深圳灣海域赤潮生物演變及赤潮預警關鍵因子分析
2022-05-14 03:14:24李麗馬方方翟曉輝楊偉思葉鵬浩劉悅
生態科學 2022年3期
李麗, 馬方方, 翟曉輝, 楊偉思, 葉鵬浩, 劉悅,*
深圳灣海域赤潮生物演變及赤潮預警關鍵因子分析
李麗1, 馬方方2, 翟曉輝1, 楊偉思1, 葉鵬浩1, 劉悅1,*
1深圳市海洋發展研究促進中心, 深圳 518029 2清華蘇州環境創新研究院, 蘇州 215163
2020年5月初,深圳灣海域發生近5年來首次赤潮, 面積達到6 km2, 前期赤潮優勢種為中肋骨條藻(), 后期轉變為赤潮異彎藻()。布放于深圳灣海域的海洋環境綜合浮標自動監測到了赤潮發生過程的海水水質和生態數據變化: 赤潮發生前(4月底)硝酸鹽和磷酸鹽均大幅升高; 赤潮發生期間, 葉綠素濃度迅速升高, 最高值達到127.1 μg·L-1(5月2日), 海水溫度持續升高, 鹽度整體下降; 5月4日開始, 赤潮逐漸消散, 當日水溫日均值達到28 ℃以上, 風力也開始升高, 鹽度則繼續降低。經研究分析, 本次深圳灣赤潮的主要調控因素可能是水溫, 活性磷酸鹽大幅升高可能是引發中肋骨條藻大規模增殖的誘因, 而赤潮后期的陸源水體輸入導致的懸浮物增加導致了赤潮快速消亡。本次赤潮優勢種從硅藻到針胞藻的轉變, 主要原因可能是赤潮前期磷酸鹽快速消耗導致的磷供應不足。通過數據分析發現, 近十幾年來, 深圳灣海域赤潮生物逐漸從硅藻轉變為甲藻及其他藻類, 其原因與海域氮磷比持續升高、磷成為限制因子有關, 故海域赤潮監測預警應重點關注磷酸鹽的變化。
深圳灣; 磷酸鹽; 赤潮; 中肋骨條藻; 赤潮異彎藻
0 前言
深圳灣位于珠江口東側, 是一個半封閉感潮海灣, 整個海灣縱深約為14 km, 寬4.5—5.6 km, 平均水深2.9 m[1]。深圳灣海域入海河流眾多, 其中深圳河、新洲河、大沙河及元朗河直接排放入灣內。由于河流入海污染物通量較大, 深圳灣水質較差、富營養化程度較高, 近年來深港兩市均采取了一系列治理措施[2–4], 海域營養鹽自2005年開始整體呈下降態勢[5–6], 但由于影響海域污染的因素眾多, 根據2018年監測結果, 深圳灣海域3月、5月、8月、10月4次水質監測結果均為劣四類[7], 香港后海灣水質管制區2018年水質整體達標率為53%, 雖然較近10年均值有所提高, 但無機氮和磷酸鹽水平仍較高, 灣內無機氮和磷酸鹽濃度分別為2.02 mg·L-1和0.13 mg·L-1[8]。
深圳灣由于富營養化程度較高、且水體交換能力較弱, 有利于赤潮生物的生長和聚集。據不完全統計, 1980年至2019年, 深圳灣海域共記錄赤潮35次, 其中赤潮大多集中在2000年至2009年間, 2010年至2019年僅發生3次赤潮。在深圳的三灣一口(大亞灣、大鵬灣、深圳灣和珠江口)4個海域當中, 深圳灣赤潮次數相對較少, 但累計赤潮面積最大[9]。深圳灣的富營養化特征導致海域隨時有暴發赤潮的潛在風險[10], 其赤潮發生頻次低于營養鹽較低的深圳大鵬灣海域, 可能是由于陸源排污及潮流沖刷導致的水體懸浮物較多限制了水中浮游植物的光合作用和正常增殖[8], 隨著污染治理的加強, 海域透明度升高可能導致赤潮次數增多, 而營養鹽的降低則可能導致赤潮生物繼續演變。