黃鱔Dmrt1基因5′端側翼序列克隆和報告基因載體活性分析
2022-05-25 07:09:42杜怡芳朱信超李賢慧曲憲成
淡水漁業 2022年3期
杜怡芳,朱信超,李賢慧,曲憲成
(上海海洋大學水產與生命學院/上海海洋大學水產科學國家級實驗教學示范中心,上海/上海海洋大學上海水產養殖工程技術研究中心,上海 201306)
魚類在脊椎動物中分布最廣且種類最多,與高等脊椎動物相比,魚類性別決定機制更加復雜多樣。脊椎動物的性別決定機制包括基因性別決定機制(GSD)、環境性別決定機制(ESD)和基因-溫度性別決定機制(GSD+TE)。黃鱔()是由雌性到雄性的自然性逆轉的溫帶淡水魚,是性逆轉研究機制的模式物種。相關研究報道顯示下丘腦—腦垂體系統在誘發黃鱔性轉變過程中發揮著重要的作用,也有學者在基因表達水平上對黃鱔性逆轉進行了研究,如450、45011和1等性別相關基因表達量的變化對黃鱔的性逆轉起著重要的調節作用。黃鱔性逆轉的研究不僅對其發育機制起著非常重要的作用,也為進一步研究脊椎動物性別分化及生物進化提供重要的理論依據。
對于性別發育相關基因研究,基因家族一直以來都是重點研究對象之一。1參與許多動物的性別發育,是繼后發現的一個與性別決定相關的重要基因,1在基因調控系統中扮演著重要角色。基因家族成員與性別決定基因和-3具有共同特征,其序列能編碼一個與DNA特異結合的、且保守的蛋白功能區域,類似于鋅指結構的保守基序。目前研究表明,基因家族參與了多種動物的性別發育。對于脊椎動物的相關報道顯示,在多種脊椎動物中1基因功能的表達既有相似性又有特異性。HUANG等在黃鱔研究結果中報道了5個基因,即1,2,3,4和5,并利用黃鱔性腺,通過RACE技術克隆了黃鱔1的四個亞型,命名為1a,1b,1c和1d。通過實時熒光定量技術研究結果顯示,1的四個亞型是通過不同方式剪接而成的,在性腺各時期都有表達,這表明1基因在黃鱔性別決定和分化中起著重要作用。
1在進化過程中具有高度保守性,有關報道顯示1位于性別調控途徑的上游,作為轉錄因子在雄性性腺分化、各種組織器官形成和功能維持中發揮重要調控作用。JIA等利用亞硫酸氫鈉DNA測序技術研究了翹嘴鲌()1啟動子在精巢和卵巢中的CpG甲基化模式,發現1的性別二態性表達受DNA甲基化調控,因此表觀遺傳修飾可能在翹嘴鲌的性腺分化中起關鍵作用。JEONG等發現細棘海豬魚()GATAx和C/EBPa結合位點可以上調1基因的表達,Sox5蛋白可以下調1的表達。這些結果表明1基因在不同物種體內的調控作用機制會有所不同。
目前,對于黃鱔1基因5′端側翼序列克隆的研究未見報道。本研究對黃鱔1基因的5′端側翼序列進行克隆,對其進行測序分析,將構建的pGL3-enhancer-1報告質粒與海參熒光素酶內參質粒pRL-TK用Lipohigh轉染試劑共轉染至HEK293細胞,在轉染32 h后,檢測啟動子雙熒光素酶的表達活性,以期為后續揭示1基因對黃鱔性別分化轉錄調控機制提供理論依據。
1 材料與方法
1.1 實驗用魚
實驗黃鱔購自于上海市臨港新蘆苑農貿市場,取黃鱔,切斷頸椎將其處死,打開腹腔,每管收集約100 mg的肌肉組織,-80 ℃保存備用。由上海海洋大學實驗動物倫理委員會的批準進行相應實驗。
1.2 主要試劑
DNA提取試劑盒、增強型瓊脂糖凝膠DNA回收試劑盒、無內毒素質粒提取試劑盒等購自天根生化科技(北京)有限公司;DH5細胞購于天根生化(上海)有限公司;DNA聚合酶、I、I酶等購自Takara工程(上海皓嘉)有限公司;Lipohigh脂質體轉染試劑購于生工生物工程(上海)有限公司;DMEM和Opti-MEM培養基購于賽默飛科技(中國)有限公司;雙熒光檢測試劑盒購于翊圣生物(上海)有限公司;pGL3-enhancer報告基因載體、pRL-TK內參質粒及HEK293細胞由上海海洋大學免疫學實驗室張慶華教授惠贈;本次實驗所有的引物合成及測序在生工生物工程(上海)有限公司進行。
