酸橙內生菌Bacillus thuringiensisBt028產幾丁質酶的發(fā)酵條件優(yōu)化
2022-05-29 02:12:28徐勤茜張澤宇陶雪婷趙梓伶李子院郝再彬李海云
食品工業(yè)科技 2022年11期
關鍵詞:影響
徐勤茜,張澤宇,陶雪婷,趙梓伶,李子院,2,郝再彬,2,李海云,2,
(1.桂林理工大學化學與生物工程學院,廣西桂林 541004;2.廣西高校食品安全與檢測重點實驗室,廣西桂林 541004)
幾丁質是由N-乙酰-D-葡糖糖胺以β-1,4 糖苷鍵連接而成的天然多糖和可再生多聚物,廣泛存在于自然界,如無脊椎動物的外骨骼、蝦蟹的外殼、真菌的細胞壁、線蟲卵殼等[1]。幾丁質是一類重要的生物資源,但由于其結構穩(wěn)定、高結晶度而難溶于水及大多數(shù)有機溶劑[2?3],其應用受到很大限制而造成巨大的資源浪費。幾丁質酶是一類能特異性催化幾丁質水解的酶[4?6],其水解產物如幾丁質寡糖、N-乙酰氨基葡萄糖等產物[7]具有良好的溶解性、生物相容性以及多種生物活性,被廣泛應用于醫(yī)藥工業(yè)[8?10]、食品工業(yè)[11]、農業(yè)[12?13]等領域。微生物是幾丁質酶的主要來源,國內外對微生物產生的幾丁質酶進行了大量研究[14?17]。盡管如此,尋找產酶性能優(yōu)良的微生物菌株仍是當前研究的主要方向之一[18]。
植物內生菌是一類生活在健康植物組織內部的,不引起宿主植物出現(xiàn)明顯病害癥狀的一類微生物[19]。在與宿主植物長期共存的過程中,植物內生菌被賦予多種獨特的生物活性,如能夠產生與宿主植物相同或相似的活性物質[20?22]、廣泛參與宿主次生代謝產物的合成與轉化[23?24]等。近年來,植物內生菌的產酶特性及其在生物催化與生物轉化領域的應用也開始受到人們的重視[25]。例如利用內生菌產葡萄糖苷酶對虎杖苷[26?27]、人參皂苷[28]的生物轉化;Harwoko 等[29]從柳葉分離出的真菌能夠對植物衍生的類黃酮進行生物轉化反應;陳智慧等[30]從花魔芋實生種子中篩選出一株具有良好的酶學性質的產β-甘露聚糖酶內生菌。
酸橙(Citrus aurantiumL.)又稱苦橙,其濃縮果汁含有豐富的黃酮類物質,具有抗氧化、抗炎癥等多種生物活性[31]。目前尚未見有酸橙內生菌的研究報道。為進一步尋找產幾丁質酶性能優(yōu)良、實用潛力大的微生物菌株,課題組前期對產幾丁質酶的酸橙內生菌進行了分離篩選,獲得一株產酶性能較好的Bacillus thuringiensisBt028 菌株。本文對該菌株產幾丁質酶的發(fā)酵條件進行優(yōu)化,為后期幾丁質酶的分離純化、酶學性質、實際應用等研究奠定基礎。
1 材料與方法
1.1 材料與儀器
酸橙 采自桂林永福蘇橋鎮(zhèn),新鮮酸橙果實;酸橙內生菌Bacillus thuringiensisBt028 分離自酸橙果實,保存于桂林理工大學生物工程實驗室;幾丁質、3,5-二硝基水楊酸、β-D-N-乙酰氨基葡萄糖 分析純,購于上海易恩化學技術有限公司;硫酸鎂、硫酸亞鐵 分析純,購于天津市大茂化學試劑廠;蛋白胨、酵母粉、瓊脂磷酸氫二鈉、磷酸二氫鈉、氯化鈉、氫氧化鈉、酒石酸鉀鈉、苯酚、結晶亞硫酸鈉、磷酸分析純,購于西隴化工股份有限公司;Bt 平板活化培養(yǎng)基:蛋白胨10 g/L、酵母粉5 g/L、氯化鈉5 g/L、瓊脂15 g/L;幾丁質酶發(fā)酵培養(yǎng)基:膠體幾丁質10 g/L、酵母粉10 g/L、K2HPO40.7 g/L、KH2PO40.3 g/L、MgSO4·7H2O 0.5 g/L、FeSO4·7 H2O 0.01 g/L,pH 7.0。
