香菇GPX基因克隆及其在采后貯藏中的表達分析
2022-04-24 15:31:46韓云云袁學(xué)文栗藝芳
保鮮與加工 2022年4期
韓云云,袁學(xué)文,栗藝芳,鄧 冰,*
(1.山西農(nóng)業(yè)大學(xué)資源環(huán)境學(xué)院,山西 晉中 030801;2.山西農(nóng)業(yè)大學(xué)食品科學(xué)與工程學(xué)院,山西 晉中 030801;3.山西省襄垣縣農(nóng)業(yè)農(nóng)村局,山西 長治 046200)
香菇(Lentinus edodes)屬擔子菌綱(Basidaiomycetes)傘菌目(Agaricales)口蘑科(Tricholomatacete)香菇屬(Lentinus),2020年國內(nèi)香菇總產(chǎn)量達到1 188.21萬t,在所有食用菌中居于首位[1]。鮮食香菇在貯藏過程中極易發(fā)生褐變,造成了嚴重的經(jīng)濟損失[2-3]。對雙孢菇和杏鮑菇的研究發(fā)現(xiàn),以酪氨酸酶(Tyrosinase,TYR)、雙酚氧化酶(Diphenoloxidase,DPO)和漆酶(Laccase,LAC)等為代表的多酚氧化酶(Poly phenol oxidase,PPO)是直接造成子實體褐變的關(guān)鍵酶之一[4]。TYR和LAC能催化酪氨酸(單酚化合物)形成3,4-二羥基苯丙氨酸,之后該化合物被進一步氧化成多巴醌,在非酶催化條件下多巴醌轉(zhuǎn)化為多巴色素(紅色)、5,6-二羥基吲哚和5,6-吲哚醌(黃色),并經(jīng)過聚合反應(yīng)形成黑色素(紫色)和黑素(黑色)等褐變物質(zhì)[4-7]。目前香菇采后褐變過程中發(fā)揮主要作用的酶種類尚無定論,同時褐變過程中涉及的生理生化機制和分子機制尚不明確。過氧化物酶(Peroxidase,POD)是真核生物抗氧化系統(tǒng)中的關(guān)鍵酶,在機體應(yīng)答生物脅迫和非生物脅迫相關(guān)的生物學(xué)過程中發(fā)揮重要作用[8-10]。另外有研究表明,在過氧化物氧化相關(guān)活性物質(zhì)產(chǎn)生色素沉積的過程中POD發(fā)揮了重要作用,而通過外源處理鈍化POD活性可以有效抑制香菇褐變發(fā)生,表明POD可能亦是造成食用菌褐變發(fā)生的關(guān)鍵酶[11-13]。谷胱甘肽過氧化物酶(Glutathione peroxidase,GPX)為含有巰基的POD酶類,目前研究發(fā)現(xiàn)GPX在調(diào)節(jié)機體生長發(fā)育、活性氧(Reactive oxygen species,ROS)代謝和抵抗壓力脅迫等過程中發(fā)揮作用,但是對于其在園藝產(chǎn)品采后褐變,尤其是以香菇為代表的食用菌采后褐變發(fā)生中的作用機制尚不清楚[14-15]。