1.3 黃鱔Dmrt1基因5′端側翼序列的克隆
運用基因組DNA提取試劑盒對黃鱔肌肉組織進行DNA提取,并根據黃鱔1基因組序列設計引物:上游5′-3′引物GGGGTACCATTAGAATATCGTGGACAAAAATTATTTC;下游5′-3′引物GCGAGCTCAGTCCAGCACCTGCTTGCGCTGCTTGTC。以黃鱔基因組DNA為模板進行PCR擴增,反應體系為:模板DNA,1 μL;10×擴增緩沖液,5 μL;dNTP MIX,1 μL;上、下游引物,各2 μL;DNA聚合酶,1 μL;補加無菌去離子水至50 L。PCR反應程序為:94 ℃預變性5 min;經94 ℃ 15 s,退火60 ℃ 30 s,72 ℃延伸4 min,共40個循環;72 ℃徹底延伸10 min;4 ℃保存。PCR產物用1%的瓊脂糖凝膠電泳檢測,最后目標片段經增強型DNA回收試劑盒進行割膠純化。
1.4 黃鱔Dmrt1基因5′端側翼序列中轉錄因子結合位點的預測分析
利用TESS-Transcription Element Search System在線數據庫:https://www.cbil.upenn.edu/tess對黃鱔15′端側翼序列進行預測分析5′端側翼序列中的轉錄因子結合位點。
1.5 黃鱔Dmrt1基因5′端側翼序列構建到pGL3-enhancer載體
將回收純化的1 PCR產物和報告載體pGL3-enhancer同時進行雙酶切,兩個酶切位點分別是Kpn I和Sac I。對酶切產物和pGL3-enhancer載體進行割膠回收,利用T4 Ligase連接酶對回收的PCR產物和報告載體pGL3-enhancer酶切產物進行16 ℃過夜連接,將連接產物轉化至大腸桿菌DH5中37 ℃培養過夜,挑取陽性克隆進行菌液PCR鑒定后篩選陽性克隆送至上海生工生物公司測序,將測序正確的菌液經質粒提取得到pGL3-enhancer-1重組質粒,最后對其雙酶切鑒定。
1.6 雙熒光素酶報告基因檢測
HEK293細胞培養于10% FBS-DMEM培養基,設置培養箱內的參數為37 ℃,5% CO,每兩天傳代1次。在轉染前的24 h左右,將細胞接種于24孔細胞培養板,接種密度約為1×10/孔,匯合率達到80%~90%即可進行轉染,轉染過程參照Lipohigh脂質體高效轉染試劑說明書,每孔中的比例為DNA∶Lipo=1∶3,將100 ng目標質粒(pGL3-enhancer和pGL3-enhancer-1)和10 ng內參質粒pRL-TK共轉染至HEK293細胞。轉染32 h后,進行啟動子報告基因雙熒光素酶活性鑒定,計算出Firefly Luciferase與Renilla Luciferase的比值。轉染時細胞處理分為空白對照組pGL3-enhancer質粒和實驗組pGL3-enhancer-1重組質粒。每次實驗設3個復孔,每組實驗重復3次。
1.7 數據分析
采用Graphpad Prism5軟件分析數據,單因素方差檢驗數據,以平均值±標準誤(Means±SEM)表示。當<0.001時,表示差異極顯著。
2 結果與分析
2.1 Dmrt1 5′端側翼序列的克隆
