基于轉錄組的荔枝泛素結合酶基因家族的鑒定及表達分析
2022-06-02 15:12:43董晨李金枝鄭雪文王弋李偉才
熱帶農業科學 2022年5期
關鍵詞:分析
董晨 李金枝 鄭雪文 王弋 李偉才

摘 ?要:泛素結合酶在泛素化級聯反應中居于中心位置,是連接泛素活化酶和泛素連接酶的橋梁,在泛素化系統中發揮著關鍵作用。本研究基于荔枝轉錄組數據庫,利用生物信息學方法對轉錄組數據庫泛素結合酶UBC基因家族進行鑒定,最終得到28個LcUBC基因家族成員。通過熒光定量PCR技術對LcUBC基因家族在‘妃子笑荔枝不同組織中進行時空表達分析,并對花穗對照和烯效唑處理花穗發育的不同時期進行表達分析。結果表明:LcUBC基因家族成員對應所編碼的氨基酸數分布在85~1146 aa之間,等電點大小在4.03~9.61之間,5個LcUBC蛋白為穩定蛋白,其余均為不穩定蛋白。2個LcUBC蛋白定位于細胞質,2個LcUBC蛋白定位于細胞質和細胞核,1個LcUBC蛋白定位于內質網,其余蛋白均定位于細胞核。系統進化分析結果表明,LcUBC蛋白分為10個亞家族;28個LcUBC基因家族成員在不同組織的表達量存在差異;烯效唑處理‘妃子笑荔枝花穗與對照相比,烯效唑處理21?d后,LcUBC基因家族成員的多個基因表達量較高。這是在轉錄組水平分析LcUBC基因家族成員,本研究結果對今后該基因家族的分類、克隆和功能研究提供了參考依據。
關鍵詞:荔枝;泛素結合酶;基因家族;表達分析中圖分類號:S667.1??????文獻標識碼:A
Transcriptome-wide Identification and Analysis of UBCGene?Family in Litchi chinensis?Sonn.
Abstract: Ubiquitin-conjugating enzymes play a central role in the ubiquitination cascade. They are the bridge between ubiquitin activating enzymes and ubiquitin ligases, and play a key role in the ubiquitination system. Through the local BLAST search and keyword search of transcriptome database, 32 ubiquitin-conjugating enzyme gene family members, including 5UEVand 28UBCgenes, were obtained through smart screening of conserved domains. OnlyLcUBC1–LcUBC28was studied in the article. Based on the litchi transcriptome database, the ubiquitin-conjugating enzyme gene family was identified by bioinformatics methods, and the temporal and spatial expression of ubiquitin-conjugating enzyme gene family in different tissues and organs of Feizixiao litchi was analyzed by the fluorescence quantitative PCR technology. Meanwhile, the expression of the control and uniconazole treatment at different stages of flower development was analyzed. The results showed that the amino acids were 85–1146?aa. The isoelectric point ranged?from 4.03 to 9.61. Five LcUBC proteins were?stable, and the others?LcUBC proteins were?unstable. Two LcUBC proteins were located in cytoplasm, two LcUBC proteins?in cytoplasm and nucleus, one?LcUBC protein in endoplasmic reticulum and the rest LcUBC proteins?in nucleus. There were?great differences