本研究基于香菇基因組數(shù)據(jù)篩選香菇LeGPX基因,通過基因克隆明確其編碼序列,利用生物學(xué)軟件預(yù)測其基本生物學(xué)信息,并通過熒光定量PCR(qRTPCR)技術(shù)研究其在香菇采后貯藏中的表達模式,通過以上研究以期為明確香菇采后褐變機制提供依據(jù)。
1 材料與方法
1.1 材料與設(shè)備
1.1.1 材料與試劑
香菇:于2021年5月采自山西省臨縣山圪嶗農(nóng)業(yè)專業(yè)合作社,挑選菌蓋直徑一致、無機械傷且未開傘香菇,采摘后置于4℃貯藏。分別在貯藏0、4、8、12、16、24、28 d留取菌蓋和菌柄部位,所有樣品經(jīng)液氮速凍后置于超低溫(-80℃)冰箱保存。
總RNA提取試劑盒、克隆載體、DH5α感受態(tài)細胞和Transetta感受態(tài)細胞:北京全式金生物技術(shù)股份有限公司;反轉(zhuǎn)錄試劑盒、質(zhì)粒小提試劑盒、DNA純化試劑盒、無縫克隆試劑盒:南京諾唯贊生物科技有限公司;pCOLD-ProS2載體:寶日醫(yī)生物技術(shù)(北京)有限公司。
1.1.2 儀器與設(shè)備
NanoDrop ONE超微量分光光度計,美國賽默飛世爾科技公司;Mini-Sub cell GT電泳系統(tǒng),CFX96熒光定量PCR儀,美國伯樂科技公司;Tone96G梯度PCR儀,德國耶拿分析儀器股份公司;Tanon4100凝膠成像儀,上海天能科技有限公司;WSC-S型色差儀,上海儀電分析儀器有限公司。
1.2 方法
1.2.1 基因鑒定
通過基迪奧云平臺(https://www.omicshare.com/tools/)對課題組已獲得轉(zhuǎn)錄組數(shù)據(jù)中所有的編碼基因進行PFAM注釋,以E值<0.01為準提取包含GPX結(jié)構(gòu)的序列,之后利用NCBIBlastP確定蛋白的保守結(jié)構(gòu)域。使用IBS1.0對蛋白注釋結(jié)果和基因結(jié)構(gòu)進行繪制。
1.2.2 基因克隆
1.2.2.1 總RNA提取和反轉(zhuǎn)錄
參照RNA提取試劑盒說明書提供的方法進行凍存樣品總RNA的提取,用100μL無酶水溶解RNA沉淀,水平電泳檢測RNA提取質(zhì)量,NanoDrop ONE超微量分光光度計檢測RNA濃度和純度。以1μg總RNA為模板,參照反轉(zhuǎn)錄試劑盒說明進行反轉(zhuǎn)錄。反應(yīng)產(chǎn)物稀釋10倍,保存于-80℃冰箱備用。
1.2.2.2 基因克隆
參照“1.2.1”中鑒定的序列,使用Primer 5軟件設(shè)計用于PCR反應(yīng)的擴增引物(表1)。