以黃鱔基因組DNA為模板,根據NCBI數據庫獲得的1基因組序列設計引物,PCR產物經電泳檢測,擴增的條帶與目標條帶大小一致。1 5′端側翼序列中的大小為1 500 bp左右(圖1)。
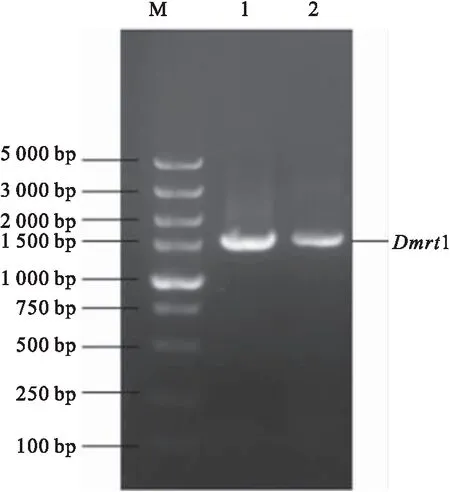
圖1 Dmrt1基因5′端側翼序列擴增片段
2.2 黃鱔Dmrt1 5′端側翼序列轉錄因子結合位點預測
利用TESS-Transcription Element Search System在線數據庫進一步對潛在的轉錄因子結合位點進行分析顯示:克隆所獲得的1 519 bp黃鱔1基因5′端側翼序列中存在多個轉錄因子反應元件,如Oct-1、GATA-1、Sp1、Pit-1a和TBP等結合位點(圖2)。其中,GATA-1、SRY、Oct-1和SP1家族蛋白和C/EBPalp的轉錄因子結合位點出現頻率較高。這些轉錄因子可能對黃鱔1基因的表達產生影響。

圖2 黃鱔Dmrt1基因5′端側翼序列及功能反應元件
2.3 黃鱔pGL3-enhancer-Dmrt1 5′端側翼序列雙酶切鑒定
pGL3-enhancer-1 5′端側翼序列經Kpn I和Sac I雙酶切后電泳,分別得到5 000 bp左右載體片段和1 500 bp左右目的片段。其大小與預期條帶一致。經測序證實,黃鱔1 5′端側翼序列正確,亦證明了構建的雙熒光素酶重組報告質粒正確(圖3)。
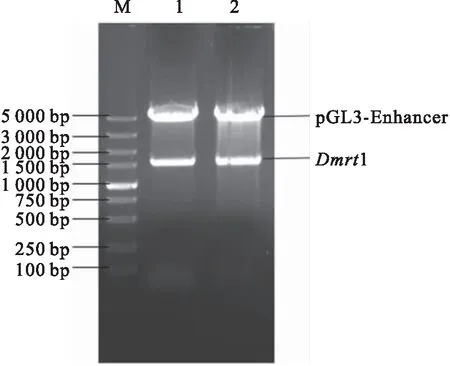
圖3 重組質粒pGL3-enhancer-Dmrt1雙酶切鑒定
2.4 黃鱔pGL3-enhancer-Dmrt1 5′端側翼序列雙熒光素酶的檢測
將pGL3-enhancer質粒和構建的pGL3-enhancer-1重組質粒分別轉染至HEK293細胞,雙熒光素酶檢測實驗結果顯示,pGL3-enhancer-1質粒轉染實驗組的相對熒光素酶活性表達極顯著高于對照組pGL3-enhancer質粒的活性表達(圖4)。表明構建的pGL3-enhancer-1報告基因載體具有強啟動活性。

圖4 重組質粒pGL3-enhancer-Dmrt1轉染HEK293細胞后熒光素酶相對表達量
3 討論
為解析黃鱔性別分化相關作用機制,眾多學者在黃鱔性別決定和性別分化的關鍵基因方面作了許多的研究。有研究顯示,將1基因導入帶有性染色體XX的雌性青鳉(),可以使遺傳雌性體表現出雄性特征,誘導雌性發生性轉化。1基因在魚類中的表達也具有組織特異性,如黑鯛()1基因只在精巢中表達,在成熟精巢中表達水平較高;與黑鯛一樣,半滑舌鰨()中1基因僅在成熟雄性的精巢中特異表達,而在其它組織中不表達;大口黑鱸()1基因在成熟魚的精巢中表達量高,而在心臟、肝臟、大腦等其他組織中表達量弱,并在不同時期的雌雄性腺中表現出性別二態性,且在雌魚中表達量很低,隨著卵巢成熟逐漸降低。對半滑舌鰨的研究結果顯示1轉錄激活因子樣效應核酸酶能有效地誘導該基因的突變,通過構建1-TALEN質粒,并顯微注射半滑舌鰨胚胎,誘導了1基因突變,導致半滑舌鰨精巢嚴重退化,出現卵巢樣結構。在尼羅羅非魚()中,1基因僅在雄性輸精管支持細胞和上皮細胞中特異性表達,而雌性個體中不表達,雄激素用于誘導XX尼羅羅非魚性逆轉時,生殖細胞周圍細胞中1基因的表達增加。其他魚類在雄性和雌性個體中也有1基因表達,如斑馬魚的1基因在精巢和卵巢中表達,經過組織切片和原位雜交發現1基因在精巢和卵巢的生殖細胞中表達,表明1基因不僅參與斑馬魚的精巢發育,也可能參與斑馬魚的卵巢分化;大西洋鱈()1基因在精巢的生殖細胞中表達,其表達水平在精子發生過程中最高;虹鱒()1基因的表達水平在已分化的精巢中較高,但在卵巢中也有表達。因此,對于雌雄異體的魚類中,1基因的表達與雄性性腺發育和性別分化密切相關。雌雄同體魚類1基因表達水平的變化與精巢的發育有關,相關研究顯示,黃鱔的1基因在精巢發育與精子形成過程中起著重要作用。