in the length of the protein and the changes in the characteristics of the protein, indicating that the proteins of the ubiquitin protease gene family had?different characteristics. In the multiple sequence alignment, all 28?LcUBC?genes contained an evolutionarily highly conserved amino acid residue cysteine active site, and the C-terminal tryptophan (W) located behind the cysteine active site.?The 28 LcUBC proteins?were also highly conserved, and the “HPNI” conserved motif was located 7–10 amino acids before the active site of cysteine. The?LcUBC?gene?family was predicted to contain 10 conserved motifs. There were?certain differences in the number and types of conserved motifs contained in the members of the LcUBC?gene family. Among them, the conserved motifs 1 and 2 were?very important conserved motifs in the LcUBC domain. Phylogenetic analysis showed that the?litchi?UBC protein was divided into 10 subfamilies, namely UBC1, UBC2/11, UBC3/7, UBC4/5, UBC6, UBC8, UBC10/12, UBC13, UBC14, UBC15.?The 28?LcUBCgenes were?expressed in at least one tissue, and the expression levels of different genes in different tissues?were?different. Among them, the expression ofLcUBC4,?LcUBC5, andLcUBC18in seeds?were?significantly higher than that of other tissues, and the expression of?LcUBC7 in male flowers was significantly higher than that of other tissues. The expression level of female flowers ofLcUBC14andLcUBC15was significantly higher than that of other tissues, andLcUBC10was highly expressed in pericarp and old leaves. Compared with the control, the expression levels of LcUBC?genes in different developmental stages of Feizixiao litchi flower spikes treated with uniconazole were higher at 21 days after the treatment with Uniconazole. This is the first time that the?litchi?UBC?gene family was?analyzed at the transcriptome level. The results of the study would?provide some referrences for future studies on the classification, cloning and function of this gene family.30975874-6D9F-44F5-991C-F8B3C587AB3B