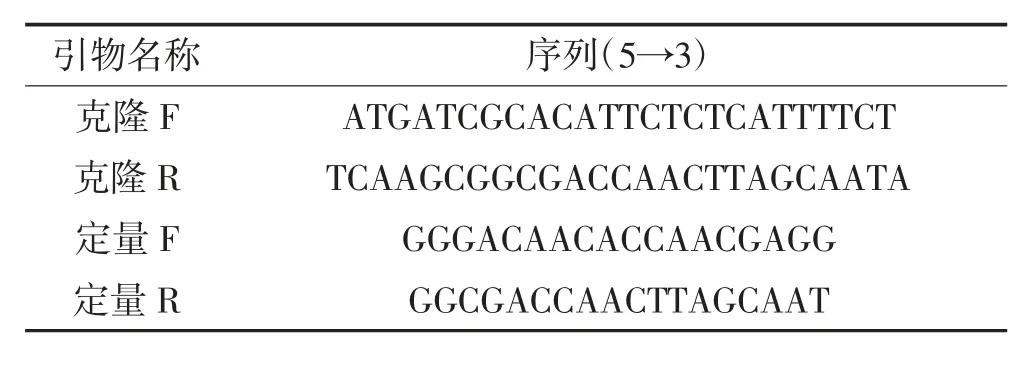
表1 本研究所用引物Table 1 Primers in thisstudy
PCR反應(yīng)體系:1μL模板,上游和下游引物(10μmol/L)各2μL,PCRSupermix25μL,無菌水20μL。
PCR反應(yīng)條件:95℃變性5 min,擴增28個循環(huán)(95℃30 s,55℃10 s,72℃50 s),72℃延伸10 min。水平電泳檢測PCR產(chǎn)物并回收目的片段凝膠,使用試劑盒純化目的片段。將純化產(chǎn)物連接至EASY cloning載體,重組載體轉(zhuǎn)入大腸桿菌感受態(tài)細胞。37℃過夜培養(yǎng)后用PCR驗證陽性菌落,委托上海生工生物工程有限公司進行測序。
1.2.2.3 系統(tǒng)發(fā)育樹的構(gòu)建及多序列比對
從NCBI下載部分真菌GPX蛋白序列,使用MEGA6.0軟件構(gòu)建系統(tǒng)發(fā)育樹。參照系統(tǒng)發(fā)育分析結(jié)果,使用BioEdit軟件對LeGPX蛋白與相近物種GPX蛋白進行多序列比對。
1.2.3 蛋白生物信息學(xué)分析
使用在線軟件Sequence Manipulation Suite對蛋白氨基酸組成、分子式和分子質(zhì)量、帶電荷氨基酸組成和等電點、不穩(wěn)定系數(shù)及總疏水指數(shù)進行預(yù)測。使用在線軟件Prabi預(yù)測二級結(jié)構(gòu)組成及分布,在線軟件SWISS-MODEL構(gòu)建蛋白質(zhì)三級結(jié)構(gòu)模型。使用在線軟件ExPASy-ProtScale分析蛋白親疏水基團組成和分布,Excel軟件繪制圖譜。使用在線軟件SignalP 4.1 Server掃描蛋白序列前60位氨基酸中信號肽分布,Excel軟件繪制圖譜。使用在線軟件TMpred Server掃描蛋白全位點跨膜結(jié)構(gòu),Excel軟件繪制圖譜。使用Cell-Ploc軟件預(yù)測亞細胞定位情況。
1.2.4 蛋白原核表達及純化
參照“1.2.2”獲得的序列和pCOLD-ProS2圖譜設(shè)計用于構(gòu)建重組載體的引物,經(jīng)片段純化和連接構(gòu)建原核表達載體,PCR驗證載體構(gòu)建成功后轉(zhuǎn)入Transetta(DE3)感受態(tài)細胞[16]。參照蘇俊等[16]的方法,采用15℃結(jié)合異丙基-β-D-硫代半乳糖苷(IPTG)(0、0.2、0.4、0.6、0.8、1.0 mmol/L)誘導(dǎo)蛋白表達,保存誘導(dǎo)前和誘導(dǎo)后菌液,十二烷基硫酸鈉(SDS)-聚丙烯酰胺凝膠電泳(PAGE)檢測蛋白表達情況。采用鎳離子親和純化方法對重組蛋白進行純化,保存貫穿液和各濃度咪唑洗脫液(30、50、100 mmol/L),SDS-PAGE檢測純化效果[17]。
