中腦腹側被蓋區多巴胺能神經元短暫激活未能誘發小鼠自身給糖行為的重建
2022-06-13 08:28:54李佩云景漫毅
中國藥理學與毒理學雜志 2022年4期
關鍵詞:小鼠
李佩云,景漫毅,吳 寧,李 錦,宋 睿
(1.南京中醫藥大學,江蘇 南京 210023;2.軍事科學院軍事醫學研究院毒物藥物研究所,抗毒藥物與毒理學國家重點實驗室,神經精神藥理學北京市重點實驗室,北京 100850;3.解放軍總醫院藥劑科,北京 100850)
肥胖可引發多種慢性、非傳染性疾病,目前已被世界衛生組織列為威脅人類健康的十大疾病之一[1]。伴隨我國經濟的快速發展和居民生活方式的改變,肥胖發生率持續迅速升高,給我國公共衛生、社會和經濟造成嚴重危害。過量飲食是引發肥胖的重要因素之一。高糖、高脂和高蛋白等食物不僅在客觀上為機體提供充分能量,有利于保持健康和種族繁衍,并且能夠誘發機體主觀的愉悅感,而后者則是驅動機體過量飲食的原動力,類似于藥物成癮性行為[2-3]。研究發現,高復吸率是成癮治療的難點,而伴隨成癮行為的相關線索是誘發復吸的危險因素之一[4]。因此,研究線索誘發覓食重建的神經生物學機制將是探索預防和治療肥胖的有效策略。
中腦邊緣皮質多巴胺(dopamine,DA)系統作為腦內邊緣系統參與調控獎賞、學習及動機等基本生理功能,是人類趨利避害、種族繁衍的重要神經生物學基礎[5]。中腦邊緣DA系統起始于中腦腹側被蓋區(ventral tegmental area,VTA),其DA能神經纖維經內側前腦束投射至伏隔核(nucleus accumbens,NAc)、內側前額葉皮質、紋狀體和杏仁核等腦區[6-7]。該系統獎賞效應的產生主要是通過提高NAc內DA的含量,作用于NAc內的γ-氨基丁酸(γ-aminobutyric acid,GABA)能神經元,從而調節基底神經節,驅動機體做出行為反應[8]。研究發現,美食作為初級強化物主要通過激活VTA的DA能神經元,促進NAc中DA神經遞質的釋放,產生獎賞效應,驅動機體形成主動性的飲食行為[9-11]。造成過量飲食的核心特征是強迫性的覓食行為,即在主動性飲食行為消退后,當與食物相關的線索等因素出現后,機體出現強烈覓食動機,最終發生不可控的覓食行為,形成過量飲食的惡性循環[12-14]。然而VTA的DA能神經元是否同時參與調控覓食行為的重建過程,目前尚無文獻報道。在藥物成癮復吸的研究中發現,在海洛因誘導大鼠建立的條件性位置偏愛模型中,相關環境線索在誘發條件位置偏愛行為重建時可記錄到VTA腦區內DA能神經元集群的電生理信號顯著增強[15],提示VTA的DA能神經元參與了條件位置偏愛行為的重建。此外,在線索誘發小鼠可卡因成癮復吸時,采用光纖記錄系統可檢測到VTA腦區DA能神經元的特異性激活[16]。以上結果表明,在線索誘發藥物成癮復吸行為時,VTA腦區的DA能神經元可能發揮重要調節作用。
目前我國高糖飲食非常普遍,被認為是導致當前肥胖流行的原因之一。蔗糖是一種天然的能量來源和獎賞性物質,通過行為學檢測其激勵獎賞價值,甚至比可卡因等成癮性物質更高[17]。因此,口服蔗糖自身給糖行為已成為過量飲食障礙研究的經典動物模型。與此同時,隨著神經科學研究工具的跨越式發展,光遺傳學技術使得人類以往對神經元之間作用的理解由僅停留在相關性上,精確到探究特定神經回路和大腦功能之間的因果關系。據此,本研究結合小鼠自身給糖模型和光遺傳學技術,以期探討VTA腦區的DA能神經元對小鼠覓糖行為重建的調控作用。
1 材料與方法
1.1 動物
