低氧對非小細胞肺癌PDHA1、GBP1表達及細胞增殖的影響
2022-05-13 03:19:46曹靜楚天驕班振英張威智艷芳曾憲旭
中國老年學雜志 2022年9期
關鍵詞:肺癌
曹靜 楚天驕 班振英 張威 智艷芳 曾憲旭
(鄭州大學第三附屬醫院病理科,河南 鄭州 450052)
肺癌最常見的組織學類型為非小細胞肺癌(NSCLC),5年生存率僅40%左右。NSCLC的發病機制非常復雜,與多種分子異常、環境及遺傳因素密切相關〔1〕。低氧是腫瘤普遍存在的微環境,低氧與代謝重編程互為因果促進了腫瘤的進展。腫瘤細胞糖代謝的特點是優先選擇糖酵解,而回避氧化磷酸化的代謝模式。既往研究發現,抑制丙酮酸脫氫酶E1α亞單位(PDHA1)的表達能促進糖酵解〔2〕。 鳥苷酸結合蛋白(GBP)1是干擾素(IFN)-γ誘導下釋放出的活性物,主要作用是抵抗各種病原體入侵〔3〕。大多數關于GBP1的研究集中在炎癥疾病上,僅有少數研究證實GBP1與腫瘤相關。本研究擬分析低氧對NSCLC患者PDHA1、GBP1表達及細胞增殖的影響。
1 材料與方法
1.1實驗材料 選取20例配對的肺癌組織及癌旁組織,其中臨床分期Ⅰ、Ⅱ期(早期癌)12例,Ⅳ期(晚期癌)8例。肺鱗癌細胞株LC-42及肺腺癌細胞株SELS均來源于挪威奧斯陸大學腫瘤研究實驗室凍存株。PDHA1抗體為Cell Signaling Technology公司生產,GBP1抗體為Novus Biologicals公司生產。主要儀器:Xvivo Closed Incubation System由美國Bio Spherix公司制造,細胞培養時可以準確調節不同含氧量。
1.2方法 肺鱗癌LC-42及肺腺癌SELS細胞株各自分別在常氧及低氧條件下培養。常氧培養:CO2培養箱的設定條件是37℃、20%O2、5%CO2、75%N2,飽和濕度。低氧培養:常氧培養24 h,待細胞貼壁,轉移至Xvivo Closed Incubation System低氧培養48 h,即設定條件是37℃、1%O2、5%CO2、94%N2,飽和濕度。
1.3Western印跡 肺癌組織及細胞經過蛋白提取、蛋白測定及處理、聚丙烯酰胺凝膠電泳、轉膜、封閉,最后一抗及二抗孵育、曝光,實驗重復6次。
1.4RT-PCR 兩種細胞系分別常氧及低氧處理后,通過胰蛋白酶消化細胞并提取RNA。根據TaKaRa逆轉錄試劑盒步驟反轉錄合成cDNA。冰上配制Template DNA/cDNA 2 μl 、RNase-free ddH2O 10 μl,10倍緩沖液3.0 μl,Taq 0.2 μl,dNTP 2.4 μl,上下游引物各1.0 μl。PCR在下述反應條件下進行:94℃變性30 s,56℃淬火30 s,72℃延伸30 s,35個周期,最后72℃延伸10 min。根據2-△△Ct,測出mRNA相對值,實驗重復6次。
1.5CCK-8法檢測低氧對細胞增殖的影響 兩種細胞系分別常氧及低氧下培養48 h,調整細胞數為1×104/ml,每孔加100 μl細胞懸液,接種于96孔板中,在培養箱內常氧培養24、48、72 h,每孔加入CCK-8溶液10 μl繼續培養2 h,于酶標儀檢測450 nm處光密度(OD)值,細胞增殖率=(處理組OD值-空白OD值)/(對照組OD值-空白OD值)×100%。
引物序列:來源于NCBI數據庫,經Primer premier5.0軟件設計及BLAST檢測其特異性,由TAG Copenhagen A/S公司合成。PDHA1上游引物:5′-TTCTCAGAAGCCGGCAAGC-3′,177 bp,下游列物5′-ATTCGGCGTACAGTCTGCAT-3′;GBP1上游引物:5′-TGGAACGTGTGAAAGCTGAG-3′,326 bp,下游引物:5′-TGACAGGAA GGCTCTGGTC-3′;GAPDH上游引物:5′-TGCCTCCTGCACCACCAACT-3′,423 bp,下游引物:5′-CCCGTTCAGCTCAGGGATGA-3′。