Keywords: Litchi chinensisSonn.; ubiquitin-conjugating enzyme; genes family; expression analysis
DOI: 10.3969/j.issn.1000-2561.2022.05.002
荔枝(Litchi chinensis Sonn.)是熱帶、亞熱帶發展中國家的最重要的作物之一。目前荔枝產業已成為中國華南熱作產業的支柱產業之一,對中國南方農村經濟的發展和熱區農民收入水平的提高發揮了舉足輕重的作用。2018年是我國有史以來最豐產的一年,據不完全統計,全國荔枝種植面積約為54.2萬hm2,產量約為301萬t,種植面積排在蘋果、柑橘、梨、葡萄和桃之后,屬于大宗果樹,但從總產量來看卻還算“小水果”。2018年荔枝結果面積按照總種植面積的80%計算,平均單產不足7.0 t/hm2[1]。‘妃子笑具有花穗長、花量大、坐果率低的特點,坐果率低是其生產中亟待解決的一個“瓶頸”問題,調控坐果的分子機理是當今果實發育領域研究的一個熱點問題。
泛素-蛋白酶體途徑(ubiquitin-proteasome pathway,UPP)廣泛存在于真核生物中,并且在維持細胞功能、細胞周期運轉、抵御環境脅迫、胚胎發育、激素響應和衰老等方面發揮著重要的作用[2-5]。在泛素化過程中,關鍵酶主要包括泛素活化酶(E1)、泛素結合酶(E2)和泛素-蛋白連接酶(E3)。編碼E2及E2-like的基因已報道37個,根據結構和序列的相似性可分為12個亞家族或14個組,E2-like基因8個[6]。E2蛋白的典型特征是包含一個長約140~200 aa的保守催化結構域——UBC結構域,內含有1個高度保守的半胱氨酸活性位點,另外還存在一類UEV(ubiqutin E2 variants)蛋白,該蛋白家族在序列和結構上與泛素結合酶相似,但是缺少半胱氨酸催化位點,其功能也與典型的泛素結合酶蛋白有所不同。E2多以基因家族的形式存在,參與許多植物生理活動[7]。泛素結合酶在泛素化級聯反應中居于中心位置,是連接泛素活化酶和泛素連接酶的橋梁,在泛素化系統中發揮關鍵的作用。
目前,對E2基因的研究多集中在對DNA的修復進程。如在核酸修復途徑中,UBC35/UBC36和 Mms2p/Uev1a蛋白形成復合物,在賴氨酸位點催化形成泛素鏈,來引發核酸修復的信號轉導,完成對受損核酸的修復[8]。UBC13協同E3連接酶RNF 8相互作用形成復合物,DNA損傷信號通過依賴泛素化的信號途徑來傳遞至染色質,引發染色質蛋白的磷酸化,促使RNF 8-UBC 13復合物定位到受損傷的核酸部位,產生泛素鏈后觸發DNA修復信號通路,使RAP 80蛋白和整個Brca1A復合物定位到受損傷的DNA部位,對損傷的DNA進行修復[9]。另外還有研究表明RNF 8-UBC 13復合物可獨立的行使底物蛋白的泛素化,并促進DNA損傷的調節子53BP1蛋白的積累[10]。
泛素結合酶在植物衰老和果實發育過程中起重要作用。PICTON等[11]首次從番茄中克隆到泛素結合酶基因,研究發現泛素結合酶基因在番茄葉片衰老和果實成熟過程中表達。香蕉采后早期抑制差減文庫中篩選得到香蕉中的泛素結合酶基因MaUCEI,該基因介導細胞內小的調節蛋白的降解,與酵母中的UBC 4/UBC 5結合,推測該基因可能通過影響果實成熟過程中淀粉酶的活性或含量而參與對淀粉降解的調控,在果實成熟衰老的分解代謝中發揮作用[2]。2013年日本學者研究表明,大豆近等基因系成熟基因座等位基因E2/E3對基因型的修飾可以提供新品種,與對照相比,新品種花期較晚且產量高,可以適應不同的緯度環境,并可保留原來的種子質量[12]。WANG等[13]研究發現,番茄中2個E2基因(SlUBC 32和SlUBC 41)參與調控番茄果實成熟,但具體分子調控機制尚不清楚。DONG等[14]研究香蕉花后果實不同發育階段E2基因家族的表達情況,結果發現E2基因家族不同成員在果實發育不同階段行使功能,推測這些基因在果實發育過程中發揮關鍵作用。陳曙等[15]研究低磷處理下玉米幼苗不同組織中E2基因的表達情況,結果發現ZmUBC 17基因具有組織表達特異性,在葉片中大量表達,推測ZmUBC 17基因是通過調控玉米對磷元素吸收和轉運間接影響光合作用效率。
通過對擬南芥基因組數據庫搜索顯示,E2基因以多拷貝形式存在,造成E2蛋白功能冗余,這可能是使E2基因的研究進展不大的主要原因[7]。近年來,隨著基因組測序技術的不斷提高,越來越多的植物完成全基因組的測序工作,越來越多的植物物種中泛素結合酶基因家族在基因組學上得到鑒定,如玉米[16]、可可[17]、水稻[18]、葡萄[19]、普通油茶[20]和香蕉[14]等。目前,關于荔枝基因家族的研究有XTH[21]、ARF[22-23]基因家族,關于荔枝泛素結合酶基因家族僅僅研究了LcUBC12基因[24]。30975874-6D9F-44F5-991C-F8B3C587AB3B