轉基因DAT-Cre小鼠〔品系:strainB6.SJL-Slc6-a3tm1.1(cre)Bkmn/J〕,8周齡,體重25~30 g,雌雄隨機,北京生命科學研究所羅敏敏教授實驗室惠贈,由本課題組繁育。小鼠在SPF級動物實驗室飼養,實驗室溫度21~23℃,濕度50%~60%,每天明暗交替各12 h(8∶00-20∶00燈光照明);訓練期間動物自由飲水,每天限食3.0 g小鼠維持飼料,實驗操作在8∶00-18∶00之間完成。所有的動物處理均符合軍事醫學研究院倫理審查委員會的要求。
1.2 試劑和主要儀器
重組腺相關病毒rAAV-EF1α-DIO-hChR2-mCherry-WPRE-pA(5.18×109g·L-1)由樞密腦科學技術有限公司提供;蛋白酶K抑制劑和2×Taq PCR MasterMix,博邁德生物公司;O.C.T.冷凍切片組織包埋劑,美國SAKURA公司;兔抗酪氨酸羥化酶(tyrosine hydroxylase,TH)多克隆抗體(批號:ab6211),美國Abcam公司;Alexa Fluor 488山羊抗兔IgG抗體和PBS緩沖液粉末,北京中杉金橋生物技術有限公司;蔗糖和多聚甲醛,國藥集團化學試劑有限公司;1454瞬干膠,北京天山新材料技術有限公司。
梯度PCR儀,德國Analytik Jena AG公司;DYY-7C瓊脂糖凝膠電泳儀,北京六一儀器廠;Fluorchem FC2凝膠成像分析系統,美國Alpha Innotech公司;小鼠腦立體定位儀和顱內導管系統(0.48 mm×0.34 mm,外徑×內徑),瑞沃德生命科學有限公司;縫合線,上海浦東金環醫療用品股份有限公司;縫合針,上海元洪醫療器械有限公司;光纖,北京首量科技有限公司;VA-I-N-473激光發射器,北京榜首科技有限公司;VLP-2000型激光功率檢測器,美國Coherent公司;CM1900型冰凍切片機和DM18型數字切片掃描與分析系統,德國Leica公司;微量注射泵,世界精密儀器商貿(上海)有限公司。
小鼠自身給糖系統(型號:Anilab SuperState Version 4.0),安來軟件儀器有限公司。該系統包括:操作性行為實驗箱(200 mm×150 mm×180 mm),箱內距底板4.5 cm的位置有左右2個鼻觸孔(直徑2 cm)。設置左鼻觸為有效鼻觸(active poke),觸發后伴隨5 s線索燈亮,同時中間孔泵出糖水;右鼻觸為無效鼻觸(inactive poke),觸發后不產生任何響應。實驗程序運行和數據采集由軟件控制完成。
1.3 腦立體定位注射和導管埋置
DAT-Cre轉基因小鼠ip給予戊巴比妥鈉70 mg·kg-1麻醉后固定在腦立體定位儀上,充分暴露顱骨頂部。用生理鹽水清洗顱骨頂部創口后,調節小鼠頭部使其保持水平。按照坐標在顱骨上打孔(前囟后側3.2 mm,中縫左側0.5 mm)。用微量注射泵吸取病毒液400 nL,垂直腦膜平面向下插入4.2 mm,以流速20 nL·min-1注入,注射后留針10 min以便病毒充分擴散和吸收。在同一位置腦膜平面向下3.7 mm處埋置導管,隨后用1454瞬干膠和牙科水泥固定導管。手術結束后,將小鼠放在電熱毯上至蘇醒,然后放回飼養間單籠飼養。
1.4 小鼠自身給糖行為實驗
自身給糖行為形成期訓練:為促進小鼠蔗糖自身給糖行為的形成,實驗前3 d小鼠開始限食(每天食物限量為3.0 g,約為正常攝食量的80%),直至訓練結束。采用固定比率1程序訓練,即當小鼠有效鼻觸1次,即可得到1次糖水泵出(10 μL,10%蔗糖)并伴隨5 s線索燈亮,然后進入5 s不應期。每天訓練2 h,小鼠鼻觸次數無上限。每天記錄小鼠體重、有效鼻觸次數、無效鼻觸次數、泵出糖水次數以及小鼠觸糖水孔的次數,訓練至連續3 d有效鼻觸次數變化百分率<10%,即達到形成期穩定操作階段(約2周),可進行消退期訓練。
