筇竹構件生物量積累分配特征及其與土壤物理性質的關系
2022-05-26 08:32:14吳義遠董文淵謝澤軒
生態學報 2022年9期
關鍵詞:分配
吳義遠,董文淵,浦 嬋,謝澤軒,鐘 歡,李 吉,張 煒
1 南京林業大學林學院,南京 210037 2 西南林業大學筇竹研究院,昆明 650224 3 大關縣林業和草原局,昭通 657400 4 西南林業大學林學院,昆明 650224
生物量是體現植物種群在生態系統中生長狀況的重要指標[1—2],生活在異質性生境的植物會產生不同程度的生物量積累和分配特征,具體表現為構件生物量分配的“可塑性”,其決定了植物對異質性資源的獲取能力[3—5]。即植物在生長發育的過程中會權衡分配資源給稈、枝、葉等構件,通過不斷優化其生物量分配格局來適應環境[6—7]。作為重要的環境因素之一,土壤物理性質的改變勢必會造成植物生物量積累和分配的差異[8—9]。然而,這些研究多集中于土壤水分與植物生物量積累、分配的關系的研究[10—11]。例如,研究表明當土壤水分受限時,植物生物量積累會減小,在構件水平上表現為植物會增加吸收器官根的生物量分配,減少葉生物量分配,而莖等支持器官的生物量則無明顯變化[12—13]。而其他土壤物理性質如容重、毛管孔隙度、土壤持水量等與植物生物量積累、分配的關系的研究較少,且多集中于植物地上、地下生物量分配格局的或某個構件生物量積累的研究,而并沒有具體到每個構件的生物量分配的研究[14—15]。因此,需要全面研究土壤物理性質與植物生物量積累、分配的聯系。
克隆植物因存在雙構性而具有更高水平的生物量分配的“可塑性”[16]。而竹子作為典型的克隆植物,其生物量分配格局不僅是遺傳控制的結果,也是對環境長期作用的響應,反映了竹子對環境的進化的調節適應能力[17—18]。土壤作為竹子生長的基質,其理化性質與竹子的生長發育密切相關[19]。目前,竹類植物生物量積累、分配與土壤物理性質關系的研究主要集中于土壤水分對竹子生物量積累、分配的研究[20—21],如Liu[22]等研究發現,在土壤水分不利的條件下,斑苦竹(Pleioblastusmaculatus)會增大地下生物量的分配,產生更多的子代分株,從而克服不利條件。針對土壤容重、孔隙度等其他物理性質對竹類植物分配格局的影響的研究較少且不夠全面,只是集中在某些土壤物理性質對竹子特定構件的生物量積累的影響[23—25],然而土壤具有復雜的不均一性和空間變異性,且不同的竹種及構件生物量的分配格局與土壤物理性質的關系有所差異[26—28]。因此,關于竹子生物量分配策略與土壤物理性質的關系,仍需進一步的探究。
筇竹(Qiongzhueatumidinoda)為禾本科(Gramineae)筇竹屬(Qiongzhuea)植物,是國家三級保護珍稀竹種之一,為地下莖復軸混生小型竹種[29]。筇竹竹稈稈環極度隆起,是制作手杖、竹工藝品、圓竹家具和庭園綠化的佳品[30],筇竹竹筍味甘鮮嫩,營養豐富,筍干、鹽漬筍等長期暢銷日本及港澳臺地區,是云南省傳統的出口創匯大宗商品[31]。近年來,隨著筇竹筍、材市場價格的大幅度上升,天然筇竹林的無序掠奪式的經營模式,使得筇竹種群加速退化[32]。因此,開展筇竹生物學生態學研究具有重要意義。目前,天然筇竹林生物量分配及其與土壤物理性質的關系的研究尚未見報道,已有研究多集中于筇竹種群生物量特征[33],筇竹分株生物量[34]、形態[35]、竹稈解剖特征[36]對土壤水分和養分的響應,而關于筇竹林的土壤容重、孔隙度等其他土壤物理性質的研究還鮮有報道。有鑒于此,以云南省大關縣3種土層厚度的天然筇竹林為研究對象,試圖解決以下幾個問題:(1)土壤物理性質變化規律;(2)筇竹生物理積累分配特征及其與土壤物理性質關系;(3)影響筇竹生物量積累的關鍵因子。通過這三個問題的闡釋旨在為筇竹林適宜立地條件選擇、適地適竹及定向培育提供的理論指導。