1.2.5 相對表達量測定
將“1.2.2”獲得的cDNA稀釋10倍作為模板,參照確定的基因序列設(shè)計用于熒光定量PCR反應(yīng)的引物(表1),按熒光定量PCR試劑盒說明進行qRT-PCR反應(yīng)程序。以起始點為對比,使用2-ΔΔt方法進行不同樣品(均包含3個生物學(xué)重復(fù))LeGPX基因相對表達量計算[18]。
1.2.6 色差測定
分別于貯藏0、4、8、12、16、24、28 d,使用WSC-S型色差儀測定菌蓋表面標記部位及菌蓋切開后中心部位(菌肉)色差值。不同部位色差值測定均以每5個香菇為一組,分別設(shè)置3組重復(fù)。
1.2.7 數(shù)據(jù)統(tǒng)計及分析
使用IBM SPSSStatistics軟件對數(shù)據(jù)進行方差分析,使用Excel 2016軟件進行數(shù)據(jù)分析和繪圖。
2 結(jié)果與分析
2.1 LeGPX基因鑒定
在香菇轉(zhuǎn)錄組數(shù)據(jù)中篩選到1條與已知物種GPX基因具有同源性的編碼序列(Coding sequence,CDS)。對該序列編碼的蛋白進行PFAM注釋,結(jié)果顯示該蛋白48~156氨基酸殘基為GPX結(jié)構(gòu)域(圖1A),結(jié)合CDD注釋結(jié)果確定篩選到的序列為LeGPX基因的CDS。對LeGPX基因DNA序列進行分析,結(jié)果顯示該基因DNA序列長度839 bp,包含5個外顯子;對該基因上游2 kb序列進行啟動子位點預(yù)測,結(jié)果顯示該基因的啟動子區(qū)位于上游802bp左右(圖1B)。
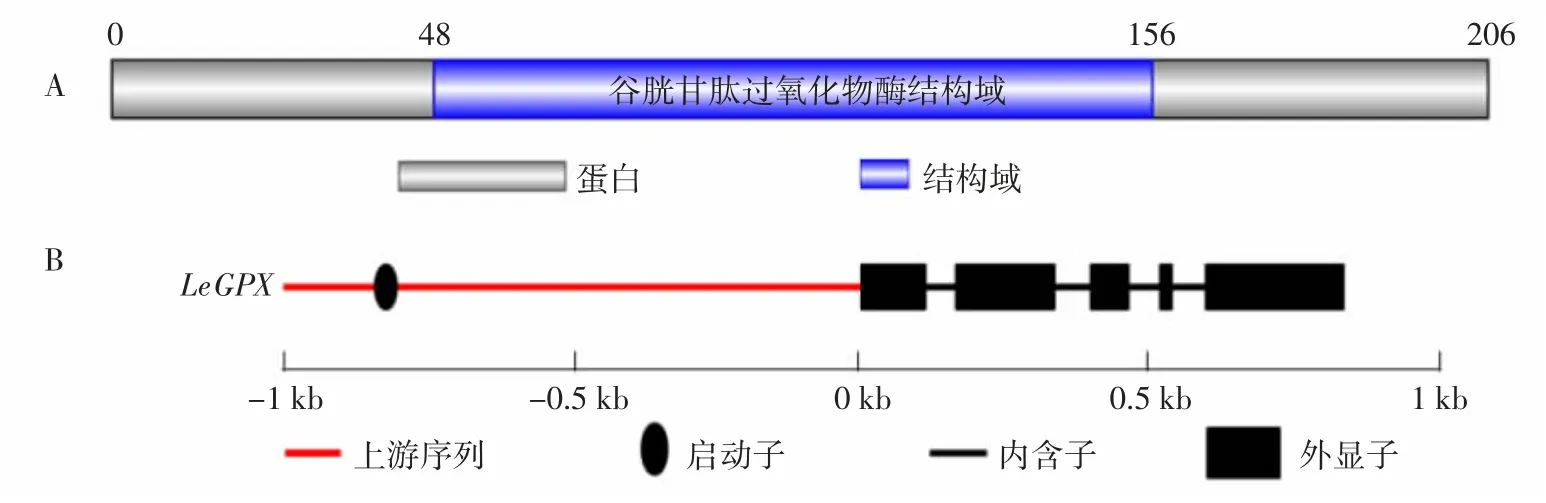
圖1 LeGPX基因PFAM注釋結(jié)果(A)及基因結(jié)構(gòu)(B)Fig.1 PFAMannotation result(A)and gene structure(B)of LeGPX
2.2 LeGPX基因克隆
參照鑒定到的LeGPX基因序列設(shè)計用于擴增CDS的引物,以香菇cDNA為模板進行PCR反應(yīng),結(jié)果顯示PCR產(chǎn)物符合預(yù)期大小(圖2A);將克隆得到的序列與香菇基因組和轉(zhuǎn)錄組數(shù)據(jù)進行比對,結(jié)果顯示所得序列與預(yù)測序列完全一致。