2020年5月初, 深圳灣暴發一次較大規模的赤潮, 赤潮原因種為中肋骨條藻()和赤潮異彎藻(), 這是2015年至今, 深圳灣首次發現赤潮, 本次赤潮暴發的生消原因是什么?深圳灣海域環境特征及赤潮生物是否均已發生變化?這些變化發生之后, 海域赤潮預警應該主要考慮哪些因素?近年來針對這方面的研究較少。本文將結合赤潮期間鄰近海域海洋環境浮標連續自動監測數據及赤潮生物的生理生態特征, 對本次赤潮的生消原因進行分析, 并進一步從深圳灣赤潮生物歷史演變及其水質變化情況, 分析海域赤潮的發生變化趨勢, 以期為深圳灣赤潮的預警提供科學支撐。
1 材料與方法
1.1 赤潮生物的采樣與分析
赤潮生物采集采用水采方式。取1 L表層樣品, 冷藏保存, 于采樣當天在萊卡倒置顯微鏡(LEICA DMi1)下用0.1 mL浮游植物計數框進行種類鑒定和計數。
1.2 浮標數據采集
本文主要數據來源于深圳灣海洋環境監測浮標的監測數據, 浮標的位置見圖1。該浮標搭載海水水質、氣象等傳感器。其中水質參數、氣象參數的監測頻率均為1 次·30 min-1; 營養鹽監測頻率為1 次·4 h-1。本文用于分析的環境參數包括水溫、鹽度、溶解氧、葉綠素、風速、硝酸鹽、活性磷酸鹽。浮標的水質監測傳感器為美國YSI公司的YSI6600型多參數水質監測儀, 水質傳感器海上運行期間均定期(每月1至2次)進行維護、校準和數據比對。
1.3 赤潮歷史數據來源
本文所提及的赤潮生物變化的數據來源于已公開發表的著作、文章及歷年的海洋環境狀況公報。其中, 1980年至2008年, 赤潮記錄主要來源于《中國典型海域赤潮災害發生規律》中深圳海域部分[11], 2009年至2017年赤潮記錄來源于廣東省海洋環境質量/狀況公報[12–13], 2018年至2019年赤潮記錄來源于廣東省生態環境狀況公報[14]。
2 結果與分析
2.1 2020年5月深圳灣赤潮發生過程
2020年4月29日至30日, 浮標自動監測數據顯示葉綠素和溶解氧含量較高, 提示海域有赤潮風險。2020年5月2日, 中心技術人員赴現場采樣, 鑒定結果顯示該海域赤潮生物優勢種為中肋骨條藻(); 5月3日, 監測結果發現赤潮優勢種轉變為赤潮異彎藻(), 赤潮累計面積約為6 km2。5月3日當天, 赤潮中心區域赤潮異彎藻的平均細胞密度為6.7×106個·L-1, 氨氮和硝酸鹽濃度分別為2.42 mg·L-1, 活性磷酸鹽濃度為0.026 mg·L-1; 5月4日, 現場監測發現赤潮消散。
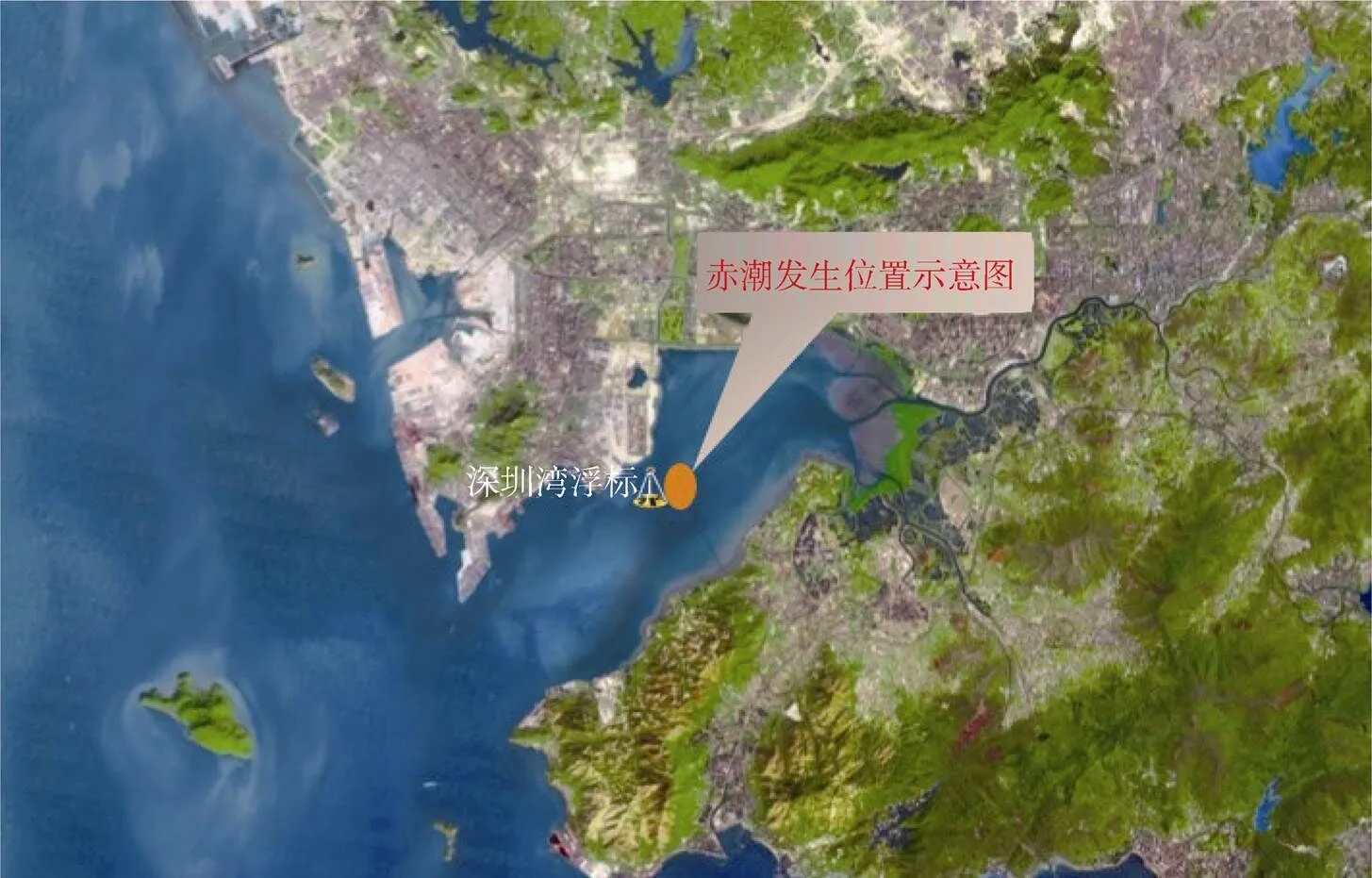
圖1 深圳灣浮標站位及赤潮發生位置示意圖
Figure 1 Buoy station and location of HAB in Shenzhen Bay
2.2 赤潮發生前后浮標數據變化
深圳灣浮標4月22日至5月5日海洋生態環境相關參數變化見圖2至圖5。