1 材料與方法
1.1 研究區概況
試驗地位于云南省大關縣木桿鎮銀吉村徐家灣(28°06′40″—28°06′66″N,103°58′02″—103°59′55″E),海拔 1310—1340 m,氣候條件為中溫帶大陸性氣候,年平均溫度 10.5 ℃,最高度溫 29 ℃,最低溫度 -10 ℃,年平均降水量1200 mm,年平均蒸發量1076 mm,相對濕度 85%。土壤呈酸性,pH為4.3(Mean±SD;0—30 cm),以黃壤土為主,成土母巖以頁巖為主,土壤深度 15—120 cm。試驗地筇竹林總面積4.5 hm2,筇竹長勢較差,經營極為粗放,不實施人工土壤養分補充。筇竹林以筍用林為主,掠奪式采筍,僅保留退筍和弱小筍。其上層喬木有扁刺錐(Castanopsisplatyacantha)、鈍齒木荷(Schimacrenata)、白檀(Symplocospaniculata)等,林分郁閉度 0.3—0.5,林下植物有粗齒冷水花(Pileasinofasciata)、卷柏 (Selaginellatamariscina)、樓梯草(Elatostemainvolucratum)等,蓋度約 60% 左右。
1.2 研究方法
1.2.1樣地設置
2019年7月,在對徐家灣天然筇竹林全面踏查的基礎上,分別選取了3種類型的經營水平基本一致的樣地(表1),在每個樣地設置3個10 m×10 m的樣方,每個樣方的竹林結構調查見表2。

表1 立地條件Table 1 Site conditions

表2 竹林結構(平均值±標準誤)Table 2 Sructure of bamboo forest (Mean±SE)
1.2.2土壤樣品采集及分析
采用五點取樣法分別在每個樣地內設置5個1 m×1 m 的采樣點,清除凋落物并挖掘剖面,采用剖面原狀土取樣法取原狀土壤,即分別用環刀(100 cm3)采集0—10 cm、10 —20 cm 及 20—30 cm 土層的土壤。每層采集2個環刀樣品,采集土樣時環刀筒內應充滿樣品,采集完后環刀兩端立即加蓋,以免水分蒸發。每個環刀樣品重復3次,一共從3種土層類型種取得270個環刀樣品,對每個樣品進行編號,原狀帶回實驗室,采用環刀法測定土壤物理性質[37],每個土壤樣品至少重復測定3次,取平均值。
1.2.3植物樣品采集及分析
分別在每塊樣方內確定不同年齡(1a、2a、3a、4a)的標準竹各3株,一共108株。然后以其為中心,在來鞭和去鞭中間位置將立竹整株挖起,再分離各構件,分為稈、枝、葉、蔸、鞭和根6個構件分別測定其鮮重(其中鞭取樣以待測立竹為中心,從左右兩側竹鞭的中間截取,即截取來鞭和去鞭各一半;根包括蔸根和鞭根兩部分)。同時分別取各構件的混合樣(其中竹稈取上、中和下部)帶回實驗室,105℃殺青后,80℃下烘干至恒重,稱干重,計算各構件的含水率和生物量(g)。
1.2.4數據分析