LRH-150-Z 振蕩培養(yǎng)箱 珠江留關市泰宏醫(yī)療器械有限公司;BX30R 立式壓力蒸汽滅菌器 上海博訊實業(yè)有限公司;CJ-ISFS 醫(yī)療設備超工作臺 天津市泰斯特儀器有限公司;VIS-7220N 可見分光光度計 北京瑞利分析儀器有限公司;HH-S2 型數(shù)顯恒溫水浴鍋 金壇市醫(yī)療儀器廠;ZXRD-B5110 鼓風干燥箱 上海智城分析儀器制造有限公司。
1.2 實驗方法
1.2.1 膠體幾丁質的制備 將幾丁質粉碎過100 目篩即得細粉幾丁質,參考文獻方法[32],稱取25 g 細粉幾丁質,加250 mL 85%磷酸,30 ℃反應1 d,然后加蒸餾水使膠體沉淀后,反復水洗直至中性。用0.02 mol/L NaH2PO4-Na2HPO4緩沖液(pH=6.0)調整終溶度至10%,4 ℃保存。
1.2.2 培養(yǎng)條件對Bt028 菌株產幾丁質酶的影響斜面保存的Bacillus thuringiensisBt028 經平板培養(yǎng)活化24 h 后,挑取一環(huán)接入100 mL 幾丁質酶發(fā)酵培養(yǎng)基中(250 mL 三角瓶),在溫度為30 ℃,轉速為180 r/min 的搖床上振蕩培養(yǎng)24 h,即為種子液,此時為菌株生長時間。將種子液按照3%的接種量接入到裝有100 mL pH7.0 的幾丁質酶發(fā)酵培養(yǎng)基的250 mL 三角瓶中,在溫度為30 ℃、轉速為180 r/min的搖床上振蕩培養(yǎng)48 h,此時為菌株產酶時間。發(fā)酵液在4 ℃、12000 r/min 條件下離心10 min,收集上清液即為粗酶液,4 ℃冰箱儲存,用于幾丁質酶酶活的測定。
1.2.2.1 單因素實驗 按上述搖瓶培養(yǎng)方法,分別考察碳源種類(粉末幾丁質、膠體幾丁質、淀粉、乳糖、蔗糖、麥芽糖、葡萄糖)、碳源濃度(0.25%、0.5%、1%、1.5%、2%、2.5%)、氮源種類(酵母浸粉、蛋白胨、酵母粉、硫酸銨、氯化銨、硝酸銨、硝酸鈉)、氮源濃度(0.5%、1.0%、1.5%、2%、2.5%、3%)、初始pH(5.5、6.0、6.5、7.0、7.5、8.0)、培養(yǎng)溫度(20、25、30、35、40、45 ℃)、接種量(0.5%、1%、2%、3%、4%)、裝液量(50、75、100、125、150 mL/250 mL)、搖床轉速(100、120、140、160、180、200 r/min)、生長時間(12、24、36、48、60、72、84、96、108、120 h)、產酶時間(12、24、36、48、60、72、84、96 h)等因素對菌株Bt028 產幾丁質酶的影響,以碳源種類為第一因素實驗時,其他條件為碳源濃度1%、氮源種類蛋白胨、氮源濃度1.0%、初始pH7.0、培養(yǎng)溫度30 ℃、接種量3%、裝液量100 mL/250 mL、搖床轉速180 r/min)、生長時間48 h、后續(xù)因素的實驗均在前一因素實驗的較優(yōu)結果上進行。
1.2.2.2 響應面優(yōu)化試驗 根據(jù)單因素實驗結果,選擇3 個對酶活影響較顯著的因素進行響應面設計,根據(jù)Box-Behnken 設計原理,運用Design-Expert 進行三因素三水平響應面試驗設計,共計17 個實驗點的響應面分析試驗,實驗選取的因素和水平見表1。