根據圖2中葉綠素變化情況, 將赤潮發生過程分為4個階段: 赤潮發生前(4月28日之前), 赤潮早期(4月29日至30日), 赤潮暴發期(5月1日至3日)和赤潮消亡期(5月4日至5日); 各階段水質參數特征值見表1。
赤潮發生前: 4月28日之前, 葉綠素整體較低且變化較小, 除偶有高值外, 大部分時間低于10 μg·L-1, 28日下午葉綠素升高。由于4月22日開始降溫, 水溫從26 ℃持續下降至22 ℃, 自26日開始, 水溫則又波動升高, 至28日水溫日均值升高至24.5 ℃; 鹽度受潮汐影響波動較大, 但日均值穩定在17—19之間。海域風速(圖4)則從23日開始保持穩定, 風力均小于3級。從圖5可知, 4月22日至26日, 營養鹽總體穩定, 硝酸鹽小幅降低、活性磷酸鹽則緩慢升高(浮標氨氮數據異常, 不納入分析), 27日開始, 硝酸鹽和活性磷酸鹽連續兩天大幅升高, 濃度分別升高1倍和1.5倍, 日均值分別達到0.799 mg·L-1和0.219 mg·L-1。
赤潮早期: 4月29日至4月30日, 葉綠素有所升高, 且變化幅度增大, 29日和30日葉綠素日均值分別為20.4 μg·L-1和26.8 μg·L-1, 峰值濃度則達到40 μg·L-1以上。溶解氧變化范圍也增大, 尤其4月30日, 葉綠素達到峰值時, 溶解氧最高值達到15 mg·L-1以上。這兩日水溫繼續升高, 4月30日水溫日均值為25.5 ℃, 鹽度日均值則整體高于上一階段, 且波動范圍較小。營養鹽也有較大變化, 活性磷酸鹽大幅降低, 降至0.05 mg·L-1, 硝酸鹽也有降低, 但降低幅度相對較小。
赤潮暴發期: 從5月1日葉綠素濃度變化可以發現, 明顯進入赤潮暴發期。5月1日至3日, 葉綠素日均值在36.9—40.5 μg·L-1之間, 其中5月2日濃度最高, 當日最高值為127.1 μg·L-1; 溶解氧與葉綠素變化趨勢保持一致, 日均值在11.78— 12.79 mg·L-1之間, 最高值則出現在5月1日, 可能與當日赤潮生物生長速率最快有關。水溫緩慢升高, 日均值從26.0 ℃升高至27.3 ℃, 風速均較小, 風力保持在3級以下, 鹽度則大幅降低, 5月1日鹽度日均值較前一日下降1.2, 至5月3日, 鹽度日均值又下降了3左右, 表明陸源輸入突然增加。5月1日活性磷酸鹽濃度較前一日降低一半, 之后保持穩定, 其中5月3日浮標磷酸鹽日均值為0.022 mg·L-1, 與現場采樣監測結果(0.026 mg·L-1)較為一致; 硝酸鹽在5月1日和2日大幅下降, 降低了30%左右, 5月3日又有所升高。
赤潮消亡期: 5月4日, 葉綠素突然降低, 峰值濃度降為22.7 μg·L-1, 日均值降至8 μg·L-1以下, 且5月5日繼續降低, 溶解氧日均值也隨之大幅降低, 5月5日日均值已降至7 mg·L-1。現場監測表明赤潮已消退。水溫、鹽度延續之前的變化, 水溫繼續升高, 日均值達到28 ℃以上, 鹽度繼續下降, 風力則有所增強, 5月5日全天大部分時間風力高于3級。硝酸鹽和活性磷酸鹽濃度保持穩定。
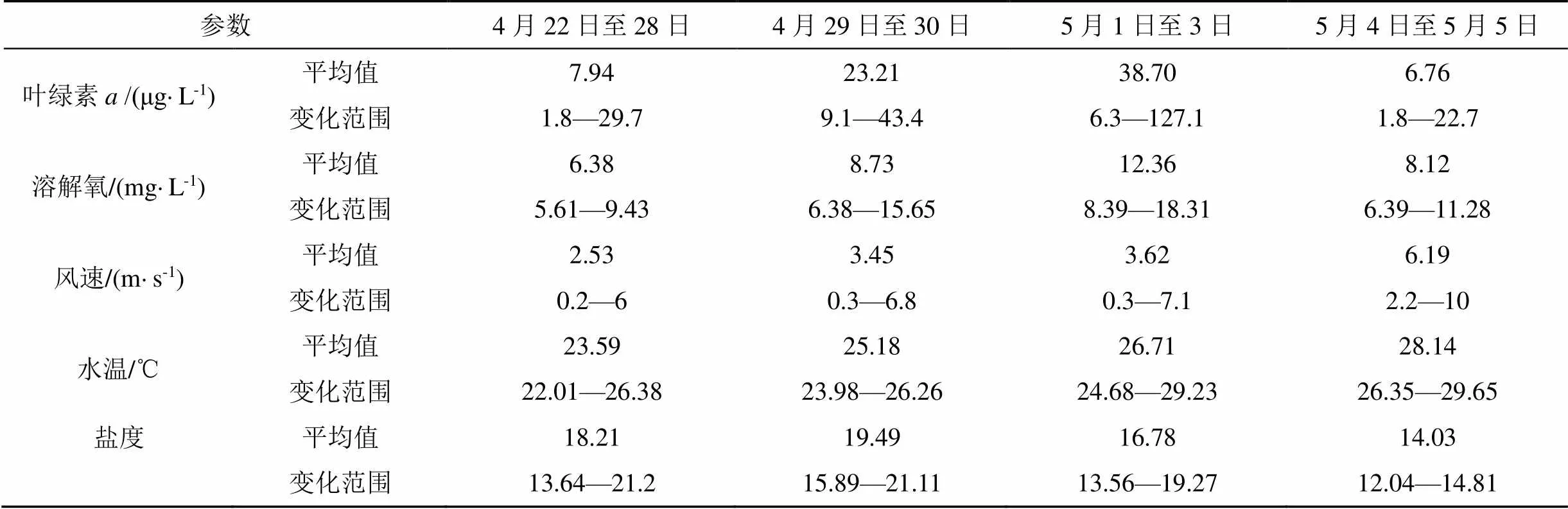
表1 赤潮各階段浮標主要參數平均值及變化范圍