為進一步驗證克隆結(jié)果的可靠性,參照已公布的其他真菌GPX氨基酸數(shù)據(jù)構(gòu)建了LeGPX蛋白系統(tǒng)發(fā)育樹,結(jié)果顯示LeGPX蛋白與小皮傘(Marasmiusfiardii)和灰色果腐菌(Moniliophthoraroreri)的GPX親緣關(guān)系較近(圖2B)。同時使用BioEdit軟件對LeGPX、MfGPX和MrGPX蛋白的氨基酸序列進行同源比對分析,結(jié)果顯示以上蛋白均包含3段GPX特征序列(NTASA(K)CGFT、I(V)L(V)AFPCNQF和KWNFEKFL(V))以及3個催化殘基位點(圖2C),符合GPX的一般特征[19]。以上結(jié)果證實已經(jīng)成功克隆了香菇LeGPX基因。
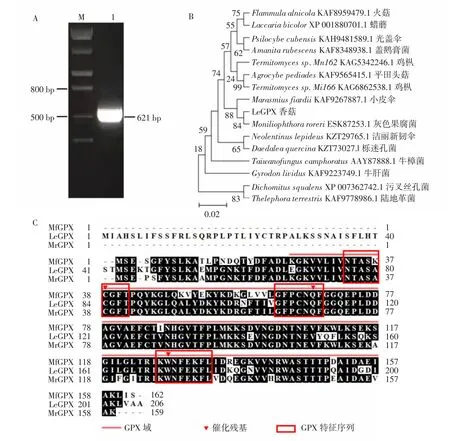
圖2 LeGPX基因克隆(A)、系統(tǒng)發(fā)育分析(B)及蛋白同源比對分析(C)Fig.2 Gene cloning(A),phylogenetic analysis(B)and homologouscomparison(C)analysisof LeGPX
2.3 LeGPX蛋白生物信息學(xué)分析
2.3.1 LeGPX理化性質(zhì)分析采用在線軟件Sequence Manipulation Suite預(yù)測LeGPX蛋白的理化性質(zhì)。LeGPX氨基酸殘基數(shù)為206,其中亮氨酸(Leu,8.70%)、蘇氨酸(Thr,7.80%)、丙氨酸(Ala,7.30%)和甘氨酸(Gly,7.30%)為該蛋白中占比最多的幾種氨基酸;分子式為C1029H1599N269O301S8,分子量為22.811 kD;帶正電荷氨基酸(精氨酸和賴氨酸)和帶負電荷氨基酸(天冬氨酸和谷氨酸)數(shù)量分別為16和44,理論等電點8.83,為堿性蛋白;不穩(wěn)定系數(shù)31.89(小于50),為穩(wěn)定蛋白。
2.3.2 LeGPX蛋白二級結(jié)構(gòu)和三級結(jié)構(gòu)分析
使用在線軟件Prabi對LeGPX蛋白的二級結(jié)構(gòu)進行預(yù)測。LeGPX蛋白的二級結(jié)構(gòu)以無規(guī)則卷曲(46.6%)和α-螺旋(29.1%)為主,延伸鏈和β折疊分別占17.5%和6.8%(圖3A)。使用在線軟件SWISS-MODEL對LeGPX蛋白的高級結(jié)構(gòu)進行預(yù)測,結(jié)果顯示無規(guī)則卷曲為該蛋白三級結(jié)構(gòu)中最明顯的組件;同時該蛋白三級結(jié)構(gòu)中還存在3個可能的催化殘基位點,與蛋白同源序列比對結(jié)果相符(圖3B)。
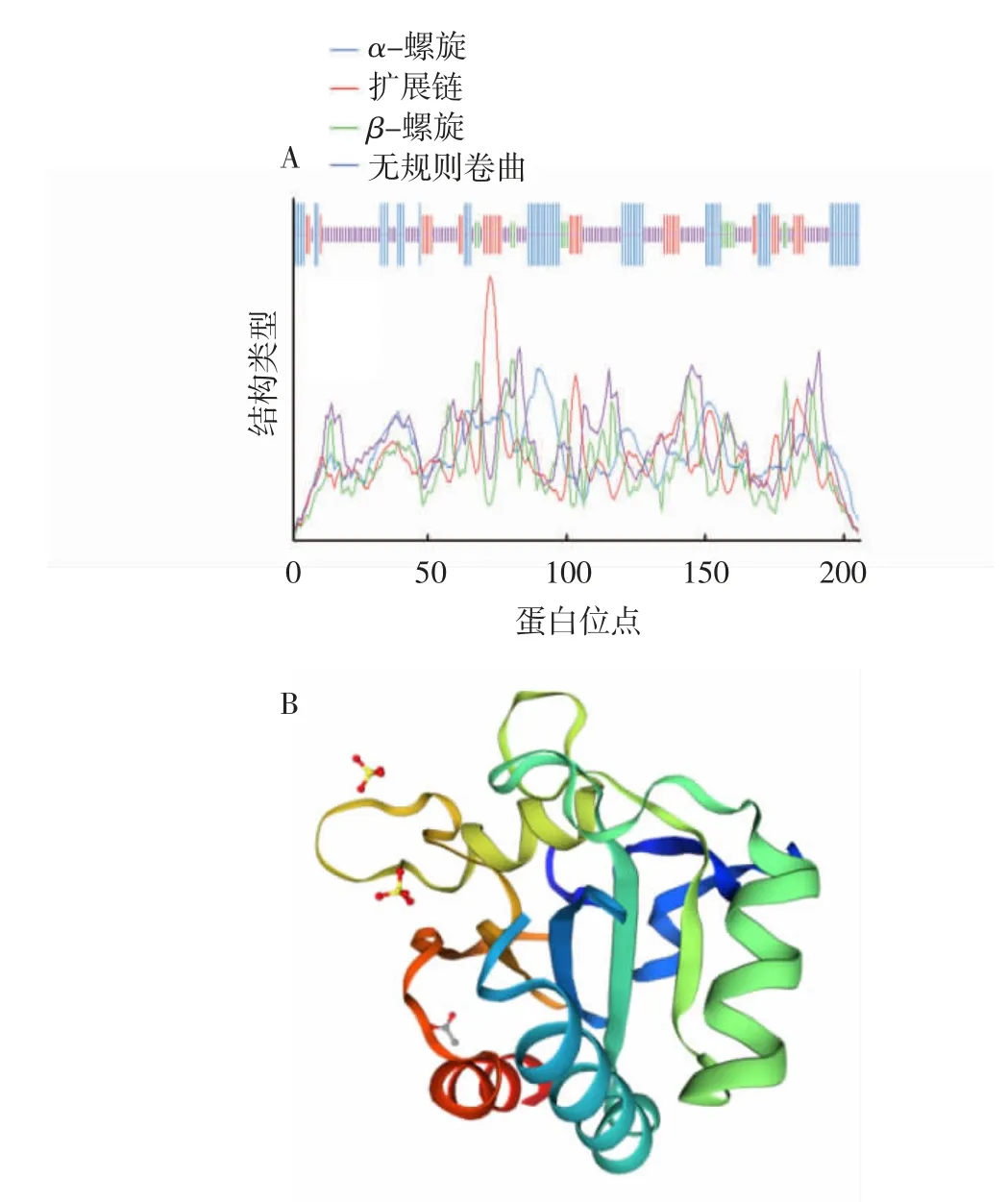