消退期訓練:將獲得穩定自身給糖的小鼠放入同樣的訓練箱中,每天訓練2 h,該過程中小鼠觸有效鼻觸孔無糖水泵出和無線索燈亮。訓練至連續3 d小鼠有效鼻觸次數<10%形成期穩定操作值,即達到首次消退期穩定階段(約16 d),可進行線索點燃測試[18],其中首次消退期穩定階段的有效(或無效)鼻觸次數分別為首次消退期最后3 d的有效(或無效)鼻觸次數的平均值。
線索點燃測試[19]:將經過消退訓練的小鼠放入訓練箱中,當小鼠觸有效鼻觸孔后,鼻觸孔內會出現5 s燈光提示,但無糖水泵出;而小鼠觸無效鼻觸無任何響應。測試時間2 h,電腦自動記錄有效鼻觸和無效鼻觸次數。其中有效(或無效)鼻觸次數變化百分率(%)=線索點燃測試期有效(或無效)鼻觸次數/首次消退期穩定階段的有效(或無效)鼻觸次數×100%。
光刺激點燃:線索點燃測試結束后,小鼠進行第2次消退,訓練至連續3 d小鼠有效鼻觸次數<10%形成期穩定操作值,即達到第2次消退期穩定階段。采用頻率20 Hz、脈沖時長15 ms的激光連續給予100次激光刺激VTA的DA能神經元,立即將小鼠放入訓練箱中2 h,然后電腦自動記錄2 h內小鼠有效鼻觸和無效鼻觸次數。隨后進行第3次消退,訓練至連續3 d小鼠有效鼻觸次數<10%形成期穩定操作值,即達到第3次消退期穩定階段,采用頻率80 Hz、脈沖時長15 ms的激光連續給予100次激光刺激VTA的DA能神經元,立即將小鼠放入訓練箱中2 h,然后電腦自動記錄2 h內小鼠有效鼻觸和無效鼻觸次數。其中第2次消退期穩定階段有效(或無效)鼻觸次數為第2次消退期最后3 d的有效(或無效)鼻觸次數平均值。
1.5 免疫熒光法驗證光敏感通道蛋白表達特異性
行為實驗結束后,將小鼠麻醉,依次用生理鹽水和4%多聚甲醛溶液灌流后取腦,并進行冰凍切片,切片厚度30 μm,取包含VTA區域的冠狀腦片進行免疫熒光染色。首先用PBS配制10%的山羊血清,加入Triton X-100(終濃度0.3%)混勻作封閉液。用PBS洗去腦片上的包埋劑,用封閉液室溫封閉2~3 h。棄去封閉液,加入用封閉液稀釋的抗TH抗體(1∶300),4℃孵育24 h。用PBST漂洗3次,每次5 min,再加入用封閉液稀釋的Alexa Fluor 488山羊抗兔IgG抗體(1∶200),室溫避光孵育2 h,最后用含DAPI的封片劑封片固定。數字切片掃描與分析系統下觀察光敏感通道蛋白表達(紅色熒光)、抗TH抗體標記的DA能神經元表達(綠色熒光)和光敏感通道蛋白與DA能神經元共表達(橙色熒光)的情況。
1.6 統計學分析
實驗結果數據以±s表示,采用GraphPad Prism 6.0軟件作圖,并對數據進行統計分析。同一動物同一指標前后結果對比采用配對樣本t檢驗,同一動物線索點燃時有效鼻觸變化百分率與無效鼻觸變化百分率的比較采用t檢驗。P<0.05表示差異具有統計學意義。
2 結果
2.1 光敏感通道蛋白在DAT-Cre小鼠VTA DA能神經元的表達
在VTA腦區病毒注射部位可觀察到攜帶紅色熒光的光敏感通道蛋白(圖1A)和DA能神經元的特異性標志物TH蛋白(圖1B)表達,且光敏感通道蛋白與TH存在良好的共表達(圖1C)。提示光敏感通道蛋白在VTA的DA能神經元特異性地表達。
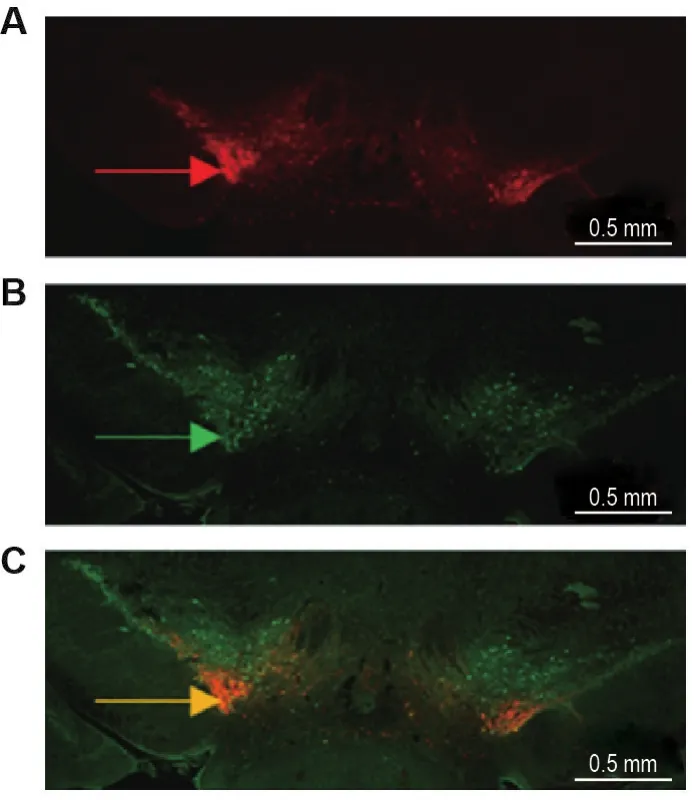