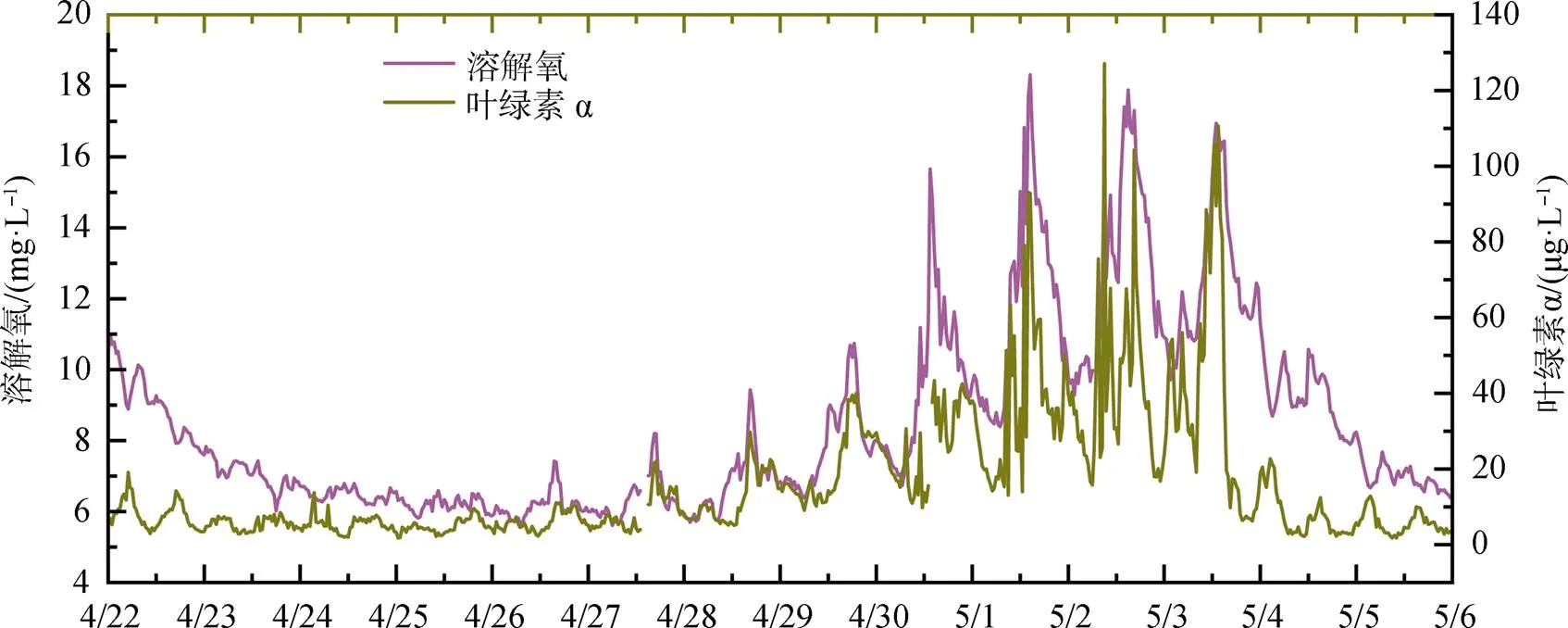
圖2 2020年4月22日至5月5日深圳灣浮標葉綠素a、溶解氧變化
Figure 2 Concentrations of chlorophylland dissolved oxygen in Shenzhen Bay during Apr. 22th and May. 5th
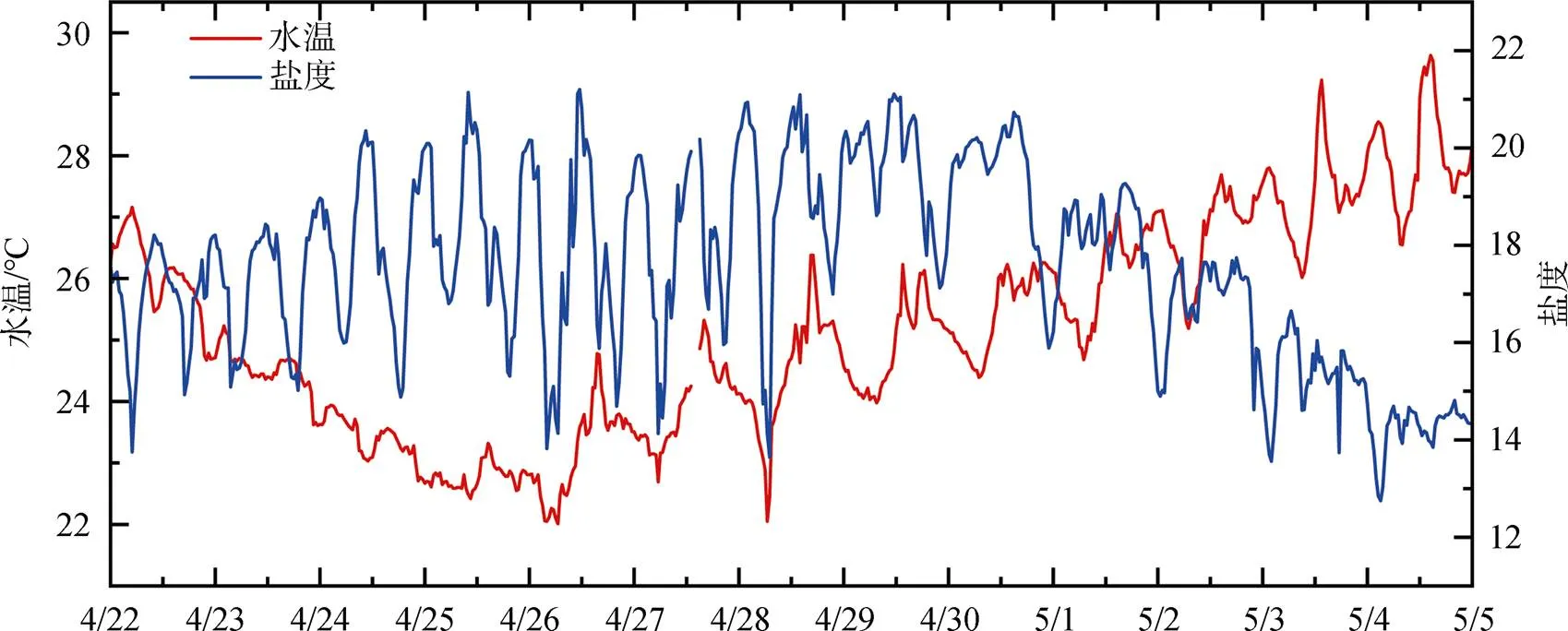
圖3 2020年4月22日至5月5日深圳灣浮標水溫、鹽度變化
Figure 3 Variations of water temperature and salinity in Shenzhen Bay during Apr. 22th and May.5th
3 討論
3.1 本次赤潮生消原因分析
赤潮發生是海洋中多種環境因素綜合作用所致, 春季由于水溫升高至20 ℃以上, 為赤潮高發期, 且深圳灣海域水體交換較差, 營養鹽濃度整體較高, 具備暴發赤潮的基礎條件。以下主要結合本次赤潮的水文氣象因子和營養鹽變化, 對赤潮生消原因進行分析。
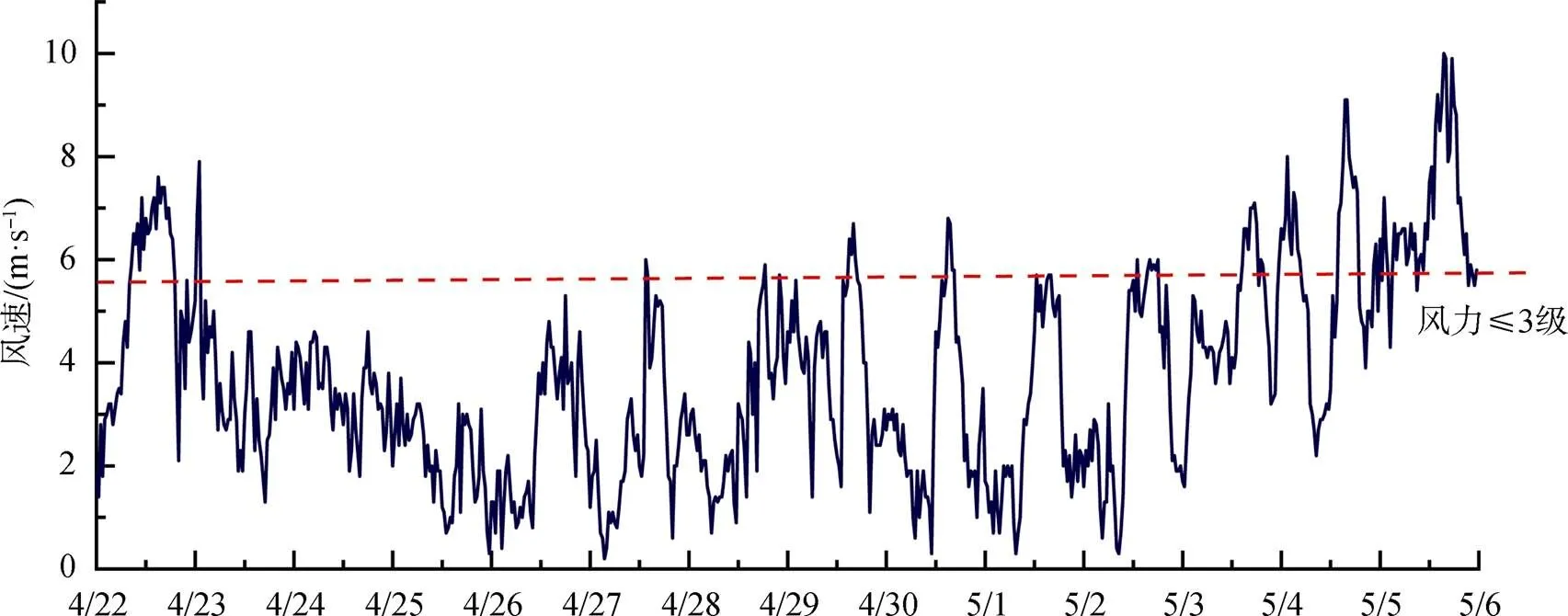
圖4 2020年4月22日至5月5日深圳灣浮標風速變化
Figure 4 Wind speed variations in Shenzhen Bay during Apr. 22th and May. 5th
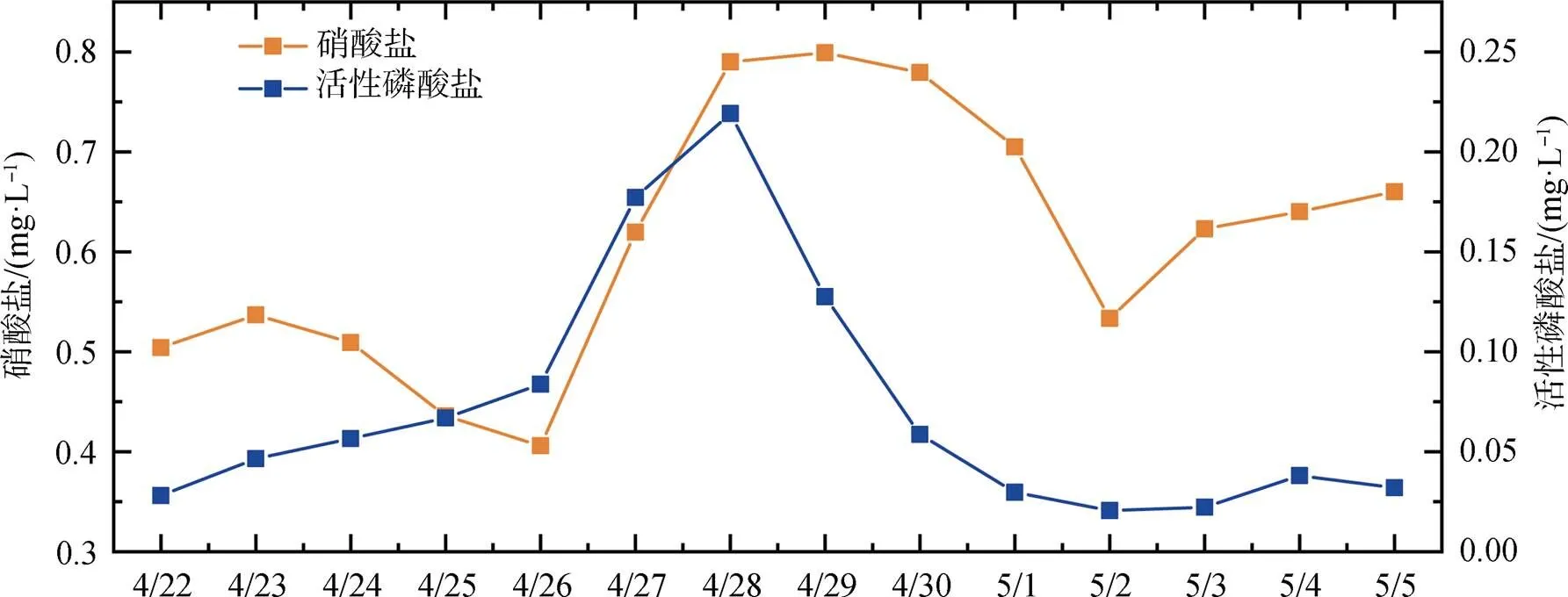
圖5 2020年4月22日至5月5日深圳灣硝酸鹽、活性磷酸鹽日均值變化
Figure 5 Daily average variations of nitrate and phosphate concentration in Shenzhen Bay during Apr. 22th and May. 5th
(1) 從第2.2部分數據分析中可以發現, 赤潮發生前水溫約為22 ℃, 并持續緩慢升高, 至中肋骨條藻赤潮發生期水溫升高至26 ℃。研究表明, 中肋骨條藻是一種廣溫廣鹽性藻類, 但其最適增殖溫度為24—28 ℃, 在其適溫范圍內, 水溫升高有利于中肋骨條藻的快速增殖[15]。赤潮異彎藻的適溫范圍同樣較廣, 25 ℃是其最適生長溫度, 但在10—30 ℃之間均能正常生長[16–17]。5月3日, 赤潮優勢種轉變為赤潮異彎藻, 當日平均水溫為27.35 ℃, 在赤潮異彎藻正常生長溫度范圍內。相關性分析結果表明, 葉綠素與水溫、溶解氧呈顯著正相關關系(表2), 溶解氧與葉綠素a顯著正相關, 與赤潮期間浮游植物光合作用過程中釋放大量氧氣有關, 因此, 水溫可能是本次赤潮的主要調控因素。
