐替尼耐藥中的關系
我們構建THOC2過表達質粒并成功穩定轉染SMMC-7721 細胞(見圖4A),Western blotting檢測發現:與對照組細胞相比,高表達THOC2細胞(SMMC-7721-THOC2)中BAX、cyclinD1、c-myc和β-catenin的表達水平顯著增高(見圖4B、4C),提示THOC2表達可能導致Wnt/β-catenin經典通路下游靶基因的激活;在此基礎上,免疫共沉淀(Co-IP)初步分析提示THOC2與β-catenin存在密切相關(見圖4D)。進一步,IHC、RT-PCR和Western blotting檢測上述侖伐替尼敏感及耐藥組肝癌中THOC2和β-catenin的表達發現:THOC2和β-catenin在肝癌組織中存在差異化表達,THOC2在肝癌侖伐替尼耐藥組中的表達水平高于敏感組(見圖4E、4F、4G);同時分析顯示THOC2在SMMC-7721、MHCC-97H和MHCC-LM3不同肝癌細胞中亦存在差異表達,且在高轉移潛能肝癌細胞株中表達較高(見圖4H)。
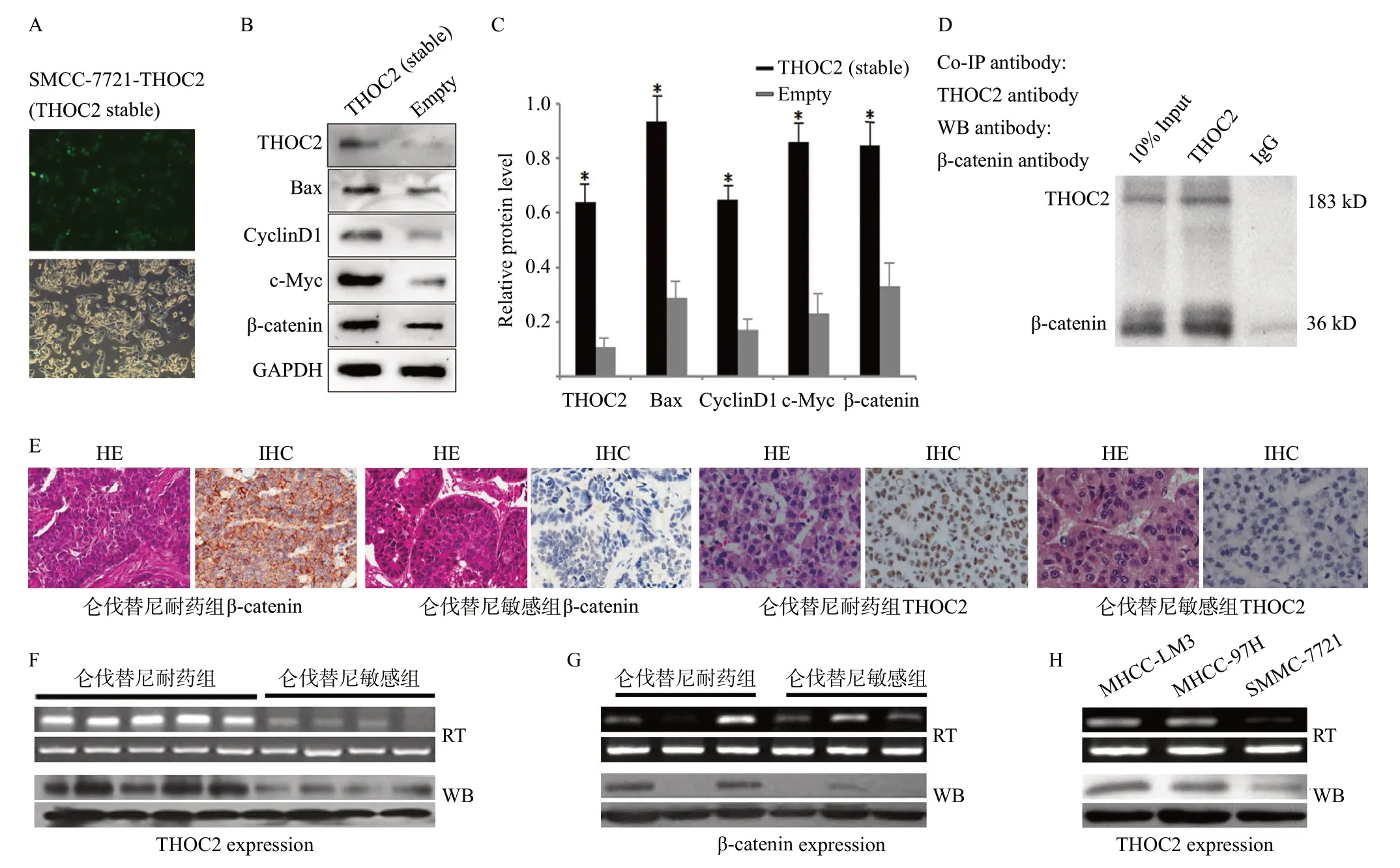
圖4 THOC2與Wnt/β-catenin通路在肝癌侖伐替尼耐藥中的關系分析
3 討論
肝癌是我國常見的惡性腫瘤之一,其發病率、病死率高,術后易復發及轉移,嚴重危害著我國公民的健康。2018年侖伐替尼獲FDA審核成了治療肝癌的新一線用藥,用于不可切除的肝細胞癌,改善了肝癌靶向治療缺藥的現狀[3]。我們近期的臨床實踐發現,部分中晚期肝癌患者在單藥服用侖伐替尼治療后,疾病尚未緩解甚至出現進展,提示侖伐替尼不敏感或耐藥。因此,明確肝癌侖伐替尼耐藥相關基因,早期檢測評估肝癌患者對侖伐替尼的敏感性并進行前瞻性用藥指導,具有重要的臨床意義。
侖伐替尼是口服多靶點的藥物,抑制參與腫瘤增殖的其他促血管生成和致癌信號通路相關RTK,主要靶點包括VEGFR1-3、FGFR1-4、PDGFR-a、KIT、RET等[4-5]。本實驗基于CRISPR-Cas9 sgRNA全基因組文庫篩選技術結合Affymetrix表達譜芯片,結果分析存在THOC2、CTNNB1(β-catenin)、MET(c-Met/HGFR)、HTATIP2、C10orf11、PRDX3、MAST4等7 個侖伐替尼耐藥相關基因。我們通過TCGA數據分析發現3 個基因(C10orf11、PRDX3、MAST4)在肝癌與癌旁中表達水平均未見統計學差異、且除PRDX3外均無生存相關。