采用Excel 2016對數據進行初步整理;采用SPSS 17.0的單因素(one-way ANOVA)方差分析不同土層厚度的筇竹生物量及土壤物理性質的差異(顯著性水平設定為α=0.05);使用 Canoco 4.5 軟件進行筇竹構件生物量與土壤物理性質的相關性分析,分別對各指標進行趨勢對應分析,得出各梯度長度最大值均小于3,故本研究均采用線性模型中的冗余分析進行趨勢分析,同時采用Monte carlo檢驗對土壤物理因子進行重要性排序,并通過Monte carlo置換檢驗確定影響筇竹構件生物量的主要因子[38]。
其中,筇竹各構件生物量占總生物量比值的各指標的計算公式如下:
葉比重(LMF)=葉生物量/總生物量×100%
枝比重(SeMF)=枝生物量/總生物量×100%
稈比重(BMF)=稈生物量/總生物量×100%
蔸比重(SuMF)=蔸生物量/總生物量×100%
鞭比重(RhMF)=鞭生物量/總生物量×100%
根比重(RoMF)=根生物量/總生物量×100%
根冠比(RSR)=(蔸生物量+鞭生物量+根生物量)/(葉生物量+枝生物量+稈生物量)×100%
2 結果與分析
2.1 不同土層厚度土壤理化性質變化
由表3分析可知,隨土層厚度的增加,筇竹林地土壤含水量(30.61%;36.70%;39.79%)、毛管持水量(22.07%;27.07%;29.43%)、最大持水量(39.65%;43.34%;44.92%)、毛管孔隙度(50.64%;57.50%;60.75%)、總孔隙度(59.02%;63.03%;66.17%)呈增加的變化趨勢,土壤容重(1.38 g/cm3;1.06 g/cm3;0.94 g/cm3)、非毛管孔隙度(8.38%;5.53%;5.42%)則呈降低的趨勢,且大部分指標差異顯著(P<0.05)。

表3 不同類型樣地0—30cm土層的土壤理化性質(平均值±標準誤)Table 3 Soil physicochemical properties in soil layer 0—30cm in different types of soil(Mean±SE)
隨土層深度增加,同一土層類型中土壤理化性質的垂直變化規律表現為隨著土層厚度的增加,土壤容重呈現出逐漸增加的趨勢,含水量、毛管持水量、最大持水量、毛管孔隙度、非毛管孔隙度、總孔隙度呈現出逐漸減少的趨勢,且大部分因子存在顯著差異(P<0.05)。
不同土層類型相同土層土壤理化性質呈現出不同的規律,其中A1和A3的土壤因子均存在顯著差異(P<0.05),具體表現為除土壤容重和非毛管孔隙度呈現出隨土層增加而減少的趨勢外,其他理化因子均呈現出隨土層增加而增加的趨勢。
2.2 不同土層厚度筇竹構件生物量特征
不同土層厚度的筇竹構件稈、枝、葉、蔸、鞭、根和總生物量皆呈現出隨土層厚度增加而增加的趨勢(表4),其中,薄土層和厚土層的稈、枝、葉、蔸、鞭、根和總生物量差異均達到顯著水平(P<0.05)。薄土層和中土層的稈、枝、葉、蔸、根和總生物量的差異顯著(P<0.05),但兩者間的鞭生物量差異則不顯著(P>0.05)。

表4 不同土層厚度筇竹構件生物量特征(平均值±標準誤)Table 4 The characteristics of module biomass of Qiongzhuea tumidinoda on different soil thickness(Mean±SE)
2.3 不同土層厚度筇竹構件生物量分配特征
隨著土層厚度的增加,筇竹構件枝、葉、蔸、鞭及根生物量和根冠比分配均隨之增加,而稈生物量分配則隨之遞減,但各構件生物量分配隨土層厚度變化的差異均不顯著(P>0.05,表5)。