表1 試驗因素和水平編碼值Table 1 Test factors and level coding values
1.2.3 幾丁質酶酶活的測定 參考戴德慧等[33]方法并加以改進,分別吸取5 mmol/Lβ-D-N-乙酰氨基葡萄糖標準液0、0.2、0.4、0.6、0.8 mL 于1、2、3、4、5 號試管中,用蒸餾水補至1 mL,再加入1 mL DNS試劑,混勻后沸水浴中煮沸5 min,冷卻后用蒸餾水稀釋至10 mL,以1 號試管為對照,測定540 nm 波長處吸光度,以A540為縱坐標、β-D-N-乙酰氨基葡萄糖濃度C(mmol/L)為橫坐標繪制β-D-N-乙酰氨基葡萄糖標準曲線,其標準曲線方程為A540=0.0074C-0.03(R2=0.9987)。
粗酶液酶活測定[33]:取0.5 mL 粗酶液與0.5 mL含1.0%膠體幾丁質的磷酸緩沖液(0.1 mol/L,pH7.0)混合,50 ℃保溫20 min 后,沸水浴10 min,冷卻后加入1 mL DNS 試劑,在沸水中煮沸5 min 后用蒸餾水稀釋至10 mL,以粗酶液加磷酸緩沖液(0.1 mol/L,pH7.0)為對照,按上法測定β-D-N-乙酰氨基葡萄糖濃度,并計算幾丁質酶酶活。
幾丁質酶酶活單位定義為:在50 ℃,pH=7.0 的條件下,每分鐘催化幾丁質生成相當于l μmolβ-DN-乙酰氨基葡萄糖所需的酶量為一個酶活單位。
1.3 數(shù)據(jù)處理
實驗中每個處理重復三次,利用Design-Expert軟件設計響應面試驗并進行結果分析,采用SPSS 軟件分析數(shù)據(jù)的顯著性,利用Origin 以及Excel 對試驗數(shù)據(jù)處理分析并進行圖表制作。
2 結果與分析
2.1 培養(yǎng)條件對Bt028 菌株產幾丁質酶的影響
2.1.1 不同碳源對菌株Bt028 產幾丁質酶的影響本實驗研究了7 種不同碳源對菌株Bt028 幾丁質酶產量的影響,由圖1 可知,采用這7 種碳源發(fā)酵培養(yǎng)基所獲得的幾丁質酶酶活有著很明顯的差異。其中以幾丁質作為碳源時,菌株Bt028 產幾丁質酶的活力遠高于其他碳源;同濃度的膠體幾丁質誘導菌株Bt028 產幾丁質酶的性能又優(yōu)于粉末幾丁質,其余5 種碳源培養(yǎng)基所得到的發(fā)酵液酶活明顯不如前者,故表明膠體幾丁質是誘導Bt028 產幾丁質酶的最佳碳源。