(2)由于本次赤潮原因種均可以適應大幅度的鹽度變化, 實際上中肋骨條藻更喜低鹽[18], 故而鹽度下降本身對本次赤潮生消影響不大, 但鹽度降低表明陸源輸入增加。4月30日之前, 鹽度每日呈現周期性變化, 主要為潮汐影響所致; 4月30日開始鹽度整體呈下降趨勢, 表明持續有陸源淡水輸入, 而根據5月3日現場調查數據, 當日赤潮中心區氨氮高達2.42 mg·L-1, 推測為近期陸源污染物大量排入灣內所致。前期陸源污染輸入導致的營養鹽增加可能對赤潮生物生長起到了一定的促進作用, 而陸源污染的持續輸入, 可能導致海域懸浮物增加、透明度下降, 進而影響赤潮生物的光合作用, 這也可能是5月4日赤潮快速消亡的原因之一。

表2 水溫、鹽度、溶解氧、風速與葉綠素a的相關性
注:**. 表示<0.01。
(3)赤潮發生前活性磷酸鹽大幅升高可能是引發中肋骨條藻大規模增殖的誘因。研究顯示, 中肋骨條藻屬于營養型浮游植物, 氮磷營養鹽的添加會極大地促進藻細胞的增殖[19]。根據浮標監測數據, 4月26日至28日硝酸鹽和活性磷酸鹽濃度升高一倍左右, 尤其是活性磷酸鹽, 其在4月22日濃度較低, 低于0.03 mg·L-1, 為貧營養狀態, 之后開始緩慢增加, 至4月26日日均值達0.05 mg·L-1, 達到富營養狀態, 之后的繼續升高為中肋骨條藻快速增殖提供了較為充足的營養基礎。由于赤潮發生前浮標氨氮數據異常, 無法判定無機氮整體水平, 但硝酸鹽日均值均在0.4 mg·L-1以上, 已經達到富營養的狀態, 結合第(2)點中陸源污染輸入, 表明海域無機氮較為充足, 不存在N限制, 硝酸鹽的升高對赤潮發生起到了促進作用, 但并非直接誘因。
3.2 從本次赤潮優勢種變化分析海域長期赤潮生物演變
根據深圳灣赤潮歷史記錄[11–14], 上世紀80、90年代赤潮生物主要為甲藻, 2000年之后演變為硅藻占優勢, 而2005年開始, 甲藻及其他種類引發赤潮次數有所升高, 硅藻藻華開始減少(表3)。赤潮生物的演變與環境因子變化密切相關, 本次赤潮中優勢種及營養鹽變化, 是海域長期環境變化的結果, 可為海域赤潮生物演變的原因提供一些數據支撐。
從現場5月2日和5月3日的監測數據得知, 赤潮前期優勢種為中肋骨條藻, 后期優勢種轉變為赤潮異彎藻。根據中肋骨條藻的高速增殖特性[20–22]及赤潮前期磷酸鹽快速降低, 也可確認本次赤潮生物經歷了以硅藻為主到以針胞藻為主的變化。由于中肋骨條藻生長較快, 其藻華通常可以快速暴發, 故而在富營養化嚴重的海域, 其經常作為優勢種首先暴發赤潮, 同時由于生長速率較快, 中肋骨條藻等小型硅藻對N、P、Si的營養需求均較高[23–24], 在消耗掉一定比例的營養鹽后, 導致赤潮后期優勢種轉變為赤潮異彎藻。而4月28日至5月1日活性磷酸鹽持續大幅降低可能是中肋骨條藻增殖逐漸加快大量消耗所致, 由于海域以無機磷為主[6], 活性磷酸鹽下降至0.03 mg·L-1之后, 其增殖受到嚴重限制; 而前期生長較慢的赤潮異彎藻則有多種營養策略(利用有機營養鹽、混合營養等)[25–26], 并可以通過化感作用抑制中肋骨條藻生長[27], 故而其在低營養鹽的條件下也可以保持正常的增殖速度。