表5 不同土層厚度筇竹構件生物量分配特征(平均值±標準誤)Table 5 The characteristics of biomass allocation of Qiongzhuea tumidinoda component on different soil thickness(Mean±SE)
2.4 筇竹構件生物量分配與土壤物理因子的相關關系
DCA排序結果顯示,排序軸中梯度長度最大值為0.26,因此采用冗余分析探討筇竹構件生物量分配和土層厚度之間的關系。RDA分析表明(表6),筇竹構件生物量分配在第Ⅰ軸、第Ⅱ軸的解釋量分別為86.3%和1.1%,即前兩軸7種土壤因子能累計解釋其生物量分配的87.4%,且其生物量分配與土壤因子關系的累計解釋量達到了100%。表明前兩軸能夠反映筇竹構件生物量分配與土壤因子關系的絕大部分信息,且第Ⅰ軸起決定。

表6 筇竹構件生物量分配RDA排序的特征值及累積解釋量Table 6 Eigenvalues and cumulative variances of RDA ordination of biomass allocation of Qiongzhuea tumidinoda component
從筇竹構件生物量分配與土壤物理因子的二維排序圖可知(圖1),除非毛管孔隙度外,其他土壤物理因子的箭頭連線均較長,表明能夠較好的解釋生物量分配的差異,這與表7的分析相一致。其中:土壤毛管持水量、總孔隙度、非毛管孔隙度與稈、鞭生物量;土壤含水量、最大持水量、毛管孔隙度、總孔隙與葉、枝、蔸生物量;土壤含水量、最大持水量與根生物量的夾角小且方向基本一致,說明總孔隙度、毛管持水量對稈、鞭生物量存在顯著正效應;土壤含水量、毛管孔隙度主要影響枝、葉、蔸生物量的大小;土壤含水量、最大持水量是影響根生物量大小的主導因子;而容重則與所有構件生物量表現出顯著的負相關性。這些結果表明土壤物理因子對筇竹構件生物量分配有著不同程度的促進或抑制作用,且其促進作用占主導地位。

圖1 筇竹構件生物量分配與土壤因子的冗余度分析Fig.1 Redundancy analysis of biomass allocation of Qiongzhuea tumidinoda componentSeb:稈生物量Stem biomass;Bb:枝生物量 Branch biomass;Lb:葉生物量Leaf biomass;Sub:蔸生物量Stump biomass;Rhb:鞭生物量Rhizome biomass;Rob:根生物量Root biomass;SMC:含水量Soil moisture;BD:容重Bulk density;CWH:毛管持水量Capillary water holding capacity;MWH:最大持水量Maximum water holding capacity;CP:毛管孔隙度Capillary porosity;NCP:非毛管孔隙度Non-Capillary porosity;TPO:總孔隙度Total porosity
綜上可以看出,不同土壤物理因子對筇竹構件生物量分配的影響差異較大,進一步對環境因子進行Monte carlo檢驗排序,研究土壤物理因子對生物量分配的重要性大小。各土壤因子對其生物量分配的貢獻如表7所示。指標的重要性排序依次為土壤含水量>總孔隙度>最大持水量>容重>毛管孔隙度>毛管持水量>非毛管孔隙度。土壤含水量、總孔隙度、最大持水量、容重、毛管孔隙度及毛管持水量的解釋量皆達到了顯著水平(P<0.05),分別占總信息量的79.0%、76.1%、74.3%、62.1%、61.2%和18.7%。