Fig.1 lmmunofluorescence expression of light sensitive channel protein in ventral tegmental area(VTA)dopaminergic neurons of DAT-Cre mice.A:the expression of channelrhodopsin 2 (ChR2).The red arrow indicates virusinfected dopaminergic neurons.B:the expression of tyrosine hydroxylase(TH).The green arrow indicates the anti-TH antibody labeled dopaminergic neurons.C:co-expression of ChR2 and TH.The orange arrow indicates the neurons in which the virus and TH antibody were co-expressed.
2.2 10%蔗糖可誘導小鼠形成自身覓糖行為
小鼠自身給糖的形成和戒斷行為檢測結果見圖2A,其中第1~14天為自身給糖形成期,第15~30天為自身給糖首次消退期,分別記錄了小鼠的有效鼻觸次數、無效鼻觸次數、泵出糖水次數(pump)和小鼠觸糖水孔次數(sucrose)。線索點燃階段檢測結果(圖2B)表明,小鼠有效鼻觸次數較首次消退期穩定階段的有效鼻觸次數顯著升高(P<0.01),無效鼻觸次數較首次消退期穩定階段的無效鼻觸次數顯著升高(P<0.01),提示燈光線索可顯著誘發小鼠覓糖復吸行為的重建,線索誘發小鼠覓糖行為重建造模成功。此外,有效鼻觸次數的變化百分率顯著高于無效鼻觸次數的變化百分率(P<0.01,圖2C),提示小鼠有意識地趨向有效鼻觸,進一步證實線索誘發小鼠覓糖行為重建造模成功。

Fig.2 10% sucrose-induced sucrose self-administration behavior in DAT-Cre mice.Mice were uniformly trained in the formation and extinction of sucrose self-administration behavior.The 1st-14thday of the training was the formation period and the 15th-30thday of the training was the first extinction period,then followed by the cue priming period.A:formation and extinction of sucrose self-administration;B:the pokes of Ext and cue priming.C:the percentage change of pokes.±s,n=13.**P<0.01,compared with the active poke during Ext;##P<0.01,compared with the inactive poke during Ext;△△P<0.01,compared with the percentage change of inactive poke.Ext means the stable stage of the first extinction period.The number of active(or inactive)pokes is the average number of active(or inactive)pokes in the last 3 days of the first extinction period.The percentage change of active(or inactive)poke(%)=active(or inactive)poke of cue priming/active(or inactive)poke of Ext×100%.
