海拔梯度變化對黃緣蜾蠃筑巢結構及相關生物學特性的影響
2022-05-20 09:47:00竇飛越吳智宏林木青束祖飛李紅英寇若玫周澤揚黃敦元肖治術
生態學報 2022年8期
竇飛越,張 歡,吳智宏,林木青,束祖飛,李紅英,寇若玫,周澤揚,黃敦元,*,肖治術
1 重慶師范大學生命科學學院,媒介昆蟲重點實驗室,重慶 401331 2 廣東車八嶺國家級自然保護區管理局,韶關 512500 3 中國科學院動物研究所,農業蟲害鼠害綜合治理研究國家重點實驗室,北京 100101 4 中國科學院大學,北京 100049
地上筑巢的野生獨棲蜂類(包括蜜蜂和胡蜂類群)喜歡選擇天然存在的空穴材料如石縫、植物莖稈、枯木中的蟲洞以及蝸牛殼等場地進行筑巢[1—2]。研究發現適宜的棲息地環境、充足的食物資源以及適合的筑巢材料是這類昆蟲繁殖的主要限制因素[3—4]。因此,對獨棲蜂類而言,在資源有限的自然環境下,如何最佳利用筑巢資源以提高種群對環境的適應性是非常重要的。研究表明,在具有雌雄體型性二型現象的物種中,親本可以根據現有的筑巢資源豐度調整對子代的投資及性別分配比例[5—6]。例如,在Osmiarufa筑巢期中,由于雌性對后代提供的食物效率下降,導致其后代體型的減少和后代性比雄性偏多[7]。然而到目前為止,這類研究通常關注的更多的是水平帶上的差異[8—9],對于同一個地區不同海拔梯度下的獨棲蜂類是如何調整其筑巢策略以適應環境變化,還需要進一步研究[10]。
山地生態類環境系統是地球表面具有顯著起伏和坡度的三維地貌體[11]。海拔梯度作為山地生境差異的主導因子,其現象表現為溫度、降水、風速、光照及土壤等自然因子隨海拔變化而出現規律性的梯度變化,進而影響物種的多樣性及組成和分布[12—13]。目前的研究表明,不同類群或不同區域下的物種多樣性隨海拔梯度的分布格局各異[14],但大致呈現4種格局:即單調遞減格局(decreasing patterns)、低平臺高原格局(low-elevation plateaus,LP)、偏峰格局(low-elevation plateaus with a mid-elevation peak,LPMP)和中峰格局(mid-elevation peak,MP),4種格局分別占25%、15%、14%、45%[15]。除此之外,研究物種的多樣性分布格局及組成結構隨海拔梯度的變化還有助于理解全球氣候變化對物種的影響,同時為生物多樣性的保護提供理論依據[16—20]。在昆蟲中,捕食性和寄生性昆蟲在調節獵物和宿主的種群動態中起著至關重要的作用[21],其中野生蜂類是研究沿海拔梯度變化的模式生物之一[22]。
蜾蠃亞科(Eumeninae)隸屬于膜翅目(Hymenoptera)細腰亞目(Apocrita)針尾部(Aculeata)胡蜂科(Vespidae)[23—24],全世界共有205屬3773種,中國紀錄51屬267種[25]。研究表明,蜾蠃亞科多為獨棲類群,喜營自由生活,雌蜂交配后開始筑巢狩獵并產卵繁殖,其巢穴類型主要有兩種:一種是用泥土修建成中空壺狀的巢穴,另一種是利用天然空穴修建的巢穴[26—27]。該亞科的狩獵方式是雌蜂用蜇針蜇刺鱗翅目幼蟲,使獵物麻痹,再帶回巢穴中儲備以供后代取食。研究發現,黃唇蜾蠃(Rhynchiumbruneum)的一個巢穴內多達45條鱗翅目害蟲,因此蜾蠃類群對農林害蟲防治能起到一定控制作用[28—29]。此外,蜾蠃類群的蜂毒因具有麻痹和貯存活體寄主的功能,所以在蜂毒成分研究和新型麻醉劑的開發應用方面也具有一定的意義[30—32]。