本研究基于荔枝轉錄組數據庫,利用生物信息學方法對轉錄組數據庫UBC基因家族進行鑒定,并通過熒光定量PCR(qPCR)技術對LcUBC基因家族在‘妃子笑荔枝不同組織中進行時空表達分析,并對花穗對照和烯效唑處理花穗發育的不同時期進行表達分析,挖掘關鍵的LcUBC基因家族成員,為今后LcUBC基因功能的研究奠定基礎。本研究結果將為從蛋白質降解方面深化荔枝坐果機理的研究和生產上調控荔枝坐果技術措施提供理論依據。
1 ?材料與方法
1.1 材料
供試品種為‘妃子笑荔枝,取自廣東省湛江市麻章區湖光鎮湖秀路一號南亞熱帶作物研究所荔枝栽培示范園,荔枝UBC蛋白序列來源于本課題組前期構建的‘妃子笑花穗發育轉錄組數據庫(GenBank accession SRP092890)。
1.2 方法
1.2.1 ?LcUBC基因家族的鑒定與分析??利用模式植物擬南芥和水稻UBC序列作為探針序列,運用本地Blast軟件對荔枝測序轉錄組數據庫中的unigene進行搜索;同時,利用關鍵詞“Ubiquitin-?conjugating enzymes”和“UBC”直接檢索,得到所有可能的UBC序列后,對檢索結果進行整合分析,去除重復序列后,利用Pfam在線軟件對序列保守結構域進行鑒定,去除不含LcUBC基因家族保守結構域的序列。
1.2.2 ?LcUBC蛋白的基本理化性質、序列比對和進化分析??運用ProtParam在線軟件(http://web.?expasy.org/protparam/)分析預測LcUBC蛋白序列的分子量、等電點、不穩定系數、脂肪指數和疏水性等基本的理化性質;采用Plant-mPLoc Server在線軟件(http://www.csbio.sjtu.edu.cn/bioinf/?plant-multi/#)分析LcUBC蛋白的亞細胞定位;運用MEME?4.12.0在線軟件(http://meme.nbcr.net/?meme/)分析LcUBC家族蛋白的保守基序,參數設置:基序的最大數目設置為10, 基序長度設為6~150個氨基酸,其他參數為默認值。通過MEGA?6.0的MUSCLE程序比對氨基酸多重序列,參數為程序默認參數。利用多重序列比對產生的LcUBC保守結構域145個氨基酸殘基后續進行NJ(Neighbor-Joining)分析。采用MEGA?6.0軟件構建進化樹,Bootstrap值設為1000次重復,以獲得更為可靠的分支聚類,其他參數為默認參數。
1.2.3 ?LcUBC基因家族在花穗調控下的表達特征分析??選擇樹齡相同、長勢相近的‘妃子笑荔枝樹,分別選取荔枝樹的不同組織如嫩葉、根、莖、老葉、雌花、雄花、果肉、果皮、種子,液氮速凍后存放于–80℃冰箱中,用于RNA的提取。
花穗處理:選擇樹齡相同、長勢相近的‘妃子笑荔枝樹,當花穗長度抽生至約18?cm時,進行如下操作:(1)對照組(CK),不做任何處理;(2)烯效唑(Un)處理,用50?mg/L烯效唑(品牌:四川國光,有效成分含量5%)噴施,噴至葉面滴水;分別取對照及烯效唑處理后0、7、14、21、28、35、42?d的花穗,液氮速凍后存放于–80℃冰箱中,用于RNA的提取。
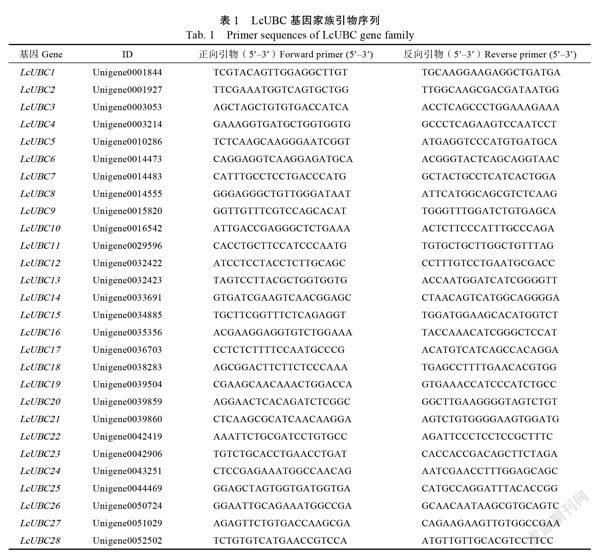
選用前期課題組篩選的LcActin為內參,根據轉錄組中注釋的LcUBC的核苷酸序列,利用在線軟件Primer?3?plus設計熒光定量引物(表1),通過Blast分析引物的特異性,引物序列委托廣州艾基生物技術有限公司合成。將處理好的材料按照RNA提取標準進行磨樣,采用華越洋生物科技有限公司RAN提取試劑盒QK型,分別對根、莖、嫩葉、老葉、雌花、雄花、果肉、果皮、種子和花穗的總RNA進行提取,提取1?μL檢測RNA的質量和濃度,樣品符合要求后利用M-MLV逆轉錄酶(TaKaRa)反轉錄合成cDNA。qPCR分析采用SYBR Premix Ex Taq?(TaKaRa)試劑盒,Roche480Ⅱ定量PCR系統(瑞士),采用384孔板,qPCR反應體系為10?μL,其中cDNA?1?μL(相當于25?ng總RNA),10??mol/L基因特異性引物1??L,2×SYBR Premix ExTaq?5?μL,最后用ddH2O補足10?μL。qPCR反應條件:50℃預變性2?min,95℃預變性2?min;95℃變性15?s,56℃退火15?s,72℃延伸30?s,共55個循環。每個樣品3個重復。采用2–DDCT法計算基因相對表達量。
2 ?結果與分析
2.1 LcUBC基因家族的鑒定
首先對轉錄組數據庫的本地Blast進行比對檢索和關鍵詞搜素,再通過SMART對保守結構
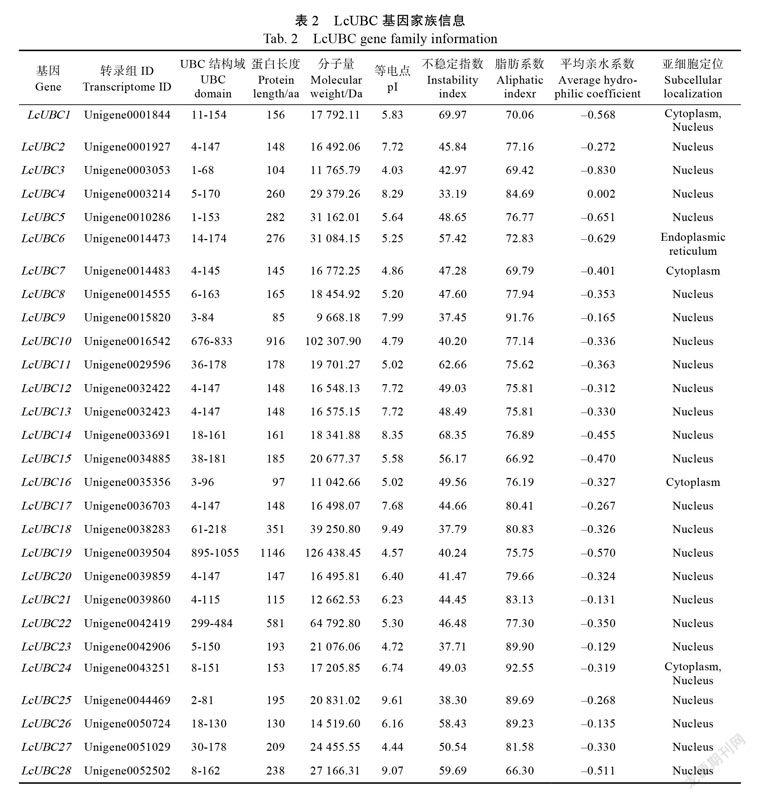