隨著生物技術的發展,基因功能不僅需要通過實驗進行驗證,還需要通過生物信息數據進行整合分析,通過各種生物信息學軟件進行預測。不同的預測軟件基于不同算法的基礎,每個軟件由于條件有限,都有一定的局限性,因此,需要使用各種合適的分析和預測軟件,整合各軟件獲得的數據,提高數據預測的正確性,為進一步的實驗奠定堅實的基礎。基因的5′端側翼序列中存在作為基因表達調控中重要的順式作用元件,在基因表達調控網絡中有大量信息,需要對基因5′端側翼序列的結構有一定的認識,基因5′端側翼序列中的轉錄因子結合位點會影響基因的轉錄活性,因此,對基因5′端側翼序列的分析、結合位點的識別及不同物種基因5′端側翼序列差異性的研究對于基因調控具有重要的現實意義。而pGL3-enhancer是一種不含啟動子的表達載體,具有氨芐抗性和雙熒光素酶(螢火蟲熒光及海腎熒光),且具有靈敏度高,易檢測等特點,廣泛應用于硬骨魚類基因5′端側翼序列活性分析的研究。當啟動子片段插入載體后,經轉染細胞系后進行表達檢測,當啟動子片段表達轉錄活性時,它將驅使雙熒光素酶報告基因的表達,從而產生不同于正常值的熒光。眾多學者利用該方法檢測基因啟動子的活性。
HE等克隆了四川裂腹魚()1 1 215 bp啟動子,并對其核心調控區域進行了分析,測序結果顯示四川裂腹魚1基因啟動子中存在正調控和負調控區域,同時表明其甲基化水平存在性別差異,另外1的表達與其啟動子CpG甲基化顯著負相關。本試驗通過TESS-Transcription Element Search System在線數據庫對潛在轉錄因子結合位點進行預測顯示,黃鱔1基因啟動子序列中存在的結合位點,如:AP-1、Oct-1和C/EBPa等真核生物通用的轉錄因子結合位點以及與性別相關基因的蛋白結合位點,如:SRY、Sox3和Sox5等。BOYER等的研究表明,不同物種的1基因5′端側翼序列具有很大的同源性,通過比較人、豬和小鼠的基因5′端側翼序列,表明它們在轉錄起始位點上游1 000 bp至3 000 bp左右具有多個保守性超過60%的調控區。其中,近端調控區主要包含Sp1、Sp3和Egr1等一些通用的轉錄因子的潛在結合位點,遠端調控區包含幾個Gata因子結合位點。在許氏平鲉()1基因5′端側翼序列中有許多作用元件與3和9基因相同,如Oct-1、GATA-3、FOXD3和AP-1等轉錄激活蛋白結合位點,以及與性別相關基因的蛋白結合位點,如:Sox5、Sox9和SRY等,而SRY和Sox5結合位點與1基因的調控機制在魚類中已有報道。此外,在許氏平鲉1基因5′端側翼序列中,預測了C/EBPa、STATx和GATAx等結合位點。GAO等在斑馬魚1基因近端5′端側翼序列中存在Sox5蛋白的結合位點,且證明Sox5蛋白能抑制1的表達。
本試驗成功克隆了黃鱔1基因5′端側翼1 519 bp序列,構建了1基因5′端側翼序列報告基因表達載體,利用雙熒光素酶報告基因檢測實驗證明了該基因5′端側翼序列具有極顯著的啟動活性。并通過對黃鱔1基因5′端側翼序列分析、結合位點的識別,為進一步研究其相關轉錄因子和調控功能奠定理論基礎。而許多功能是通過轉錄因子之間相互協調發揮作用的,在今后的試驗研究中,仍需通過構建的重組質粒pGL3-enhancer-1報告載體進行定點突變、缺失,以及染色質免疫沉淀等方法,對預測的轉錄因子結合位點的作用機制進行深入研究。