黃緣蜾蠃(Anterhynchiumflavomarginatum)隸屬于蜾蠃亞科(Eumeninae)緣蜾蠃屬(Anterhynchium)[25],主要分布在我國浙江、上海、江西、湖南、四川、福建、廣西、云南等地[33—35]。研究表明,該蜂自然條件下選擇甲蟲廢棄的地道、樹木、竹子以及蘆葦等材料中筑巢,主要以鱗翅目幼蟲為食,是一種重要的捕食性天敵昆蟲[9]。
人工巢穴技術(trap-nesting)是利用蘆葦、紙筒、竹子等材料制成一端開口一端封閉的巢管,或直接在木頭上鉆蛀不同大小的巢孔并放置在野外用以收集獨棲蜂類的一種誘集裝置[36—39]。1967年,Karl首次利用巢管收集獨棲性蜂類[40],隨著該技術的推廣和完善,目前人工巢穴技術常和網掃、黃盤誘集、馬來氏網、飛行阻截等采集方法一起對研究區域進行標本采集并作為昆蟲多樣性分析的依據[41—42],也廣泛用于研究獨棲蜂類后代的性比及性別分配機制[5,9,43],還用于傳粉昆蟲和捕食性天敵的放養等[44—46]。本研究利用蘆葦制作的人工巢管,在廣東車八嶺國家級自然保護區中誘集不同海拔梯度下的黃緣蜾蠃,研究海拔梯度對該蜂筑巢策略的影響,旨在進一步探討黃緣蜾蠃等獨棲性蜂類對不同海拔梯度下特定生境的響應。
1 材料與方法1.1 研究地概況
廣東車八嶺國家級自然保護區位于中國廣東省始興縣東南部,北緯24°40′29″—24°46′21″,東經114°07′39″—114°16′46″,總面積75.45 km2。該區地勢西北高東南低,最低處樟棟水海拔330 m,最高峰天平架海拔1256 m。保護區年均溫度19.6 ℃,最高溫38.4 ℃,年降水量1,150—2,126 mm。保護區植物區系屬于南亞熱帶向中亞熱帶的過度地帶,是南嶺山脈南緣具有代表性的中亞熱帶常綠闊葉林[47]。
1.2 樣地設置
本研究自2017年底在廣東車八嶺國家級自然保護區布設人工巢箱對區內野生蜂類多樣性進行監測。人工巢箱布設方案:以整個保護區全境及周邊為單位制作1 km × 1 km網格地圖,共設置100個調查網格,每個調查網格布設3個人工巢箱(其中1個在林內,2個在林緣),每個巢箱有2個PVC管組成(PVC規格:長25 cm,內徑110 cm),每個PVC管內放有約70根巢管(巢管規格:長度在8—20 cm之間,內徑在0.5—1.7 cm之間),共300個巢箱(圖1)。使用GPS記錄所有巢箱的地理信息。野外調查時間從2018年5月初至2019年5月初,每隔1—2個月收集1次筑巢巢管并帶回室內飼養觀察,同時在對應巢箱中補充相同數目的空巢管;室內飼養觀察持續到2019年12月。
圖1 廣東車八嶺國家級自然保護區巢管位點分布及安裝示意圖Fig.1 Site distribution and installation of trap nests in the Chebaling National Nature Reserve,Guangdong Province黑色 的點代表巢管安裝的位點分布
1.3 研究方法
室內對黃緣蜾蠃的筑巢巢管進行編號,用解剖刀把巢管剖開約1/3,使用電子游標卡尺測量巢管的內部結構大小,包括:巢管內徑、蟲室長度、間室長度和前庭長度(相鄰隔板間的距離),并統計每根巢管中的蟲室數量和間室數量(圖2),最后將解剖后的巢管放入無菌玻璃試管內并用藥棉堵塞管口,以便后續觀測記錄。羽化成蟲及其寄生類群用5 mL凍蟲管獨立分裝,酒精浸泡后放入冰柜中冷凍保存以便后續相關研究。使用電子游標卡尺測量兩翅膀基部間的距離(胸寬)。