圖3 LeGPX蛋白二級結(jié)構(gòu)(A)和三級結(jié)構(gòu)(B)預(yù)測結(jié)果Fig.3 Theprediction of secondary structure(A)and tertiary structure(B)of LeGPX protein
2.3.3 LeGPX蛋白親水性分析
親水性分析結(jié)果顯示:LeGPX蛋白表面存在198個暴露的親疏水基團,其中氨基酸殘基第75位(纈氨酸)疏水性作用最強,為1.967;第98位(賴氨酸)疏水性作用最弱,為-2.511(圖4)。整體來看LeGPX蛋白表面的總親水位點數(shù)量(122個)多于總疏水位點(76個),整體疏水指數(shù)為-0.164(圖4),說明該蛋白為親水蛋白。
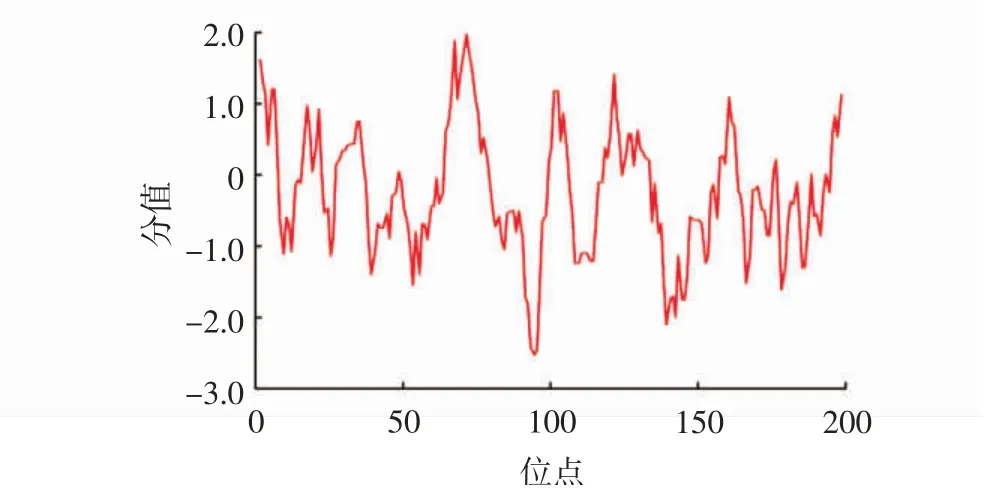
圖4 LeGPX蛋白親水性分析Fig.4 The hydrophilicity analysisof LeGPX protein
2.3.4 LeGPX蛋白細胞定位與信號肽分析
蛋白質(zhì)在合成后需要轉(zhuǎn)運至相應(yīng)位置發(fā)揮作用,明確細胞定位對于探明其生物學(xué)功能具有重要意義。使用SignalP4.1 Server軟件預(yù)測LeGPX蛋白信號肽,結(jié)果顯示前60位氨基酸中最高信號肽分值(Max.C)、最高原始剪切位點分值(Max.Y)和最高綜合剪切位點分值(Max.S)均小于0.5(圖5A),且信號肽預(yù)測結(jié)果顯示為不存在信號肽。使用在線軟件TMpred Server預(yù)測蛋白跨膜結(jié)構(gòu)域,結(jié)果顯示該蛋白不存在跨膜結(jié)構(gòu)(圖5B)。綜合信號肽和跨膜結(jié)構(gòu)域預(yù)測結(jié)果可以說明LeGPX蛋白為非分泌蛋白,該蛋白在合成之后不進入內(nèi)質(zhì)網(wǎng)-高爾基體分泌系統(tǒng),并且不存在進入其他細胞器的可能。使用在線軟件Cell-Ploc分析其亞細胞定位情況,結(jié)果亦顯示LeGPX蛋白定位于細胞質(zhì)。

圖5 LeGPX信號肽和跨膜結(jié)構(gòu)域分析Fig.5 Signal peptide and transmembrane structural domain analysisof LeGPX protein
2.4 香菇GPX蛋白原核表達分析
將構(gòu)建成功的pCOLD-ProS2-LeGPX載體轉(zhuǎn)進Transetta感受態(tài)細胞,經(jīng)低溫結(jié)合IPTG誘導(dǎo)產(chǎn)生了大小約為50 kD的重組蛋白(圖6中2~7泳道),并且對照組中未出現(xiàn)重組蛋白(圖6中1泳道)。LeGPX-ProS2重組蛋白表達成功,并且外源IPTG濃度不會影響該重組蛋白的積累(圖6中2~7泳道)。使用鎳離子親和純化方法對上清蛋白進行純化,結(jié)果發(fā)現(xiàn)采用50 mmol/L咪唑可有效洗脫LeGPX-ProS2重組蛋白(圖7)。