域進行篩選,最終得到28條LcUBC蛋白序列。為了便于研究,按照轉錄組中的基因ID大小前后順序統一為LcUBC蛋白編號為LcUBC 1~LcUBC 28(表2),對應所編碼的氨基酸數分布在85~ 1149?aa之間;預測的理論分子量大小范圍在9.668~102.307?kDa之間。等電點大小范圍在4.03~9.61之間,其中 18個LcUBC蛋白的等電點小于7,顯酸性;10個LcUBC蛋白等電點大于7,顯堿性;LcUBC蛋白平均等電點小于7,表明UBC蛋白為弱酸性,在酸性的亞細胞環境中發揮作用。不穩定指數分析結果發現,5個LcUBC蛋白(LcUBC?4、LcUBC 9、LcUBC 18、LcUBC 23和LcUBC 25)的不穩定指數小于40,為穩定蛋白,其余均為不穩定蛋白。平均親水系數結果表明,除了LcUBC 4蛋白疏水,其余均為親水蛋白。亞細胞定位分析結果表明,LcUBC 7和LcUBC 16蛋白定位于細胞質,LcUBC 1和LcUBC 24蛋白定位于細胞質和細胞核,LcUBC 6蛋白定位于內質網,其余蛋白均定位于細胞核。綜合上述分析結果,無論從氨基酸序列的長度還是蛋白的特性變化分析,LcUBC蛋白都有很大差異,這表明LcUBC基因家族蛋白具有不同特性。30975874-6D9F-44F5-991C-F8B3C587AB3B
2.2 LcUBC基因家族蛋白保守基序分析
通過多重序列比對LcUBC基因家族的氨基酸序列全長,比較不同LcUBC家族成員間UBC保守結構域的特點(圖1),多重序列比對中,28個LcUBC基因均含有進化上高度保守的氨基酸殘基半胱氨酸的活性位點,位于半胱氨酸活性位點后的C-端色氨酸(W)在28個LcUBC蛋白中也高度保守,“HPNI”保守基序位于半胱氨酸活性位點之前的7~10位氨基酸。
通過在線軟件MEME分析LcUBC蛋白中的保守基序,結果見圖2。預測LcUBC基因家族中含有10個保守基序,LcUBC基因家族成員之間所包含的保守基序數目及種類存在一定的差異。在LcUBC基因家族中有25個成員含Motif?1基序,有15個成員含Motif 2基序。通過SMART分析Motif?1、Motif 2為LcUBC蛋白結構域中非常重要的保守基序。
為進一步了解LcUBC基因家族成員之間的進化關系,利用模式物種擬南芥中的37個AtUBC和酵母中的13個ScUBC同荔枝中的28個LcUBC構建系統進化樹(圖3),結果表明,荔枝中的28個LcUBC分為10個亞家族,分別為UBC1、UBC2/11、UBC3/7、UBC4/5、UBC6、UBC8、UBC10/12、UBC13、UBC14、UBC15,其中UBC14和UBC15在酵母基因組中缺失,推測UBC14和UBC15是植物中特有的或是酵母進化過程中丟失。
2.3LcUBC基因家族時空表達分析
UBC參與DNA的損失修復、生長、發育及脅迫響應等方面,為了研究LcUBC基因家族在不同發育過程中的功能,分析9個不同組織(如嫩葉、嫩莖、果皮、種子、根、雄花、老葉、雌花、果肉)中LcUBC基因的表達情況(圖4)。在植物正常的發育過程中,不是所有的UBC基因在不同組織部位都表達,28個LcUBC基因家族成員中至少在某一個組織部位表達(圖5),推測這些基因有可能是在生物脅迫或非生物脅迫條件下表達。其中,LcUBC4、LcUBC5、LcUBC18在種子中的表達顯著高于其他組織,LcUBC7在雄花中的表達上顯著較高于其他組織,LcUBC14、LcUBC15在雌花中的表達量顯著高于其他組織,LcUBC10在果皮和老葉中高表達。
2.4LcUBC基因家族在花穗調控表達模式
利用qPCR分析LcUBC基因家族在荔枝花穗調控的表達量(圖5),結果表明,與對照相比,LcUBC基因家族在烯效唑處理后表達量存在顯著變化,經烯效唑處理后,LcUBC27在7、14、21、28、35 d表達量上調,LcUBC2、LcUBC7、LcUBC12、LcUBC 13、LcUBC15、LcUBC16、LcUBC17、LcUBC28在21?d表達量最高,LcUBC 20、LcUBC21在35?d表達量最高,LcUBC16表達呈現先上升后下降的趨勢,在21?d表達量最高。LcUBC12、LcUBC 23、LcUBC24、LcUBC25與對照相比表達有變化,但差異不顯著。
13、LcUBC15、LcUBC16、LcUBC17、LcUBC28在21?d表達量最高,LcUBC 20、LcUBC21在35?d表達量最高,LcUBC16表達呈現先上升后下降的趨勢,在21?d表達量最高。LcUBC12、LcUBC 23、LcUBC24、LcUBC25與對照相比表達有變化,但差異不顯著。
3??討論
本研究參考擬南芥和水稻UBC家族基因的信息,基于烯效唑處理荔枝花穗轉錄組數據,利用生物信息學方法全面分析鑒定LcUBC基因家族。結果共鑒定出28個LcUBC基因家族成員,并對其基本理化性質、亞細胞定位進行了分析。通過對LcUBC基因家族蛋白保守結構域分析可看出,LcUBC蛋白的結構域具有高度的保守性,包含典型的保守結構域UBC domain,且含有半胱氨酸活性位點,這與香蕉中UBC蛋白的分析結果一致[14]。