2.3 激光特異性激活VTA DA能神經元對小鼠覓糖行為重建的影響
如圖3所示,連續100次20 Hz 15 ms的激光刺激誘發的小鼠有效鼻觸次數與第2次消退期穩定階段有效鼻觸次數無顯著性差異,無效鼻觸次數也無顯著性差異。連續100次80 Hz 15 ms激光刺激誘發的小鼠有效鼻觸次數與第3次消退期穩定階段有效鼻觸次數無顯著性差異,無效鼻觸次數也無顯著性差異。提示短暫集中激活VTA單一類型的DA能神經元不能顯著誘發覓糖行為重建。
3 討論
本研究采用了小鼠蔗糖自身給糖模型,行為檢測結果顯示,與蔗糖攝入行為相關聯的燈光線索可顯著誘發小鼠的覓糖行為,提示在本實驗系統下,經過訓練的小鼠可對蔗糖產生強烈渴求。該行為特征是成癮行為的核心癥狀,因此該模型可用于小鼠自身給糖行為重建的評價。
光遺傳學技術可特異性操控特定核團特定神經元。有研究發現,將光敏感通道蛋白特異性地表達在VTA的DA能神經元中,連續給予100次20 Hz或80 Hz激光刺激后,可顯著誘發DA能神經元激活,并介導獎賞體驗[19-21],主要表現為小鼠自發活動性的升高、小鼠自身給光行為的誘導及自身給光行為隨著刺激頻率增加和刺激時間的延長而升高[19],提示正性強化作用驅動了自身給光行為的產生,而VTA釋放的DA與正性強化作用正相關。因此推測VTA DA能神經元刺激強度的增加可能與其投射靶腦區NAc內DA含量的升高呈正相關,但目前尚無文獻報道。不同頻率的光刺激VTA的DA能神經元后是否會改變NAc腦區DA的含量及其變化規律仍需深入研究。在本研究中,連續給予100次20 Hz或80 Hz的激光刺激并不能誘發小鼠覓糖行為的重建,該結果初步提示,在本實驗條件下,在自身給糖小鼠模型中,VTA單一類型DA能神經元的短暫激活不足以誘發小鼠覓糖行為的重建。
高糖、高脂和高蛋白等食物誘發機體產生的主觀愉悅感是驅動其發展為強迫性飲食及不可控覓食行為的重要基礎[3]。中腦邊緣皮質DA能神經系統是機體的獎賞中樞,調控了獎賞、動機和學習等重要生理功能,是機體趨利避害的神經結構和物質基礎。長時間大量的研究發現,蔗糖等食物誘發的獎賞也是通過上調VTA對NAc神經遞質DA的釋放而實現的[10-11],是其成為初級正性強化物的神經生物學基礎。因此,該DA能神經系統在自然獎賞過程中發揮了關鍵作用,但對于不可控的覓糖行為,該系統的作用尚不明確。本研究發現,小鼠在主動覓糖行為消退后,特異性地激活VTA中單一的DA能神經元并未能誘發覓糖行為的出現,這與本課題組前期在線索誘發毒品成癮復吸實驗的研究結果相一致[16],但與Mahler等[22]報道的采用化學遺傳學持續激活VTA的DA能神經元可誘發可卡因成癮小鼠的覓藥復吸行為結果不盡相同。二者區別之一在于,本研究所采用光遺傳學技術是在激光作用下短暫激活需要干預的特異性神經元,而Mahler等所用的化學遺傳學技術是在藥物的作用下對特異性神經元進行較長時間(氯氮平氮氧化物的作用時間可維持2 h左右)的持續干預,雖然對于神經元細胞而言僅是激活時間和強度的不同,但在其生物學效應上卻會產生很大差別。2項研究另一重要區別在于突觸間隙DA含量不同,相較于短暫的DA能神經元激活,長時間的DA能神經元激活可在突觸間隙釋放和蓄積更高濃度的DA,從而產生更為顯著的生物學變化。因此,VTA DA能神經元的持續激活誘發覓糖行為的出現值得進一步深入研究。另一方面,飲食所帶來的獎賞行為大致可分為“想要”(wanting,對獎賞的動機)和“喜歡”〔liking,獎賞的享樂價值(hedonic value)〕2類。“想要”可誘導獎賞行為,“喜歡”一般不直接誘導獎賞行為,但可增強獎賞行為。最新研究發現,小鼠腦內VTA DA能神經投射所支配的NAc核部主導了“喜歡”樣行為,干預NAc殼部的活性可延長小鼠的飲食時間,但不能增強小鼠的飲食動機,說明該核團亞區的活性改變僅能增加獎賞效應、促進獎賞行為,而不能誘發獎賞動機[23];而大量藥物成癮研究發現,作為VTA DA能神經投射支配的該核團另一亞區——NAc殼部,在獎賞動機的誘發中發揮了關鍵性作用[24-25]。而本研究在VTA腦區進行群體的DA能神經元的干預,可能會存在對不同投射腦區的DA能神經元感染效率的不同,因而造成對不同神經投射通路的刺激效率的不同,而整體表現為激活DA神經元后對小鼠的覓糖行為的重建無顯著影響。
綜上所述,同一腦區同一神經元可能介導不同的功能,包括投射到不同的腦區、投射到同一腦區支配不同的神經元和投射到同一腦區支配同一神經元的不同亞群。結合本研究結果和已有文獻報道進一步提示,對覓糖行為重建的調控,不僅關系到腦區神經元的調控作用,更需要細化到精準的神經投射通路研究,才能更為準確地揭示自身給糖行為重建的神經生物學機制。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