圖1 碳源種類對幾丁質酶合成的影響Fig.1 Effects of carbon sources on synthesis of chitinase注:不同小寫字母代表差異顯著(P<0.05);圖2~圖11 同。
2.1.2 膠體幾丁質濃度對菌株Bt028 產幾丁質酶的影響 以膠體幾丁質為碳源,考察其濃度對菌株Bt028 產幾丁質酶的影響。圖2 結果顯示,隨著膠體幾丁質濃度的增加酶活力逐漸上升,在濃度為1%時達到最高,與其他膠體幾丁質濃度下幾丁質酶活性存在顯著性差異(P<0.05)。在此基礎上增加培養(yǎng)基中幾丁質的濃度,產酶能力反而下降。這說明Bt028不僅對碳源的種類有著專一的要求,而且對碳源的濃度也比較嚴格,在膠體幾丁質濃度小于0.5%的條件下,產酶能力迅速降低。當膠體幾丁質濃度高于1%,隨著其濃度的增加,酶活不斷降低。幾丁質含有的大量羥基和氨基具有很強的吸附能力[34],當濃度過高時酶會被吸附在幾丁質上從而降低了離心后上清液的酶活。

圖2 膠體幾丁質濃度對幾丁質酶合成的影響Fig.2 Effect of colloidal chitin concentration on the synthesis of chitinase
2.1.3 氮源種類對菌株Bt028 產幾丁質酶的影響細菌可利用的氮源可分為有機氮和無機氮。將Bt028菌種液接種于含不同氮源的產酶培養(yǎng)基中,測定幾丁質酶活,結果如圖3 所示,添加的幾種有機氮源酵母浸粉、蛋白胨、酵母粉均能提高產酶能力,其中酵母粉的效果最明顯,銨鹽、硝酸鹽等無機氮源則使產酶能力有不同程度下降,故酵母粉這種有機氮源更容易被利用,從而提高產酶能力。從酶活的大小來看,有機氮(酵母浸粉、蛋白胨、酵母粉)明顯優(yōu)于無機氮(硫酸銨、氯化銨、硝酸銨、硝酸鈉),表明菌株Bt028 幾丁質酶合成對氮源的種類具有選擇性。

圖3 氮源種類對幾丁質酶合成的影響Fig.3 Effects of nitrogen sources on chitinase synthesis
2.1.4 酵母粉濃度對菌株Bt028 產幾丁質酶的影響 以酵母粉為氮源,考察其濃度對菌株Bt028 產幾丁質酶的影響,結果如圖4 所示。結果表明,當酵母粉濃度為1%時,菌株Bt028 發(fā)酵液中幾丁質酶酶活最高。當?shù)礉舛雀哂?%時,隨著底物濃度的升高使得溶液的粘稠度增加,影響酶的作用,因此酶活迅速降低,故酵母粉濃度以1%為最佳。
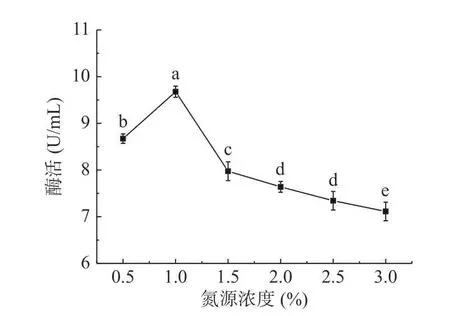
圖4 酵母粉濃度對幾丁質酶合成的影響Fig.4 Effect of yeast powder concentration on chitinase synthesis
2.1.5 培養(yǎng)基初始pH 對菌株Bt028 產幾丁質酶的影響 微生物各項生命活動都與酶反應息息相關,pH直接影響各種酶活力,在最適pH 條件下,才可能發(fā)揮最佳酶活。將Bt028 菌株接種于初始pH 不同的產酶培養(yǎng)液中,測定酶活力,結果見圖5。當培養(yǎng)基pH 為5.5 時,與pH7.0 時酶活相比,幾丁質酶酶活顯著降低(P<0.05);當pH 大于5.5 時,酶活不斷提高,pH 為7.0 時,幾丁質酶酶活最高,且與其他pH 下幾丁質酶活存在顯著性差異(P<0.05)。結果表明菌株Bt028 產酶最佳的初始pH 為7.0,該菌株適合在中性環(huán)境下生長代謝。