研究已表明:c-Met、HTATIP2、PRDX3、MAST4均可參與PI3K/ALK/Akt通路,其中c-Met尚可參與Ras-Rac/Rho通路、HTATIP2可介導JAK-STAT3、MAST4可參與PI3K-Akt-mTOR通路[6-9];而c-Met和HTATIP2已被發現可分別導致肝癌對侖伐替尼和索拉非尼耐藥[6,10];β-catenin可介導Wnt/β-catenin通路,促進肝癌發生發展,且與肝癌對索拉非尼耐藥密切相關[11-12]。β-catenin、c-Met和HTATIP2已在肝癌中已多有報到并與肝癌耐藥相關,尚未見THCO2在肝癌耐藥中可能的相關研究報道。
THOC2為THO復合物(THO complex)的一員,定位于染色體Xq25,含13.2 kb序列,編碼含1593氨基酸的蛋白(分子量為18.3kDa),常表達于細胞核中,參與mRNA轉錄,在轉錄延伸、核RNA輸出和基因組穩定中起著重要的調控作用[13-14]。在腫瘤相關研究中,THOC2可促進黑色素瘤侵襲增殖,并且在其在腫瘤組織高表達與預后相關[15]。在耐輻射三陰性乳腺癌(TNBC)細胞中,THCO2上調并通過促進性別決定區Y-盒轉錄因子SOX2 和同源盒轉錄因子NANOG轉錄產物的核釋放,以THOC5依賴的方式促進TNBC細胞的干性和抗輻射能力。我們通過TCGA數據庫分析表明,THOC2在肝癌較癌旁組織中高表達,且與患者臨床分期及預后密切相關。細胞水平發現上調THOC2(SMMC-7721細胞株)可使Wnt/β-catenin經典通路下游BAX、cyclinD1和c-myc蛋白水平顯著增高,免疫共沉淀同時表明THOC2與β-catenin存在密切相關。進一步IHC、RT-PCR和WB提示THOC2 和β-catenin在肝癌組織和中存在差異化表達,THOC2及β-catenin在侖伐替尼肝癌耐藥組中的表達高于敏感組,且THOC2在高轉移潛能肝癌細胞株MHCC-LM3、MHCC-97H中較SMMC-7721細胞表達增高。有相關研究報道,干擾THOC2 可明顯抑制肝癌細胞增殖和侵襲能力,并且外泌體THOC2聯合MYL6B可作為HCC預后和復發風險預測的分子標志物,這與我們的研究結果一致。因此,THOC2可能是導致肝癌侖伐替尼耐藥及復發轉移的關鍵基因。
綜上,我們基于CRISPR-Cas9 sgRNA全基因組耐藥篩選得到的THOC2基因高表達可介導Wnt/β-catenin通路并與侖伐替尼耐藥密切相關;THOC2基因在侖伐替尼肝癌靶向耐藥中可能具有潛在的重要臨床價值。
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年17期)2015-03-01 02:03:58
中國當代醫藥(2015年16期)2015-03-01 02:03:11
癌變·畸變·突變(2015年4期)2015-02-27 06:15:25
