生防枯草芽孢桿菌HMB19198 發酵培養基的篩選及優化
2022-06-14 09:31:18曲遠航郭慶港李社增宣立鋒張曉云
農藥學學報 2022年3期
曲遠航, 郭慶港*,, 李社增, 宣立鋒, 張曉云, 馬 平
(1. 河北省農林科學院 植物保護研究所/河北省農業有害生物綜合防治工程技術研究中心/農業農村部華北北部作物有害生物綜合治理重點實驗室,河北 保定 071000;2. 河北省農林科學院 石家莊果樹研究所,石家莊 050061)
枯草芽孢桿菌Bacillus subtilis及其近源菌種由于能產生多種抑菌活性物質,并且能形成耐逆、抗熱的芽孢而利于制劑的研制,因此成為了開發微生物源殺菌劑的重要資源[1-2]。目前防治作物土傳病害的微生物源殺菌劑制劑多為可濕性粉劑和干粉種衣劑,這些制劑加工過程中的瞬時高溫易殺死以營養體形態存在的菌體,而以芽孢形態存在的菌體則可能存活。因此,在考查微生物發酵指標時,既要考慮菌體的數量,還應考慮耐熱芽孢的濃度[3]。理想的發酵體系首先應能提高菌體的發酵水平,其次還能通過營養物質調控和環境條件的改變使菌體最大可能地轉化為耐逆性強的芽孢[4]。枯草芽孢桿菌菌體生長和芽孢的形成受多種因素影響,而培養基中的碳源和氮源是其主要影響因素,充足的碳源和氮源可促進菌體的生長,但芽孢的形成則一般發生在營養匱乏等逆境情況下[5]。由于不同菌株對適合自身菌體生長和芽孢形成所需的營養物質不同,因此,針對某一特定菌株,需要探索其特定的最適營養物質組成。
對于防治氣傳病害的微生物源殺菌劑,除了考慮菌體數量和活芽孢數量外,還要考慮其活性成分的含量。枯草芽孢桿菌類細菌產生的活性物質主要是脂肽類抗生素,包括泛革素 (fengycin) 、表面活性素 (surfactin) 和依枯草素 (iturins) ,其中泛革素對多種植物病原菌具有較強的抑制活性,在植物病害防治中發揮著重要作用。Islam 等[6]鑒定出枯草芽孢桿菌C9 菌株產生的主要抗真菌物質為泛革素,并通過單因素試驗發現以甘露醇為碳源、以黃豆粉為氮源時泛革素的產量最高。Yaseen等[7]則發現,不同碳源和氮源會影響泛革素啟動子的表達,以尿素或尿素和銨鹽的混合物為氮源、以甘露醇為碳源時,可以提高泛革素啟動子的活性,從而提高其產量。
目前工業化的細菌培養基原料主要包括糖類、玉米粉、淀粉、各種油料作物餅粉和魚粉等農副產品,以及銨鹽、硝酸鹽和鉀、鈉、鎂等礦物質元素。其中葡萄糖、糖蜜、淀粉及玉米粉主要提供碳源,豆餅粉、魚粉、銨鹽和硝酸鹽主要提供氮源。鐘蔚[8]通過單因素篩選試驗發現,以麥芽浸粉作為碳源時,枯草芽孢桿菌BS1 菌株芽孢濃度最高;以黃豆粉作為氮源時,雖然芽孢濃度較低,但由于菌體濃度大,所以其芽孢產量仍遠高于其他氮源條件下的芽孢產量,因此仍可作為最適氮源用于BS1 菌株的發酵。也有研究報道,紅糖、淀粉和葡萄糖可作為枯草芽孢桿菌的最佳碳源,而花生餅粉、豆粕、酵母浸粉及牛肉膏可作為枯草芽孢桿菌的最佳氮源[9-12]。由此可見,不同菌株適宜的最佳碳源和氮源各不相同。