圖2 黃緣蜾蠃的巢管結構Fig.2 Nest structure of A. flavomarginatum
1.4 數據分析
廣義加合模型(generalized additive model,GAM)的特征是不需要預先設定自變量和因變量之間的具體函數關系,其可以通過自變量和因變量之間的光滑函數來構建數學關系,所以能最大程度符合原始數據的規律[48]。該模型的數學表達式如下[49]:
F(x)=f1(x1)+f2(x2)+…+fn(xn)+b0
式中,F(x)為鏈接函數,fn為解釋變量關系的非參數函數,b0為常數截距項。
本研究中,運用GAM模型分別對巢內徑、巢室長度、巢室數量、雌雄數量及雌蟲胸寬進行回歸分析。數據經過Excel 2016軟件統計后,利用R 3.6.2軟件中“mgcv”包中的“gam”函數對數據進行GAM模型分析[50],通過“plot”函數實現所有的圖形可視化[51],分析結果通過P值和調整判定系數(R2)來表征。其中,P值代表統計結果的顯著性水平,本文的顯著性水平取P<0.05,R2用于判定回歸方程的擬合效果,值域為[0,1],值越大擬合效果越好。
2 結果與分析
2.1 海拔梯度對巢管大小選擇的影響
對野外回收167根黃緣蜾蠃筑巢巢管的內徑和長度進行測量發現,雌蟲偏好選擇的巢管內徑范圍在5.63—19.13 mm之間,平均長度為(11.98 ± 2.20)mm(n=167)。對不同海拔梯度下該蜂的筑巢巢管內徑數據進行回歸分析發現,隨著海拔梯度增加,巢管內徑呈現先增后減的趨勢(R2=0.28,P<0.01)(圖3)。

圖3 基于GAM模型的海拔與巢管內徑最優擬合結果 Fig.3 Results of GAM analysis on altitude and nest inner diameter 實點表示觀測的巢管內徑,實線為模型預測的擬合曲線,虛線表示模型預測的95%置信區間,縱坐標為平滑函數的值
2.2 海拔梯度對巢室長度的影響
通過室內對黃緣蜾蠃筑巢巢管的相關參數進行測量發現:該蜂筑巢巢管的蟲室、間室、前庭的長度范圍分別在8.68—45.65 mm、4.63—39.26 mm、3.75—49.98 mm,平均長度分別為(24.64 ± 7.51)mm(n=302)、(16.21 ± 7.30)mm(n=152)、(19.60 ± 10.39)mm(n=122)。回歸結果顯示,蟲室、間室和前庭與海拔之間具有相關性,但解釋程度較低,為弱相關,模型顯示蟲室、間室和前庭的長度隨著海拔增加總體上都呈現先減后增的趨勢(R2=0.14,P<0.01;R2=0.16,P<0.01;R2=0.46,P<0.01)(圖4)。

圖4 基于GAM模型的海拔與巢室長度(蟲室長度,間室長度,前庭長度)最優擬合結果Fig.4 Results of GAM analysis on altitude and nest length (brood cell length;intercalary cell length;vestibular cell length)實點表示觀測的巢室長度,實線為模型預測的擬合曲線,虛線表示模型預測的95%置信區間,縱坐標為平滑函數的值
2.3 海拔梯度對巢室數量的影響
對黃緣蜾蠃所有筑巢巢管內的巢室數量統計發現:每個巢管中的蟲室數量在1—6之間,平均為(2.39 ± 1.06)(n=167);間室數量在1—4之間,平均值為(1.66 ± 0.71)(n=167)。對該蜂不同海拔梯度下的筑巢巢管的蟲室數目和間室數目進行回歸分析發現,隨著海拔梯度增加,蟲室數和間室數均表現為先增后減的趨勢,但解釋程度較低,為弱相關(R2=0.10,P<0.001;R2=0.15,P<0.01)(圖5)。