通過轉錄組及qPCR的結果分析,在荔枝的時空表達和烯效唑處理后的花穗調控中,大多數LcUBC基因家族的表達水平發生了變化,推測這些基因很可能通過參與激素信號轉導途徑或調控果實相關轉錄因子的水平,進而影響荔枝果實成熟過程中的各種生理變化。LcUBC 4、LcUBC 5、LcUBC8、LcUBC18、LcUBC19在種子中表達量較高,推測這5個LcUBC家族基因在果實種子發育過程中發揮著重要的作用;LcUBC 6、LcUBC14、LcUBC15、LcUBC23、LcUBC24在雌花中表達量較高,LcUBC 7、LcUBC 26在雄花中表達量較高,推測這7個LcUBC家族基因在果實花發育過程中發揮著重要的作用;LcUBC 10、LcUBC12、LcUBC18、LcUBC27在老葉中表達量較高,而LcUBC 13、LcUBC17、LcUBC20、LcUBC21在果皮中表達量較高。多個LcUBC家族基因的表達水平在烯效唑處理后第21天的表達量較高,這表明LcUBC基因家族在該時期調控荔枝花穗發育過程中發揮著關鍵作用。該研究結果為進一步研究LcUBC基因家族的生物學功能奠定了堅實的基礎。
參考文獻
[1]?李建國, 王惠聰, 周碧燕, 趙明磊, 李彩琴, 夏??瑞, 黃旭明. 荔枝花果發育生理和分子生物學研究進展[J]. 華南農業大學學報, 2019, 40(5): 119-127.LI J G, WANG H C, ZHOU B Y, ZHAO M L, LI C Q, XIA R, HUANG X M. Research advances in physiology and molecular biology of flower and fruit development in litchi[J]. Journal of South China Agricultural University, 2019, 40(5): 119-127. (in Chinese)30975874-6D9F-44F5-991C-F8B3C587AB3B
[2]?董發才, 宋純鵬. 植物細胞中的泛素及其生理功能[J]. 植物生理學報, 1999, 35(1): 54-59.DONG F C, SONG C P. The ubiquitin and its physilolgical functions in plants[J]. Plant Physiology Journal, 1999, 35(1): 54-59. (in Chinese)
[3]?覃??碧. 泛素蛋白酶體途徑及其對植物激素信號轉導的調控[J]. 熱帶農業科學, 2013, 33(4): 39-45.QI B. Ubiquitin-proteasome pathway and its regulation of plant hormone signaling[J]. Chinese Journal of Tropical Agriculture,?2013, 33(4): 39-45. (in Chinese)
[4]?王??園, 王甲水, 謝學立, 雷曉明, 金志強. 香蕉泛素結合酶基因與果實成熟關系的研究[J]. 園藝學報, 2010, 37(5): 705-712.WANG Y, WANG J S, XIE X L, LEI X M, JIN Z Q, Studies of the relationship betweenMaUCE1and banana fruit ripening[J]. Acta Horticulturae Sinica, 2010, 37(5): 705-712. (in Chinese)
[5]?盛仙永, 胡正海, 林金星. 泛素/蛋白酶體途徑及其在高等植物有性生殖中的作用[J]. 西北植物學報, 2004, 24(8): 1527-1536.SHENG X Y, HU Z H.LIN J X. Ubiquitin/proteasome pathway and its role in high plant sexual reproduction[J]. Acta Botanica Boreali-Occidentalia Sinica, 2004, 24(8): 1527-1536. (in Chinese)
[6]?Bachmair A, Novatchkova M, Potuschak T, Eisenhaber?F. Ubiquitylation in plants: a post-genomic look at a post-translational modification[J]. Trends in Plant Science,?2001, 6(10): 463-470.