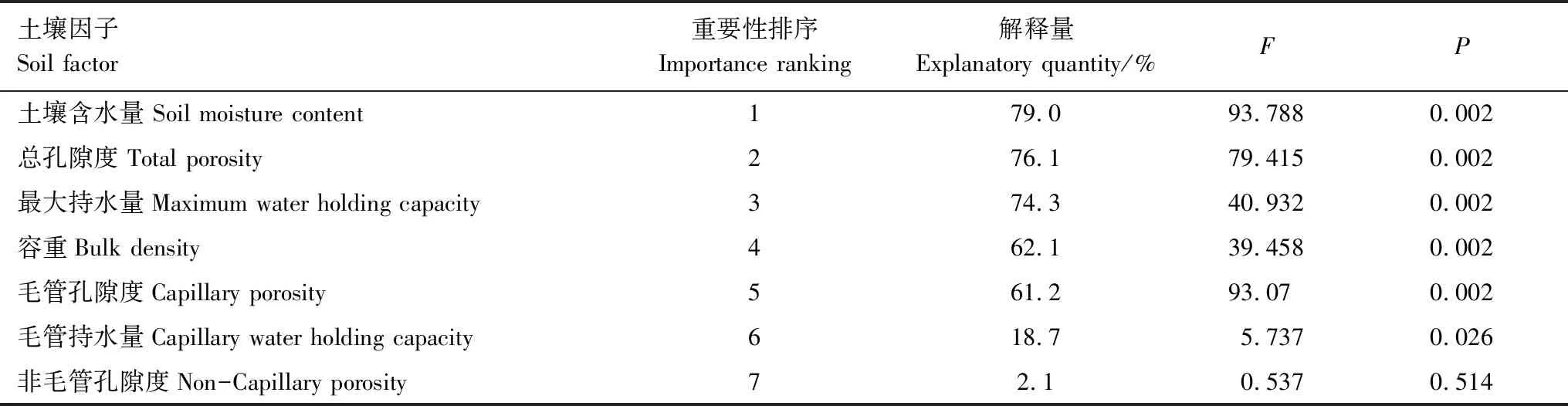
表7 土壤物理因子變量解釋的重要性排序和顯著性檢驗結果Table 7 Importance and signification level of soil physical factors
3 討論
土層厚度對筇竹總生物量和構件生物量大小有著不同程度的影響,具體表現為隨著土層厚度的遞增,筇竹總生物量及構件生物量隨之顯著遞增。這是由于厚土層的物理性質最好,而土壤物理性質直接決定了土壤的養分和水分供給能力[39],使得生長其中的筇竹及構件積累了較大的生物量。而在水分含量較低的薄土層中,筇竹為了平衡種群資源的分配,其個體及構件生物量較低。這與大部分學者的研究結果一致,例如:苦斑竹在土壤水分含量較高的條件下所積累的地上、地下及總生物量皆較大[22]。石灰巖地淡竹(Phyllostachysglauca)分株生物量及構件生物量在土壤物理性質較好的低裸巖率地處最高[26];雷竹(Phyllostachysviolascens)分株生物量和地上、地下生物量隨著土壤水分的增加皆顯著增加[40]。
McConnaughay等[41]提出的生物量最優理論認為,植物通過調節各構件生物量分配來響應環境因子的變化,目的是最大化地獲取光、水分及養分資源。因此,為了增強生態適應性,植物會減少非限制性資源構件的分配,而增大獲取限制性資源的構件分配。本研究發現,不同土層厚度的筇竹構件生物量分配存在差異,但這種差異性均不顯著,可能原因一是試驗地氣候條件決定了土壤水分等物理因子對筇竹生長限制性不強,更多的可能是土壤養分、生長空間等的影響。即本研究中土壤物理因子還沒有達到影響筇竹產生生物量可塑性對策的程度,這符合可塑性成本與限制假說,即無論可塑性是適應的或非適應的,都可能涉及其產生和維持的成本,只有收益大于成本時,可塑性才會被檢測到[42—43]。二是忽略了不同土層厚度的筇竹立竹的平均密度、胸徑的差異所造成的影響,研究表明,植物種群胸徑、密度的大小會導致個體之間產生地上部光的競爭及地下部水分的競爭[44],如四季竹(Oligostachyumlubricum)在種植密度超出合適范圍時,會將更多地生物量分配給構件稈,從而促進立竹縱向生長來獲取更多的光資源[7]。
但隨著土層厚度的遞增,筇竹各構件生物量存在著一定的變化規律。研究表明,葉片是植物蒸騰作用的器官,其生物量分配與水分利用密切相關[45]。在本研究中,葉的生物量分配存在隨土層的遞增而遞增的趨勢,這可能是在水分資源條件較低的薄層土生境中,筇竹通過減少葉片生物量分配,從而降低蒸騰速率來減少水分的損失,以保證其存活。筇竹稈生物量分配隨著土層厚度的遞增而遞減,這與竹子莖稈具有一定的貯水功能的研究結果相一致[46]。即在水分相對貧瘠的薄土層和中土層生境中,筇竹增大對儲水構件稈的生物量分配,以儲存更多的水分來維持其正常生長。梁寬等[47]對淡竹(Phyllostachysglauca)的研究也得到了相似的結論,即在土壤水分等土壤物理因子較差的高裸巖率生境中,淡竹會通過增加稈生物量、降低葉生物量來克服不利條件。3種土層生境的筇竹的根冠比隨著土層厚度的遞增而遞增,這與雷竹、斑苦竹在低水分資源下增大地下部分生物量分配有所差異[22,40],其原因進一步說明試驗地筇竹生長并不是水分的主要影響,可能是地下生長空間或養分等。總之,不同處理間筇竹生物量分配雖然差異不顯著,但亦呈現了一定的規律性。在生產上,可以選擇土層厚度較大的立地來造林,或者通過增加土層厚度來改造筇竹低效林,從而達到促進筇竹的生長發育、種群穩定與擴張的效果。