1.6統計學方法 采用SPSS19.0軟件進行t檢驗和單因素方差分析。
2 結 果
2.1NSCLC及癌旁組織中PDHA1和GBP1蛋白表達比較 NSCLC臨床分期增高,PDHA1蛋白表達減少,而GBP1蛋白表達增加,差異有統計學意義(P<0.05),見圖1,表1。

圖1 NSCLC與癌旁組織中PDHA1和GBP1蛋白表達
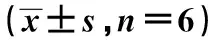
表1 NSCLC組織中PDHA1和GBP1蛋白表達
2.2低氧對肺鱗癌LC-42及肺腺癌SELS細胞系中PDHA1和GBP1蛋白表達的影響 與20%O2組相比,1%O2組PDHA1蛋白表達顯著降低,而GBP1蛋白表達顯著提高,差異有統計學意義(P<0.05)。見表2,圖2。
2.3低氧對肺鱗癌LC-42及肺腺癌SELS細胞系中PDHA1和GBP1 mRNA表達的影響 與20%O2組相比,1%O2組PDHA1 mRNA表達顯著降低,而GBP1 mRNA表達顯著提高,差異有統計學意義(P<0.05)。見表2。
表2 肺癌兩種細胞系PDHA1和GBP1蛋白及mRNA的表達
2.4CCK8法檢測低氧對細胞增殖的影響 與20%O2組相比,培養24及48 h,1%O2組增殖率顯著低于對照組(P<0.05),而培養72 h,兩種細胞系的增殖率顯著高于對照組(P<0.05)。見表3。

1~4:20%O2組LC-42、20%O2組SELS、1%O2組LC-42、1%O2組SELS圖2 低氧及常氧培養兩種肺癌細胞系PDHA1、GBP1蛋白表達

表3 肺癌兩種細胞系細胞增殖率比較
3 討 論
本課題組前期研究結果表明,低氧刺激前列腺癌及卵巢癌導致PDHA1表達降低,而GBP1表達增強,且提高了癌細胞干性及增殖能力。使用細胞周期抑制劑治療前列腺癌時,存活的前列腺癌細胞會轉向Warburg效應途徑(糖酵解),且伴有GBP1表達上調。而且,通過對NSCLC細胞系siRNA基因沉默PDHA1及GBP1進行相應研究,發現GBP1是PDHA1的下游基因。
目前,低氧對NSCLC患者PDHA1、GBP1表達及細胞增殖影響的報道仍少見。本研究結果提示,PDHA1低表達和GBP1過表達與NSCLC進展有關。將肺鱗癌LC-42及肺腺癌SELS細胞株給予低氧培養,導致兩種細胞系PDHA1蛋白及mRNA表達減少,而GBP1蛋白及mRNA表達增加。PDHA1及GBP1在體外低氧培養細胞的表達趨勢,與二者在NSCLC組織中的表達規律基本一致。CCK8法檢測低氧對兩種細胞增殖的影響表明,培養48 h以內,低氧處理細胞增殖緩慢,培養72 h則顯現出截然不同的結果,即低氧處理細胞增殖迅速。低氧雖然可以短暫抑制腫瘤增殖,但培養時間只要達到足夠長,則表現為細胞增殖能力增強。內在原因可能是低氧導致腫瘤細胞糖代謝選擇糖酵解的Warburg效應,而糖酵解是由PDHA1低表達觸發。GBP1作為PDHA1的下游基因,則表現為GBP1過表達促進了NSCLC進展。據此推斷,低氧、PDHA1低表達及GBP1高表達與NSCLC進展一脈相承,互為因果。
腫瘤細胞的社會微環境和物理微環境構成腫瘤的微環境。社會微環境是指腫瘤周圍的免疫細胞、成纖維細胞、內皮細胞及細胞外基質。物理微環境是指低氧、營養壓力、低pH值、氧化壓力〔4〕。腫瘤細胞會隨著微環境變化,代謝模式不斷做出適應性調整,這種特有的代謝變化稱為代謝重編程。因此,低氧是腫瘤細胞普遍存在的微環境要素之一。在低氧情況下,一些基因和蛋白的表達也會發生改變,甚至是參與了代謝重編程。腫瘤細胞吸收、利用最多的營養物質為葡萄糖,正常細胞攝取葡萄糖,經歷了線粒體的氧化磷酸化進行產能,而腫瘤細胞則選擇另外一種糖酵解的代謝方式。而氧化磷酸化與糖酵解轉換的關鍵節點就是PDHA1。有學者證明PDHA1下調會促進糖酵解代謝及腫瘤進展〔5〕。