[7]?王金利, 史勝青, 賈利強, 江澤平. 植物泛素結合酶E2功能研究進展[J]. 生物技術通報, 2010(4):?7-10.Wang J L, Shi S Q, Jia L Q, Jiang Z P. Progress on functions of ubiquitin-conjugating enzyme (E2) in plants[J]. Biotechnology Bulletin, 2010(4): 7-10. (in Chinese)
[8]?Hoege C, Pfander B, Moldovan G, Pyrowolakis G, Jentsch S. RAD6-dependent DNA repair is linked to modification of PCNA by ubiquitin and SUMO[J]. Nature, 2002, 419(6903): 135-141.
[9]?Bin W, Stephen J E. Ubc13/Rnf8 ubiquitin ligases control foci formation of the Rap80/Abraxas/Brca1/Brcc36 complex in response to DNA damage[J]. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(52): 20759-20763.
[10]?Huen M, Huang J, Yuan J s, Yamamoto M, Akira S z, Ashley C, Xiao W, Chen J J.?Noncanonical E2 variant-independent function of UBC13 in promoting checkpoint protein assembly[J]. Molecular & Cellular Biology, 2008, 28(19): 6104-6112.
[11]?PICTON S, GRAY J E, LOWE A, BARTON S L, GRIERSON D. Sequence of a cloned tomato ubiquitin conjugating enzyme[J].?Plant Physiology,?1993, 103(4):?1471-1472.
[12]?YAMADA T, HAJIKA M, YAMADA N, HIRATA K, OKABE A, OKI N, TAKAHASHI K, SEKI K, OKANO K, FUJITA Y, KAGA A, SHIMIZU T, SAYAMA T, ISHIMOTO M. Effects on flowering and seed yield of dominant alleles at maturity loci E2 and E3 in a Japanese cultivar, Enrei[J]. Breeding Science, 2012, 61(5): 653-660.30975874-6D9F-44F5-991C-F8B3C587AB3B
[13]?WANG Y Y, WANG W H, CAI J H, ZHANG Y R, QIN G Z, TIAN S P.Tomato nuclear proteome reveals the involvement of specific E2 ubiquitin-conjugating enzymes in fruit ripening[J]. Genome Biology,?2014, 15(12): 548.
[14]?Dong C, Hu H G, Jue D W, Zhao Q F, Chen H l, Xie J h, Jia L q. The banana E2 gene family: Genomic identification, characterization, expression profiling analysis[J]. Plant Science, 2016, 245: 11-24.
[15]?陳??曙, 趙秋芳, 陳宏良, 賈利強. 玉米泛素結合酶基因家族的生物信息學及表達分析[J]. 南方農業學報, 2018, 49(8): 1476-1483.CHEN S, ZHAO Q f, CHEN H l, JIA L q.?Bioinformatics and expression analysis of ubiquitin conjugating enzyme gene family in maize[J]. Journal of Southern Agriculture, 2018, 49(8): 1476-1483. (in Chinese)
[16]?JUE D W, SANG X L, LU S Q, DONG C, ZHAO Q F, CHEN H L,?JIA L Q. Genome-wide identification, phylogenetic and expression analyses of the ubiquitin-conjugating enzyme gene family in maize[J]. PLoS?One, 2015, 10(11): e143488.