圖5 初始pH 對幾丁質酶合成的影響Fig.5 Effect of initial pH on chitinase synthesis
2.1.6 培養(yǎng)溫度對菌株Bt028 產幾丁質酶的影響發(fā)酵溫度主要影響細胞產生酶的活力,溫度過高或過低都會降低酶活;只有發(fā)酵溫度適宜時,菌株才會最大限度利用培養(yǎng)基中營養(yǎng)物質。將Bt028 菌株接種于產酶培養(yǎng)基中,分別在不同溫度下培養(yǎng),測定酶活力。結果見圖6,當培養(yǎng)溫度為20~30 ℃時,酶活隨溫度的升高而增加;當溫度高于30 ℃時,酶活開始急劇下降。因此,當培養(yǎng)溫度達到30 ℃左右時,菌株生長代謝所產生的幾丁質酶活性最高,且與其他幾個溫度下幾丁質酶酶活顯著性差異(P<0.05)。

圖6 溫度對幾丁質酶合成的影響Fig.6 Effect of temperature on chitinase synthesis
2.1.7 接種量對菌株Bt028 產幾丁質酶的影響 如圖7 所示,當接種3%菌種液時,酶活力最高為10.11 U/mL;在接種量低于3%時,隨著接種量的升高,幾丁質酶活不斷提升;當接種量大于3%時酶活下降,是由于接種量過大導致瓶內菌株大量繁殖,過快消耗瓶內營養(yǎng)物質和氧氣,導致菌體停止合成分泌幾丁質酶甚至死亡,使酶活降低,因此最佳接種量為3%。

圖7 接種量對幾丁質酶合成的影響Fig.7 Effect of inoculation amount on chitinase synthesis
2.1.8 裝液量對菌株Bt028 產幾丁質酶的影響 圖8表明,當裝液量為50~100 mL/250 mL時,菌株產幾丁質酶酶活緩慢上升,到100 mL/250 mL達到最高,這是因為裝液量較少,發(fā)酵液中溶氧量充足;但當裝液量超過100 mL/250 mL 時,搖瓶內溶氧不足,從而使菌株代謝受到抑制,發(fā)酵液活性反而降低,由此說明最適裝液量為100 mL/250 mL。

圖8 裝液量對幾丁質酶合成的影響Fig.8 Effect of liquid loading on chitinase synthesis
2.1.9 搖床轉速對菌株Bt028 產幾丁質酶的影響搖床轉速影響到發(fā)酵液中氧氣的溶解量以及底物、產物的傳質過程。如圖9 所示,搖床轉速在150~200 r/min 范圍內,酶活力呈現(xiàn)先升高后減小,在轉速達到180 r/min 時,測得的酶活力達到最高。當搖床轉速在100~150 r/min 時,酶活力隨著轉速的增加不斷升高,轉速低導致培養(yǎng)液中的溶氧率太低,而高轉速提供的通氣量能較好地滿足菌體生成的需求;當轉速高于180 r/min 時,雖然培養(yǎng)液中的通氣量得到提升,但過高的振蕩速率會菌體產生影響,不利于菌體和底物的接觸,影響了菌株的生長,從而使得菌株產酶能力的降低,故Bt028 的最優(yōu)搖床轉速為180 r/min。

圖9 搖床轉速對幾丁質酶合成的影響Fig.9 Effect of table speed on chitinase synthesis
2.1.10 培養(yǎng)時間對菌株Bt028 生長和產幾丁質酶的影響 圖10(a)為條件優(yōu)化后的菌株生長曲線,結果表明培養(yǎng)36 h 內菌株處于對數(shù)生長期,菌體濃度迅速提高并達到最大值;在36~72 h,由于培養(yǎng)基中營養(yǎng)物質消耗、代謝產物積累等不利因素的影響,細菌繁殖速度漸趨下降,故菌體生長曲線在36~72 h 時逐漸下降,在72 h 后趨于穩(wěn)定。菌株Bt028 培養(yǎng)過程中酶活隨時間的變化如圖10(b)所示。在前36 h,Bt028 處于對數(shù)生長期,該時期主要是菌株利用營養(yǎng)成分進行繁殖生長,幾丁質酶產量較少;在36~48 h期間,隨著菌體的快速增殖以及培養(yǎng)基中營養(yǎng)成分的減少,菌株開始迅速分泌幾丁質酶并在48 h 左右達到最大,之后酶活力開始下降并趨于穩(wěn)定。因此菌株Bt028 產幾丁質酶的培養(yǎng)時間確定為48 h。