圖5 基于GAM模型的海拔與巢室數量(蟲室數量,間室數量)最優擬合結果Fig.5 Results of GAM analysis on altitude and nest number (brood cell number;intercalary cell length)實點表示觀測的巢室數量,實線為模型預測的擬合曲線,虛線表示模型預測的95%置信區間,縱坐標為平滑函數的值
2.4 海拔梯度對子代雌雄分配的影響
2018年5月初至2019年5月初期間,只有46%的羽化蜂被成功收集(在巢管解剖前就有蜂羽化出巢,或者在室內孵育過程因寄生生物、病蟲害造成死亡,以及未知原因造成的死亡),共173只,其中雌性個體55只,雄性個體118只,雌雄性比為1∶2.1。對該蜂不同海拔梯度下的雌雄數量進行回歸分析發現,隨著海拔梯度增加,單根巢管內的雄性數量呈現多波峰的非線性關系(R2=0.14,P=0.02),而單根巢管內的雌性數量呈先增后減的趨勢(R2=0.28,P<0.01),但對單根巢管內的雄性數量解釋程度較低,為弱相關(圖6)。
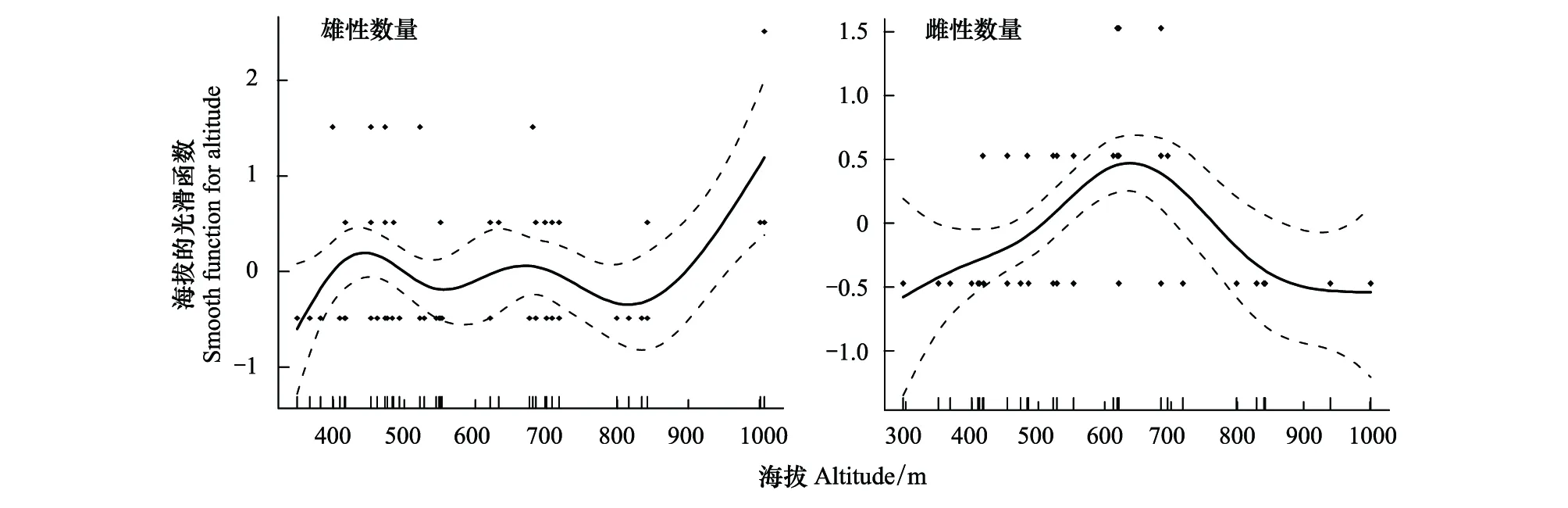
圖6 基于GAM模型的海拔與子代雌雄數量(雄性數量,雌性數量)最優擬合結果Fig.6 Results of GAM analysis on altitude and number of male and female offspring (male number;female number)實點表示觀測的子代雌雄數量,實線為模型預測的擬合曲線,虛線表示模型預測的95%置信區間,縱坐標為平滑函數的值
2.5 海拔梯度對雌性成蟲胸寬的影響
對羽化出巢后的雌性黃緣蜾蠃的胸寬進行測量發現:該蜂雌性成蟲的胸寬在4.50—6.16 mm,平均值為(5.23 ± 0.35)mm(n=55)。對該蜂不同海拔梯度下雌性成蟲的胸寬大小進行回歸分析發現:隨著海拔梯度增加,雌性成蟲胸寬總體呈先增后減的趨勢,在海拔400—700 m之間逐漸上升,在約700 m之后逐漸下降(R2=0.31,P<0.01)(圖7)。

圖7 基于GAM模型的海拔與子代雌性成蟲胸寬最優擬合結果 Fig.7 Results of GAM analysis on altitude and female breast wide of offspring實點表示觀測的子代雌性成蟲胸寬,實線為模型預測的擬合曲線,虛線表示模型預測的95%置信區間,縱坐標為平滑函數的值
3 結果和討論
獨棲蜂類依賴于現存的孔洞筑巢,一旦雌蜂筑好巢穴,其后代從卵、幼蟲到蛹會一直在巢穴中生活[37],因此雌蜂對巢穴大小的選擇就顯得非常重要。目前已有研究表明巢穴尺寸會影響到這類昆蟲后代的性別比偏向,具體而言,在內徑較小的巢穴中,雄性比例會上升,而在內徑較大的巢穴中,雌性比例會上升[52]。同樣地,郭鵬飛等人在研究黃緣蜾蠃后代性別的分配時也發現,隨著巢管直徑的增加,雌性比例增加的現象[9]。為了進一步探究這種現象背后的原因,本研究將海拔梯度作為解釋變量,實驗結果發現了,隨著海拔的增加,黃緣蜾蠃選擇的巢管內徑尺寸先增后減,而后代中雌性數量也出現了先增后減的趨勢。盡管研究還發現了后代中雄性數量有著與雌性數量變化相反的趨勢,但是模型顯示了雄性數量與海拔之間相關性較弱。