[17]?趙秋芳, 董??晨, 決登偉, 陳宏良, 賈利強. 可可泛素結合酶基因家族的生物信息學初步分析[J]. 熱帶作物學報, 2016, 37(9): 1732-1740.Zhao Q f, Dong C, Jue D w, Chen H l, Jia L q. Bioinformatics preliminary analysis of ubiquitin conjugating enzyme gene family in cocoa[J]. Chineese Journal of Tropical Crops, 2016, 37(9): 1732-1740. (in Chinese)
[18]?劉??鑫, 張??恒, 闞虎飛, 周立帥, 黃??昊, 宋林林, 翟煥趁, 張??君, 魯國東. 水稻泛素結合酶基因家族的生物信息學與表達分析[J]. 中國水稻科學, 2016, 30(3): 223-231.LIU X, ZHANG H, KAN H F, ZHOU L S, HUANG H, SONG L L, ZHAI H C, ZHANG J, LU G D. Bioinformatic and expression analysis of rice ubiquitin-conjugating enzyme gene family[J]. Chinese Journal of Rice Science, 2016, 30(3): 223-231. (in Chinese)
[19]?高營營. 葡萄泛素結合酶E2基因家族全基因組鑒定及功能結構分析[D]. 武漢: 中國科學院大學(中國科學院武漢植物園), 2018.GAO Y Y. Genome-wide identification and function-structure analysis of ubiquitin-conjugating E2 genes family in grape[D]. Wuhan:?University of Chinese Academy of Sciences (Wuhan Botanical Garden, Chinese Academy of Sciences), 2018. (in Chinese)
[20]?林??萍, 姚小華, 曹永慶, 龍??偉, 王開良, 滕建華. 普通油茶泛素結合酶UBE2-J2的cDNA序列及蛋白質結構分析[J]. 林業科學研究, 2013, 26(6): 744-751.LIN P, YAO X H, CAO Y Q, LONG W, WANG K L, TENG J H. Characterization of a novel ubiquitin-conjugating enzyme fromCamellia oleifera[J]. Forest Research, 2013, 26(6): 744-751. (in Chinese)
[21]?董??晨, 魏永贊, 王??弋, 鄭雪文, 李偉才. 基于轉錄組的荔枝XTH家族基因的鑒定及分析[J].?分子植物育種, 2019, 17(12):?3865-3873.DONG C, WEI Y Z, WANG Y, ZHENG X W, LI W C. Transcriptome-wide identification and analysis of xyloglucan endotransglucosylase/hydrolase (XTH) family gene inLitchi chinensisSonn.[J]. Molecular Plant Breeding, 2019, 17(12):?3865-3873. (in Chinese)30975874-6D9F-44F5-991C-F8B3C587AB3B
[22]?董??晨, 魏永贊, 王??弋, 鄭雪文, 李偉才. 基于轉錄組的荔枝ARF基因家族的鑒定及表達分析[J]. 熱帶作物學報, 2017, 38(8): 1485-1491.DONG C, WEI Y Z, WANG Y, ZHENG X W, LI W C. Transcriptome-wide identification and expression profiling of the?auxin response factor (ARF) gene family inLitchi chinensisSonn.[J]. Chinese Journal of Tropical Crops, 2017, 38(8): 1485-1491. (in Chinese)
[23]?董??晨, 魏永贊, 王??弋, 鄭雪文, 李偉才. 不同處理方式的妃子笑荔枝花穗ARF基因家族的表達分析[J]. 熱帶作物學報, 2018, 39(6):?1122-1127.DONG C, WEI Y Z, WANG Y, ZHENG X W, LI W C. Expression analysis of auxin response factor (ARF) gene family inLitchi chinesis Sonn.[J]. Chinese Journal of Tropical Crops, 2018, 39(6):?1122-1127.?(in Chinese)
[24]?董??晨, 魏永贊, 王??弋, 鄭雪文, 李偉才,?石勝友. 荔枝泛素結合酶基因(LcUBC12)的生物信息學及表達特性分析[J].?南方農業學報, 2020, 51(5):?1091-1097.DONG C, WEI Y Z, WANG Y, ZHENG X W, LI W C, SHI S Y. Bioinformatics and expression characteristics analysis ofLitchi chinesisSonn.?ubiquitin-conjugating enzyme gene?(LcUBC12)[J]. Journal of Southern Agriculture, 2020, 51(5):?1091-1097. (in Chinese)30975874-6D9F-44F5-991C-F8B3C587AB3B
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