深圳灣海域營養鹽濃度、比例和成分的變化可能導致了赤潮生物的演變, 體現在本次赤潮中導致了優勢種的變化, 體現在長時間周期中, 則導致了海域赤潮生物的演變。深圳灣上世紀80年代至2005年前后赤潮生物從甲藻為主逐步演變為硅藻為主, 與深圳灣海域營養鹽濃度持續升高有關, 研究顯示80年代至2005年前后, 深圳灣營養鹽為波動升高的趨勢[9,28–29], 這與深圳的飛速發展和人口快速增長密切相關, 而營養鹽充足的海域硅藻更容易成為優勢種而引發赤潮。2005年開始, 營養鹽則整體為下降趨勢[5–8], 部分年份為磷中等限制潛在性富營養狀態[9]。有研究表明, 近幾十年來中國近海氮輸入量不斷增加, 而磷輸入量不斷減少, 磷限制趨勢日益嚴重[30], 臨近深圳灣的珠江口海域也是如此[31]。深圳灣海域磷酸鹽雖整體較高, 但近年來氮磷比持續升高[9], 相對來說磷成為限制因子, 本次赤潮中也體現出了這一點。營養鹽限制的環境可能有利于甲藻、針胞藻等采取K競爭策略的藻類[21,32], 除此之外, 營養鹽限制的條件下化感作用更加明顯[33–36], 例如, 錐狀斯氏藻、海洋原甲藻、赤潮異彎藻均能釋放出化感物質, 并對中肋骨條藻生長產生抑制作用。
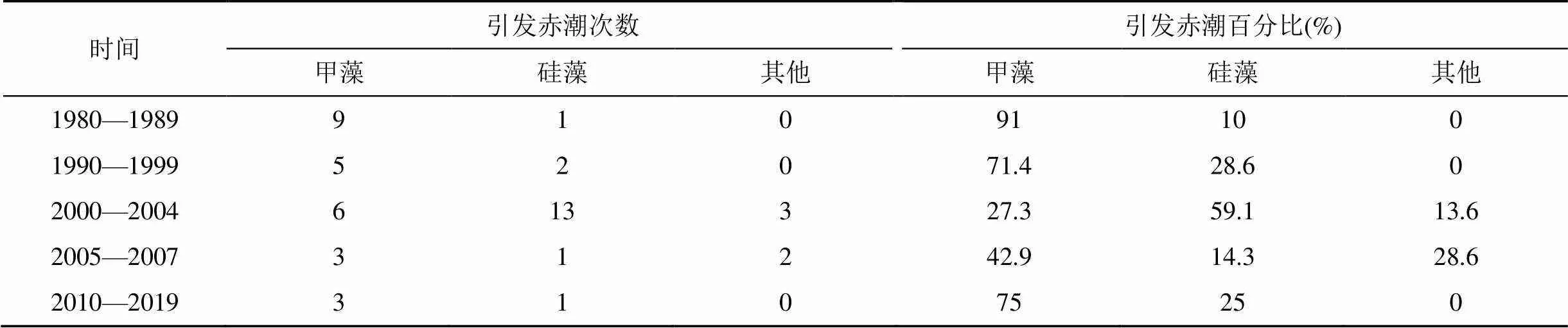
表3 1980—2019年深圳灣赤潮生物引發赤潮次數統計
3.3 深圳灣海域赤潮預警關鍵因子分析
影響赤潮的主要因素包括水溫、風力、流場、降雨、透明度(影響表層浮游植物光合作用)、營養鹽等; 深圳灣灣內由于海流較弱[37], 且除冬季部分時間水溫較低以外, 其他大部分時間水溫均適宜藻類增殖, 故應重點關注降雨、光照和營養鹽三個因素。降雨對赤潮具有雙重作用, 營養鹽較低的海域, 降雨后導致的陸源營養鹽輸入常常是赤潮暴發的誘因, 而降雨也會導致赤潮消散, 對深圳灣海域而言, 第一種情況較少發生, 由于海域營養鹽較高, 降雨的稀釋作用反而會導致營養鹽降低, 海域豐水期營養鹽相對較低[5–6, 10]即為印證, 因此深圳灣海域只需關注降雨對赤潮的抑制作用。由于陸源污染輸入對海域透明度的影響, 以及海域水深較淺, 潮流的沖刷作用也會導致水體垂直混合加劇, 進而影響表層透明度, 這些均會影響浮游植物對光的吸收, 是影響深圳灣赤潮的一項關鍵因素, 而隨著深圳灣污染治理的加強, 海域透明度整體升高可能會導致赤潮增加。深圳灣海域氮較為充足, 磷則可能成為限制因素, 故而成為深圳灣赤潮預警的另一項關鍵指標, 在其他因素均適宜的條件下, 如果同時監測到磷酸鹽和葉綠素大幅升高, 則海域發生赤潮的幾率極高, 在后續的赤潮預警和監測中可以進行持續驗證。
4 結論
(1)本次深圳灣赤潮初期第一優勢種為中肋骨條藻, 后期轉變為赤潮異彎藻; 水溫可能是本次赤潮的主要調控因素, 活性磷酸鹽大幅升高可能是本次赤潮暴發的主要誘因, 陸源輸入導致的水體懸浮物增加可能是導致赤潮快速消亡的原因。
(2)深圳灣海域赤潮生物近十幾年來硅藻藻華減少、甲藻及其他藻類赤潮增多, 可能是由于海域氮磷比升高、磷成為限制因子所致。
(3)隨著深圳灣污染治理工作的推進, 海域透明度升高可能會導致赤潮頻次增加, 深圳灣海域赤潮預警時可以關注透明度、磷酸鹽, 重點關注磷酸鹽的變化。
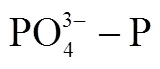
[2] 胡嘉東, 秦華鵬. 深圳河灣水系生態需水的污水資源化[J]. 水資源保護, 2008, 24(3): 20–23.