因此,本研究認為海拔梯度主要影響了黃緣蜾蠃對巢管內徑的選擇以及對后代中雌性的投資比例。根據以往的研究結果顯示,巢穴附近的食物豐富度會影響獨棲蜂類對后代性別比的分配。Seidelmann等人認為,在巢管空間不限制下,親本將調整對后代的投資至上限,這種調整將有利于體型較大的雌性個體[1,53],而巢穴的內徑大小又決定了每個蟲室中可投資的食物上限[54]。因此,在食物可觀的情況下,親本會選擇較大的巢管來繁殖更多的雌性后代。吳艷光在調查長白山地區的昆蟲多樣性中發現,在不同海拔高度的垂直帶中,針闊混交林群落相比其它群落具有更高的昆蟲物種數和個體數[55]。梁召俊在調查川西高原森林昆蟲種類多樣性發現,鱗翅目昆蟲在人工林的數量總體少于原始林[56]。而對于黃緣蜾蠃,該蜂是一種以鱗翅目幼蟲為食的獨棲性捕食昆蟲,據研究顯示,車八嶺植被垂直類型為:從低海拔以常綠闊葉林、針葉林和人工林為主(330—500 m),中海拔以常綠闊葉林和針葉林為主(500—800 m),高海拔以灌叢草坡為主(800—1256 m)[57]。因此在該片區,中海拔的常綠闊葉林和針葉林具有的昆蟲多樣性更高,黃緣蜾蠃可以獲得更多的食物資源,從而促使其選擇更大內徑的巢穴,以繁殖更多的雌性后代。
獨棲蜂的親本對后代體型有高度控制權,食物的攝入量與獨棲性后代體型大小密切相關[6]。Alonso等人在調查一種泥蜂時發現,體重更大的泥蜂能捕捉獵物體重的范圍更大[52]。此外,對于這類獨棲昆蟲而言,雌性通常扮演者筑巢、捕食和繁殖的角色,而雄性通常只負責求偶交配。顯然,雌性在種群中具有更多的功能屬性。在本研究,黃緣蜾蠃后代中雌蜂的數量和胸寬都隨海拔增加先增后減,而雄性數量與海拔呈弱相關性。這可能就是因為,雌性在種群中的重要地位決定的。而雌蜂的這種重要性,促使親本在食物更多的中海拔區對雌性子代具有更多的投資,結果表現為更大體型和更多數量的雌性后代,以提高種群生存和繁殖的競爭力。相反,對于雄性的投資就顯得不太重要。
獨棲蜂類的巢管結構主要包括蟲室(brood cells/provisioned cells)、間室(intercalary cell)及前庭(vestibular cell)3個部分[30]。Polidori等人[5]認為,野生蜂類親本除了考慮選擇適宜的巢管大小外,還要考慮對筑巢空間的分配,即雌蜂需要根據現有環境權衡后代大小和數量間的關系,Smith和Fretwell研究發現,親本對子代數量上的投資大于后代個體大小的投資,因為子代數量對整個種群穩定的維持更加顯著[58]。而在海拔梯度上,本研究結果發現,盡管隨著海拔增加,蟲室長度、間室長度、蟲室數和間室數分別有對應的變化趨勢,但模型的解釋程度較低,這些因子與海拔之間呈弱相關。相反,前庭長度表現出較高相關性。在Asís等人的研究中發現,較長的前庭不僅能夠減少子代在發育最初階段的死亡率,還能夠減少巢穴外剩余空間大小,以防止其它雌性前來筑巢[59]。因此,結果表明了海拔在筑巢空間上主要體現在對前庭長度的影響。
對獨棲蜂類而言,在資源有限的自然環境下,如何最佳利用筑巢資源以提高種群對環境的適應性是非常重要的。本研究顯示,海拔主要影響的是黃緣蜾蠃對巢管內徑大小的選擇,而不是對巢室長度和數量的分配,同時,黃緣蜾蠃對巢管內徑的選擇可以影響到后代性別比,這種策略主要體現在親本對雌性子代的投資,而不是雄性子代。由此可見黃緣蜾蠃可通過調整其筑巢策略以應對不同海拔下的環境,但對于其它獨棲蜂類而言,是否也具有一致的調整策略尚未清楚,仍需進一步研究。