圖10 Bt028 生長曲線和產酶曲線Fig.10 Growth curve and enzyme production curve of Bt028注:a 為Bt028 生長曲線;b 為Bt028 產酶曲線。
2.2 菌株Bt028 產幾丁質酶發(fā)酵條件的響應面優(yōu)化
2.2.1 回歸模型建立及方差分析 根據(jù)單因素實驗中對菌株Bt028 產幾丁質酶影響相對較大的3 個因素:膠體幾丁質的濃度、培養(yǎng)溫度和培養(yǎng)基初始pH 對產酶培養(yǎng)條件進行優(yōu)化。以膠體幾丁質濃度(A)、培養(yǎng)溫度(B)、培養(yǎng)基初始pH(C)3 因素為自變量,以幾丁質酶活力為響應值,設計3 因素3 水平共17 個實驗點,分為12 個析因點和5 個零點。試驗因素水平選取見表1,實驗設計與結果如表2 所示。

表2 Box-Behnken 試驗設計及結果Table 2 Design and results of Box-Behnken tests
該響應面模型的顯著性檢驗和方差分析結果(見表3)顯示,該回歸模型的P值<0.01,而失擬項的P值>0.05,證明模型建立成功;從表中P值可以看出其自變量B 對響應值的影響顯著(P<0.05),自變量A、C、BC、A2、B2、C2對響應值影響極顯著(P<0.01),AB、AC 影響不顯著(P>0.05);回歸系數(shù)R2=99.65%大于90%,校正決定系數(shù)R2Adj=0.9921>0.9,表明模型相關度很好;本試驗的變異系數(shù)為1.03%,說明模型擬合較好;本模型精密度為45.594>4.0,說明膠體幾丁質含量、培養(yǎng)溫度、培養(yǎng)基初始pH 對產幾丁質酶活力影響的試驗數(shù)據(jù)可靠,此模型可用來分析和預測實驗結果。各因素對蘇云金芽孢桿菌Bt028 產幾丁質酶酶活力影響的二次回歸模型編碼方程為:
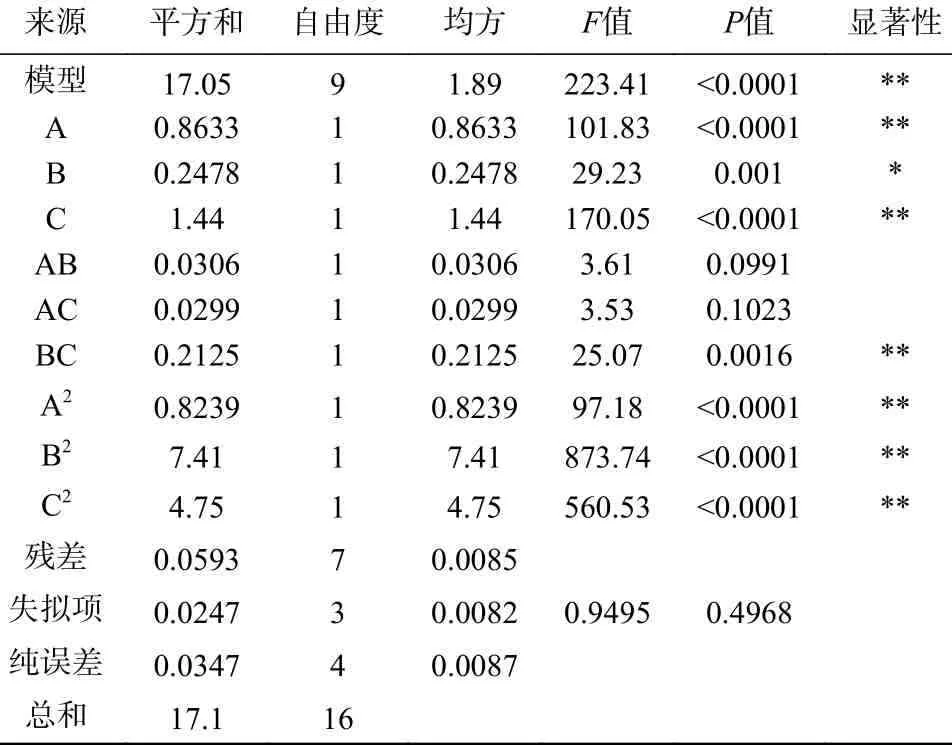
表3 回歸模型方差分析Table 3 Variance analysis of regression model
酶活力(U/mL)=10.40?0.33A?0.18B+0.43C+0.09 AB+0.09AC+0.23BC?0.45A2?1.34B2?1.072C2。
2.2.2 響應曲面分析 對各因素對幾丁質酶活力的交互影響進行響應面分析,結果見圖11~圖13。響應面圖表明了實驗因素對實驗結果的影響,由圖11~圖13 可以看出,三個因素兩兩之間的交互作用響應面曲面均呈凸起趨勢,說明存在最大響應值。運用Design-Expert12.0 軟件對其結果進行分析,預測最佳發(fā)酵條件為膠體幾丁質0.839%、培養(yǎng)溫度29.615 ℃、培養(yǎng)基初始pH6.9155,幾丁質酶活力理論值為10.50 U/mL。