[3] 劉寧, 呂銳鋒. 深圳河灣水污染水環境治理[M]. 北京:中國水利水電出版社, 2006: 1–30.
[4] 李威, 鄺璐, 劉宇航, 等. 深圳灣海域水環境治理現狀及展望[J]. 中國給水排水, 2016, 32(18): 29–31.
[5] 李緒錄, 張軍曉, 史華明, 等. 深圳灣及鄰近沿岸水域總溶解氮的分布、組成和來源及氮形態的轉化[J]. 環境科學學報, 2014, 34(8): 2027–2034.
[6] 張軍曉, 李緒錄, 肖志建. 深圳灣及鄰近水域總溶解磷的來源和時空分布[J]. 中國環境科學, 2015, 35(3): 871–878.
[7] 深圳市海洋漁業局. 深圳市海洋環境狀況公報[R]. 深圳: 深圳市海洋漁業局, 2018: 8–11.
[8] 香港特別行政區環境保護署. 香港海水水質報告[R]. 香港: 香港特別行政區環境保護署, 2018: 21–23.
[9] 桓清柳, 龐仁松, 周秋伶, 等. 深圳近岸海域氮、磷營養鹽變化趨勢及其與赤潮發生的關系[J]. 海洋環境科學, 2016, 35(6): 908–914.
[10] 張靜, 張瑜斌, 周凱, 等. 深圳灣海域營養鹽的時空分布及潛在性富營養化程度評價[J]. 生態環境學報, 2010, 19(2): 253–261.
[11] 趙東至等. 中國典型海域赤潮災害發生規律[M]. 北京:海洋出版社, 2010: 269–273.
[12] 廣東省海洋與漁業局. 廣東省海洋環境質量公報(2009, 2010) [EB/OL]. http://gdee.gd.gov.cn/hjzkgb/index_2.html.
[13] 廣東省海洋與漁業局. 廣東省海洋環境狀況公報(2011, 2012, 2013, 2014, 2015, 2016, 2017) [EB/OL]. http://gdee. gd.gov.cn/hjzkgb/index_2.html .
[14] 廣東省生態環境廳. 廣東省生態環境狀況公報(2018, 2019)[EB/OL].http://gdee.gd.gov.cn/hjzkgb/ index_2.html.
[15] 霍文毅, 俞志明, 鄒景忠, 等. 膠州灣中肋骨條藻赤潮與環境因子的關系[J]. 海洋與湖沼, 2001, 32(3): 311–318.
[16] 顏天, 周名江, 錢培元. 赤潮異彎藻的生長特性[J]. 海洋與湖沼, 2001, 33(2): 209–214.
[17] 王燕, 宋洪軍, 李艷, 等. 溫度對赤潮異彎藻生長速率及細胞體積和生化組成影響的研究[J]. 海洋與湖沼, 2015, 45(1): 41–46.
[18] 李金濤, 趙衛紅, 楊登峰, 等. 長江口海水鹽度和懸浮體對中肋骨條藻生長的影響[J]. 海洋科學, 2005, 29(1): 34–37.
[19] 王金花, 唐洪杰, 楊茹君, 等. 氮磷營養鹽對中肋骨條藻生長及硝酸還原酶活性的影響[J]. 海洋科學, 2008, 32(12): 64–68.