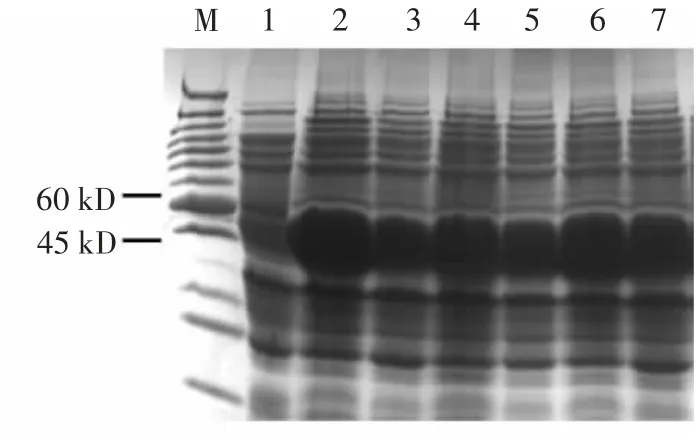
圖6 LeGPX蛋白原核表達Fig.6 Theprokaryotic expressionsof LeGPXprotein
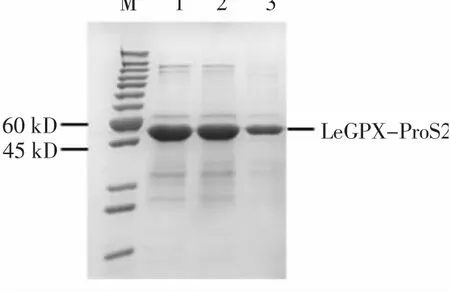
圖7 LeGPX-ProS2重組蛋白純化Fig.7 Purification of LeGPX-ProS2 recombinant protein
2.5 LeGPX基因在香菇采后貯藏期間的表達模式分析
香菇在采后貯藏期間會出現(xiàn)明顯的褐變現(xiàn)象,有研究表明該現(xiàn)象的發(fā)生可能與代謝產(chǎn)物積累導(dǎo)致的氧化還原失衡有關(guān)。如圖8A所示,香菇采后貯藏期間菌蓋表面L*值呈逐漸下降趨勢,反映在子實體上表現(xiàn)為表皮顏色由灰褐色逐步轉(zhuǎn)變?yōu)楹谏>獠课话l(fā)生褐變較晚,該部位L*值表現(xiàn)為貯藏初期無顯著變化,至貯藏中期(12~16 d)和貯藏末期(20~24 d)出現(xiàn)兩次顯著下降(圖8B)。通過相對表達量的分析發(fā)現(xiàn),菌蓋部位LeGPX基因表達量在貯藏期間整體呈先上升后下降的趨勢,采后8、12、16、20、28d時LeGPX基因的相對表達量均顯著高于貯藏初值(P<0.05),其中采后16、20 d時表達量達到初值12倍左右(圖8C)。菌柄部位LeGPX基因在貯藏初期(4~8 d)即顯著上調(diào)表達(15倍),貯藏12~28 d仍保持相對高表達狀態(tài)(圖8D)。整體來看,與菌蓋部位相比,香菇采后貯藏期間LeGPX基因在菌柄部位表達更為活躍
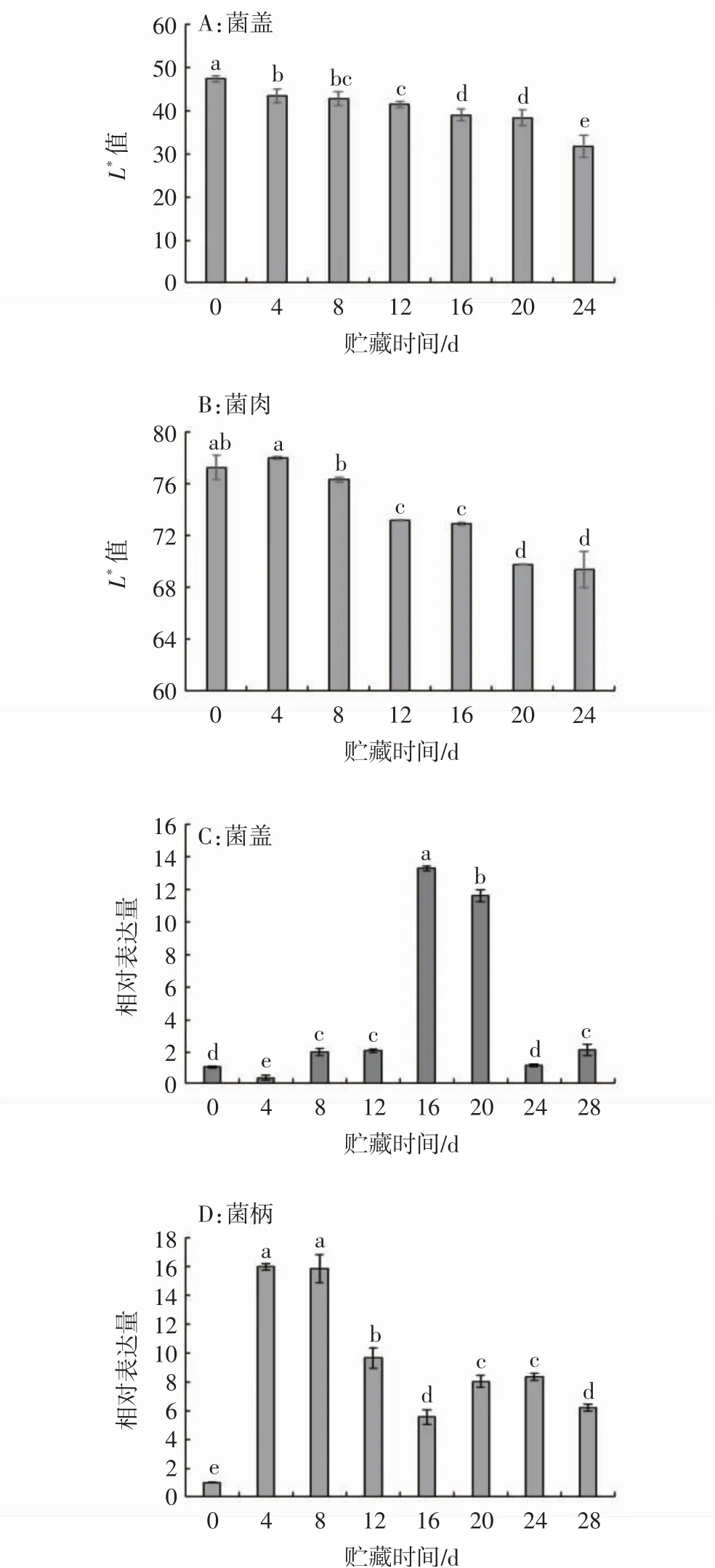
圖8 香菇采后L*值及LeGPX基因相對表達量變化Fig.8 L*valuesand the relative expression levels changesof LeGPX gene in postharvest L.edodes
3 討論與結(jié)論
真核生物氧化還原系統(tǒng)中存在豐富的抗氧化酶,其中POD是消除活性氧、酚類和胺類等氧化產(chǎn)物的關(guān)鍵酶[5]。目前動植物中已鑒定到的POD酶體包括辣根過氧化物酶(Horseradish peroxidase,HRP)、甲狀腺過氧化物酶(Thyroid peroxidase,TPO)、血紅素過氧化物酶(Heme peroxidase,HP)、木質(zhì)素過氧化物酶(Lignin peroxidase,LiP)、錳過氧化物酶(Manganeseperoxidase,MnP)和GPX等,其中GPX為動物、植物和真菌共有[20]。