圖11 膠體幾丁質濃度和溫度交互作用對幾丁質酶活影響的曲面圖及等高線Fig.11 Response surface polt and contour polt showing the effects of interactions of colloidal chitin concentration and temperature on chitinase activity

圖12 溫度和pH 交互作用對幾丁質酶活影響的曲面圖及等高線Fig.12 Response surface polt and contour polt showing the effects of interactions of temperature and pH on chitinase activity

圖13 膠體幾丁質濃度和pH 交互作用對酶活力影響的曲面圖及等高線Fig.13 Response surface polt and contour polt showing the effects of interactions of colloidal chitin concentration and pH on chitinase activity
2.2.3 驗證試驗 在考慮到實際實驗操作的基礎上,對模型的準確性進行驗證,將軟件分析得出的最佳發(fā)酵條件進行調整:膠體幾丁質0.8%、酵母粉1%、發(fā)酵溫度30 ℃、初始pH6.9、轉速為180 r/min、裝液量100 mL/250 mL、接種量3%、發(fā)酵時間48 h,3 次平行實驗所得幾丁質酶的平均酶活為10.48±0.28 U/mL,與理論值10.50 U/mL 接近,表明響應面法所得結果準確可靠。
3 結論
本實驗通過單因素實驗和響應面試驗最終確定了Bt028 產酶的最適培養(yǎng)基組成為:膠體幾丁質0.8%、酵母粉1%;最適產酶條件為初始pH 為6.9、接種量3%、裝樣量100 mL/250 mL、溫度30 ℃、培養(yǎng)時間48 h。在此條件下,該菌株酶活最高可達到10.48 U/mL,較篩選時的3.56 U/mL 提高了1.94 倍。通過實驗觀察發(fā)現(xiàn),本文篩選的Bt028 菌株從培養(yǎng)24 h 后開始大量產酶,在48 h 時達到最大值,說明該菌株有良好的產酶性能,具有較高的產酶效率,應用潛力較大,為幾丁質酶的生產應用奠定了良好的基礎。本文僅對該菌株的實驗室搖瓶培養(yǎng)產酶條件進行了研究,對于該菌株產幾丁質酶的工業(yè)生產工藝、酶的分離純化、酶的催化特性及其在食品、農業(yè)等領域的應用等還有待于進一步深入研究。
猜你喜歡
中學生數(shù)理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現(xiàn)代檢驗醫(yī)學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