[20] 陳炳章, 王宗靈, 朱明遠, 等. 溫度、鹽度對具齒原甲藻生長的影響及其與中肋骨條藻的比較[J]. 海洋科學進展, 2005, 23(1): 60–64.
[21] 葛蔚, 汪芳, 柴超. 氮和磷濃度對中肋骨條藻和錐狀斯氏藻種間競爭的影響[J]. 植物生態學報, 2012, 36(7): 697–704.
[22] 孫科, 丘仲鋒, 何宜軍, 等. 東海原甲藻與中肋骨條藻的種間競爭數值模擬[J]. 生態學報, 2014, 32(23): 6898– 6909.
[23] RIEGMAN R. Nutrient-related selection mechanisms in marine phytoplankton communities and the impact of eutrophication on the planktonic food web[J]. Water Science and Technology, 1995, 32(4): 63–75.
[24] GLIBERT P M, BURKHOLDER J M. Harmful algal blooms and eutrophication: “strategies” for nutrient uptake and growth outside the Redfield comfort zone[J]. Chinese Journal of Oceanology and Limnology, 2011, 29(4): 724–738.
[25] KUDELA R M, LANE J Q, COCHLAN W P. The potential role of anthropogenically derived nitrogen in the growth of harmful algae in California, USA[J]. Harmful Algae, 2008, 8(1), doi: 10.1016/j.hal.2008.08.019.
[26] HERNDON J, COCHLAN W P. Nitrogen utilization by the raphidophyte: Growth and uptake kinetics in laboratory cultures[J]. Harmful Algae, 2007, 6(2): 260–270.
[27] HONJO T. Overview on bloom dynamics and physio- logical ecology of[M]∥SMAYDA T J, SHIMIZU Y. Toxic Phytoplankton bloom in the sea. Amsterdam: Elsevier Science Publishers, 1993: 33–41.
[28] 孫金水, WAI O W , 王偉, 等. 深圳灣海域氮磷營養鹽變化及富營養化特征[J]. 北京大學學報(自然科學版), 2010, 46(6): 960–964.
[29] 王越興. 深圳灣水環境質量及變化趨勢分析[J]. 環境科學導刊, 2011, 30(3): 94–96.
[30] 金杰, 劉素美. 海洋浮游植物對磷的響應研究進展[J]. 地球科學進展, 2013, 28(2): 253–261.
[31] YIN Kedong, HARRISON P J, BROOM M, et al. Ratio of nitrogen to phosphorus in the Pearl River and effects on the estuarine coastal waters: Nutrient management strategy in Hong Kong[J]. Physics and Chemistry of the Earth Parts A/B/C, 2011, 36(9/11): 411–419.
[32] 呂頌輝, 李英. 我國東海4種赤潮藻的細胞氮磷營養儲存能力對比[J]. 過程工程學報, 2006, 6(3): 439–444.
[33] JI Xiaoqing, HAN Xiaotian, ZHEN Li, et al. Allelopathic interactions betweenandorin laboratory Cultures[J]. Chinese Journal of Oceanology and Limnology, 2011, 29 (4): 840–848.
[34] FLYNN K, FRANCO J M, FERNáNDEZ P, et al. Changes in toxin content,biomass and pigments of the dinoflagellateduring nitrogen refeeding and growth into nitrogen and phosphorus stress[J]. Marine Ecology Progress Series, 1994, 111: 99–109.
[35] GRANéLI E, JOHANSSON N. Increase in the production of allelopathic substances bycells grown under N- or P-deficient conditions[J]. Harmful Algae, 2003, 2(2): 135–145.
[36] YAN Jiaying, LIU Jingya, CAI Yu, et al. Allelopathic effects and mechanisms ofisolated from the Pearl River Estuary[J]. Journal of Applied Phycology, 2019, 31: 2957–2967.
[37] 張靜, 孫省利, 林建國, 等. 深圳灣海域環境容量及污染總量控制研究——I.潮汐、潮流數值模擬[J]. 海洋通報, 2010, 29(1): 22–28.
Analysis on the key factors for the population evolution and early warning of harmful algal blooms based on an algal bloom in Shenzhen bay, the South China Sea
LI Li1, MA Fangfang2, ZHAI Xiaohui1, YANG Weisi1, YE Penghao1,LIU Yue1,*
1. Marine Development Research and Promotion Center of Shenzhen, Shenzhen 518067, China 2. Research Institute for Environmental Innovation (Suzhou) Tsinghua, Suzhou 215163, China
A harmful algae bloom was recorded in Shenzhen Bay, the South China Sea, in May 2020. The causative specie wasin the first beginning, then shifted to. The data from the water quality buoy in the area showed that concentrations of nitrate and phosphate increased significantly before the outbreak of the bloom. During the bloom, the concentration of chlorophyllreached its maximum value to 127.1 μg·L-1on May 2, accompanied by a continuously rising temperature and a decreasing salinity. In the process of the bloom disintegration, water temperatures kept increasing to as high as 28 ℃, whileas the salinity exhibited a continuous decrease. Analytic results indicated that temperature might be the dominant factor for the HAB.The rapid increase of phosphate imput might be the causative factor for thebloom, whileas the increases of suspended matters from riverine input promoted the rapid decline of the bloom. The shift of causative species from diatom to raphidophyte during the bloom was speculated to the consumption of large amount of phosphate during the HAB. The results from a long term data analysis showed that HAB causative species shift from diatoms to dinoflagellates and other taxas in the last decadewas due to continuously increase of N/P ratio. Phosphorus is a limited nutrient in the area. Authors suggest that monitoring of phosphate is of the first importance for the early warning of HABs in Shenzhen Bay.
Shenzhen Bay; phosphate; harmful algae bloom;;
李麗, 馬方方, 翟曉輝, 等. 深圳灣海域赤潮生物演變及赤潮預警關鍵因子分析[J]. 生態科學, 2022, 41(3): 82–89.
LI Li, MA Fangfang, ZHAI Xiaohui, et al. Analysis on the key factors for the population evolution and early warning of harmful algal blooms based on an algal bloom in Shenzhen bay, the South China Sea[J]. Ecological Science, 2022, 41(3): 82–89.
10.14108/j.cnki.1008-8873.2022.03.009
X55
A
1008-8873(2022)03-082-08
2021-04-25;
2021-07-03
國家自然科學基金項目(41606176)
李麗(1983—), 女, 河南信陽人, 博士, 助理研究員, 海洋生態及資源監測, E-mail: wuyoulily@126.com
劉悅(1965—), 女, 高級工程師, 海洋生態及資源監測, E-mail: liuyue@pnr.sz.gov.cn