動物GPX分子結(jié)構(gòu)中一般含有1個硒代半胱氨酸,是酶分子中具有催化活性的作用位點;植物GPX與動物GPX序列高度相似,但是目前尚未發(fā)現(xiàn)含有硒的植物,其酶分子催化位點是半胱氨酸(Cystine,Cys)[21]。本研究通過基因克隆明確了LeGPX蛋白的氨基酸殘基序列,通過同源序列比對發(fā)現(xiàn)其酶分子中的催化位點(Cys)與植物相同(靈芝以及草菇GPX研究中存在相同結(jié)果),并且催化活性區(qū)(NTASA(K)CGFT)和標志基序(I(V)L(V)AFPCNQF)也與其他植物GPX相似[19,22]。前期研究發(fā)現(xiàn)多種食用菌中均能檢測到硒代半胱氨酸,結(jié)合香菇LeGPX催化位點特性,暗示其可能與植物GPX功能類似[23]。除此之外,本研究發(fā)現(xiàn)LeGPX蛋白不存在信號肽和跨膜結(jié)構(gòu)域,是定位于細胞質(zhì)的非分泌蛋白,該結(jié)果與多種植物GPX細胞定位情況相同[24]。
菌蓋表皮和菌肉褐變是香菇采后貯藏期間的主要問題之一,目前對于該類褐變發(fā)生的機制尚無系統(tǒng)研究。荔枝表皮、鴨梨果心以及雙孢蘑菇表皮采后褐變相關(guān)的研究中發(fā)現(xiàn),褐變發(fā)生初始階段機體的POD活性會迅速上升,褐變發(fā)生中期活性降低并保持平穩(wěn),褐變后期POD活性會隨褐變程度加深而逐步升高;POD可能通過催化谷胱甘肽或酚類物質(zhì)氧化引起呈色物質(zhì)積累,進而造成機體發(fā)生褐變[4,25-26]。本研究發(fā)現(xiàn)菌蓋和菌柄部位LeGPX基因在香菇發(fā)生褐變前期和末期均會出現(xiàn)顯著上調(diào)表達,暗示該基因可能是造成香菇采后褐變的因素之一。本研究還發(fā)現(xiàn),與菌蓋部位相比LeGPX基因在菌柄部位更為活躍。脫離栽培基質(zhì)后的菌柄可能承擔為彈射孢子提供能量的作用,相比菌蓋該部位代謝更為旺盛,由此造成的氧化損傷產(chǎn)物積累可能是引起香菇褐變發(fā)生的誘因。
本研究基于基因組數(shù)據(jù)鑒定并克隆了香菇LeGPX基因的編碼序列,通過生物信息學(xué)分析明確了LeGPX蛋白的生物學(xué)特性,通過構(gòu)建重組載體、原核表達和金屬離子親和純化獲得了LeGPX可溶蛋白,同時借助qRT-PCR方法明確了LeGPX基因在香菇褐變發(fā)生中的表達模式。以上結(jié)果為深入開展香菇采后褐變發(fā)生機制研究提供了基礎(chǔ)。