冗余分析表明,筇竹構件生物量分配與土壤含水量、最大持水量、毛管持水量、總孔隙度、毛管孔隙度皆呈顯著正相關,與毛管孔隙度呈正相關,這與前人的研究結果基本一致[48—53],即土壤含水量可以直接影響土壤的酶促作用及生物化學的強度,改善土壤肥力。總孔隙度直接決定了土壤養分的生物有效性,并對土壤肥力的發揮和通氣透水性有直接影響。管孔隙度越大,則土壤保持水能力越好。而非毛管孔隙度越大,透氣性則越好。最大持水量和毛管持水量分別反映了土壤的蓄水能力和供水能力。綜合可知,這些物理因子對植物的生長皆有不同程度的促進作用。筇竹構件生物量分配與土壤容重呈現顯著的負相關,這和李倩倩等[54]研究結果一致,土壤容重的變化直接導致土壤緊實度的改變,土壤容重越大,土壤緊實度越大,土壤儲水空間越小,不利于植物生物量的積累,因此兩者呈現負相關性。
本研究得出土壤物理因子與筇竹構件生物量有著不同程度的關系,土壤含水量、總孔隙度、最大持水量和容重起著決定性的作用,這與其他的學者研究結果有所差異,其原因是研究地土壤特性和植物生物學特性的差異所造成的結果。如張志云等[55]研究得出在花崗巖為母巖發育的土壤中,影響杉木(Cunninghamialanceolata)生物量積累的主導物理因子是容重和非毛管孔隙度。而劉姝媛等[56]研究表明土壤毛管孔隙度和毛管持水量是影響廣東省桉樹(Eucalyptusrobusta)人工林生物量積累分配的主導因子。
4 結論
(1)3種土層厚度中,厚土層的物理性質最好,中土層次之,薄土層最差,其中厚土層和薄土層的各物理因子均達顯著水平(P<0.05)。隨著土層厚度的增加,筇竹總生物量和構件生物量均顯著增加(P<0.05)。
(2)不同土層厚度的筇竹生物量分配雖然差異不顯著(P>0.05),但亦呈現了一定的規律性,即在水分貧瘠的生境中,筇竹會優先將生物量分配給稈,并通過減少葉生物量來響應水分缺失的不利環境。
(3)土壤物理性質可以解釋筇竹構件生物量分配的絕大部分變異,其中,土壤含水量、總孔隙度、最大持水量和容重是影響筇竹構件生物量分配的主導因子。在生產中,可以通過竹闊葉混交、墾覆松土等措施來改善土壤物理性質,從而促進筇竹的生長。
(4)本實驗存在一定局限性,即沒有排除筇竹立竹胸徑、密度以及土壤養分對筇竹構件生物量積累、分配所造成的影響。
猜你喜歡
天水行政學院學報(2022年4期)2022-11-18 09:02:36
艦船科學技術(2022年13期)2022-08-11 09:30:02
鐵道通信信號(2020年9期)2020-02-06 09:15:22
漢語世界(The World of Chinese)(2019年3期)2019-07-01 02:37:48
數學大王·趣味邏輯(2019年5期)2019-06-13 20:27:43
小學科學(學生版)(2019年5期)2019-05-21 01:00:18
中學生數理化·中考版(2018年10期)2018-12-07 00:44:52
經濟技術協作信息(2018年30期)2018-11-22 06:20:24
中央社會主義學院學報(2017年1期)2017-04-16 05:34:07
中國衛生(2014年12期)2014-11-12 13:12:40