最早研究發現腫瘤細胞可對葡萄糖進行高效吸收,且相應的吸收路徑為糖酵解,這種偏于糖酵解的代謝模式稱為Warburg效應〔6〕。葡萄糖進行糖酵解產生丙酮酸,丙酮酸會有兩種代謝途徑選擇:一種代謝途徑是線粒體膜上的丙酮酸轉運載體把丙酮酸從胞質運至線粒體基質,繼而線粒體基質內包含PDHA1的丙酮酸脫氫酶復合體催化丙酮酸和CoA生成乙酰-CoA和CO2,乙酰-CoA再進入三羧酸循環進行氧化磷酸化;另一種代謝途徑是丙酮酸在原來糖酵解的基礎上繼續代謝產生乳酸。因此,PDHA1是丙酮酸脫氫酶復合體的關鍵組件,連接著糖酵解和三羧酸循環,負責糖酵解和三羧酸循環之間的相互轉換。當PDHA1下調,丙酮酸脫氫酶復合體活性下降,乙酰-CoA生成減少,三羧酸循環障礙,糖代謝轉向糖酵解為主,Warburg效應增強,腫瘤更易侵襲及進展。而PDHA1表達正常時,糖代謝則以三羧酸循環氧化磷酸化為主〔7〕。有學者用miR-21-5p直接靶向PDHA1,抑制PDHA1表達,促進糖酵解和細胞增殖。miR-21-5p是一種MicroRNA,可綁定靶向RNA的3′端未翻譯區域,導致與目的mRNA分離,蛋白翻譯受阻〔8〕。PDHA1低表達預示卵巢癌、乳腺癌、大腸癌預后差〔9,10〕。GBP1為GTP酶的一種,被Ⅰ和Ⅱ型IFN誘導產生,對抗細菌、病毒及寄生蟲。其后相關研究不斷深入,且發現對細胞生長還具有調節作用,如通過對基質金屬蛋白酶(MMP)-1 、核轉錄因子(NF)-κB的調控實現加速細胞增殖〔11~13〕。Yamakita等〔14〕通過RT-PCR實驗顯示浸潤性肺腺癌GBP1表達的相對量是正常肺組織的3倍。用各種siRNA轉染RERF-LC-OK細胞,沉默GBP1基因,得出劃痕實驗顯著低于對照組。免疫組化顯示浸潤性肺腺癌的GBP1陽性率顯著高于正常肺組織,且GBP1表達與癌組織淋巴血管侵犯正相關〔14〕。已有實驗研究表明,GBP1可促進乳腺癌、膠質瘤、食管癌的形成,在癌細胞的生長中起一定調節作用。關于GBP1的潛在促腫瘤作用是目前此領域的研究重點〔15~17〕。本實驗結果與其研究結果是大致相同。
PDHA1及GBP1可通過信號通路進行調節,而影響了腫瘤細胞的代謝重編程,發揮調控細胞增殖及凋亡的功能。因此,在抗腫瘤治療中,可適當增強PDHA1及減弱GBP1的表達而抑制腫瘤生長。下一步應該研究其相關的信號通路及PDHA1、GBP1活性影響因素,從基因及物質能量代謝角度進行調控,從而有效預防NSCLC的發生,可將二者作為腫瘤治療的新靶點,更好為患者的精確靶向治療提供理論支持。
猜你喜歡
保健醫苑(2023年2期)2023-03-15 09:03:04
中國臨床醫學影像雜志(2022年2期)2022-05-25 13:24:34
中國藥學藥品知識倉庫(2022年1期)2022-03-23 04:16:57
昆明醫科大學學報(2021年4期)2021-07-23 01:21:44
天津醫科大學學報(2021年2期)2021-03-29 05:30:38
天津醫科大學學報(2019年6期)2019-08-13 07:04:26
癌變·畸變·突變(2016年3期)2016-02-27 06:15:34
醫學研究雜志(2015年12期)2015-06-10 06:57:46
中國當代醫藥(2015年7期)2015-03-01 02:01:19
鄭州大學學報(醫學版)(2015年1期)2015-02-27 14:50:26