枯草芽孢桿菌HMB19198 菌株是河北省農林科學院植物保護研究所植物病害生物防治實驗室分離獲得的一株生防細菌,已證明以其為活性成分的微生物菌劑能有效防治番茄灰霉病和瓜類白粉病,且該菌株可產生脂肽類抗生素泛革素和表面活性素等活性物質,其中泛革素是主要的抑菌活性物質[13]。為提高HMB19198 菌株的發酵水平,降低相關菌劑的生產成本,本研究以活芽孢濃度為指標,首先從工業生產常用培養基中篩選出較適合HMB19198 菌株芽孢形成的基礎配方,并進一步對碳源和氮源進行篩選及優化,結合Plackett-Bruman 設計與響應曲面分析 (RSA) ,獲得到了利于HMB19198 菌株芽孢形成的發酵培養基。
1 材料與方法
1.1 供試菌株及培養條件
枯草芽孢桿菌Bacillus subtilisHMB19198 菌株,由河北省農林科學院植物保護研究所分離和保存,中國微生物菌種保藏中心保藏編號為CGMCC No. 5613,于?80 ℃長期保存在含有體積分數30%甘油的LB 培養基 (胰蛋白胨10 g/L、酵母粉5 g/L、NaCl 5 g/L) 中,使用前在LB 固體培養基上于37 ℃進行活化。
1.2 芽孢濃度測定
取1 mL 菌液,于80 ℃高溫處理10 min,之后用滅菌水系列稀釋,分別取100 μL 10?4和10?5稀釋倍數的菌懸液,涂布于LB 固體培養基上,于30 ℃培養過夜,統計芽孢濃度。以未經高溫處理的菌懸液為空白對照,并按式 (1) 計算芽孢的形成率[14]。
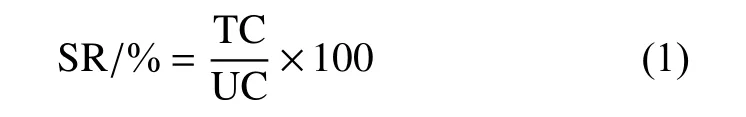
式中:SR 為芽孢形成率,%;TC 為80 ℃高溫處理組菌落數,CFU/mL;UC 為空白對照組菌落數,CFU/mL。
1.3 基礎培養基篩選
根據文獻報道[7,15-18],選擇8 種常用于芽孢桿菌的培養基 (標號1 ~ 8) 作為HMB19198 菌株的基礎培養基 (表1) 。將活化好的HMB19198 菌株接種于5 mL LB 培養基中,37 ℃、160 r/min 振蕩培養12 h后,按照1 : 100 的比例接種于100 mL 各基礎培養基中,37 ℃、160 r/min 振蕩培養48 h,篩選發酵液中芽孢濃度最高的培養基作為HMB19198 菌株的基礎培養基。基礎培養基配方信息詳見表1。
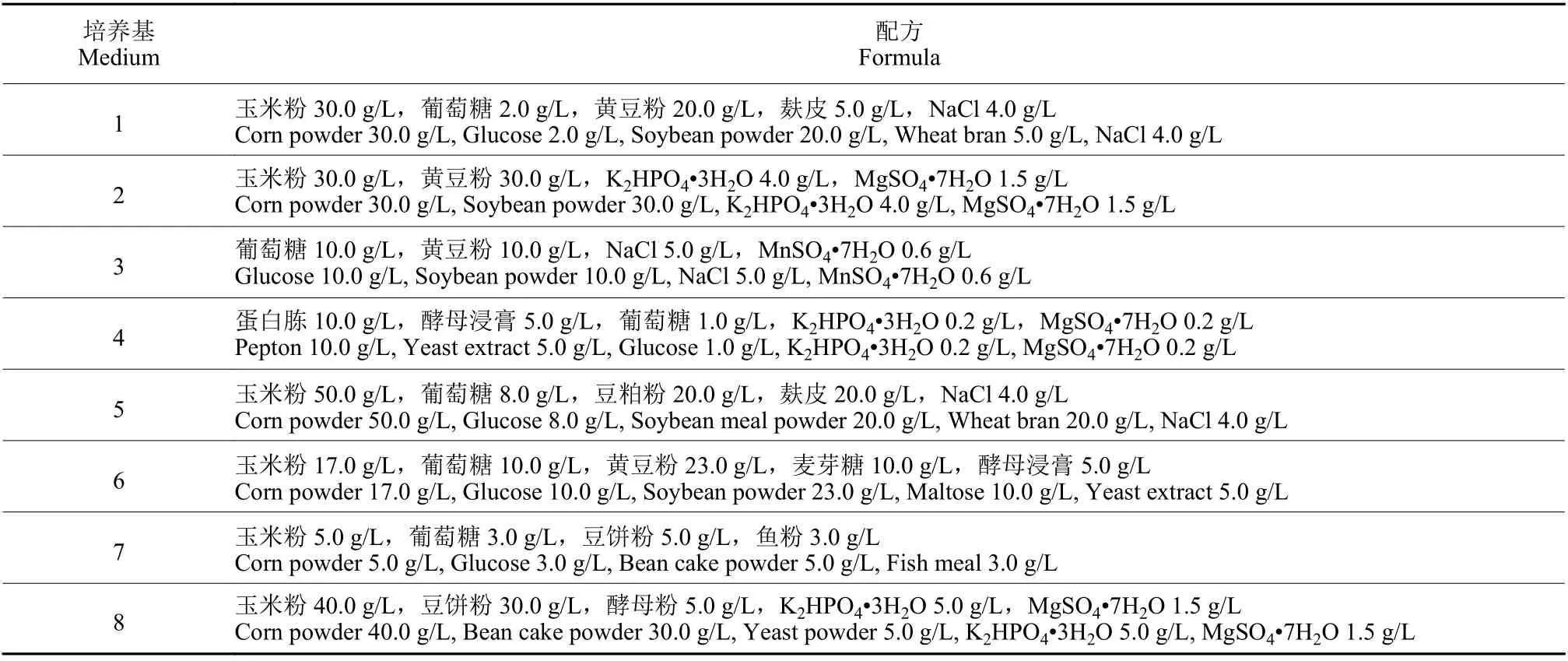
表1 芽孢桿菌基礎培養基Table 1 Basic mediums for Bacillus
1.4 最適碳源及氮源篩選
在篩選出利于HMB19198 菌株芽孢形成的基礎培養基基礎上,采用單因素試驗方法,選取12種碳源 (麥麩粉、水解玉米粉、玉米粉、糖蜜、葡萄糖、麥芽糖、蔗糖、阿拉伯糖、果糖、甘露糖、可溶性玉米淀粉和蘋果酸) 及10 種氮源 (豆餅浸粉、酶解玉米漿、花生餅浸粉、棉籽餅粉、蛋白胨、酵母粉、牛肉浸膏、魚粉、黃豆粉和硫酸銨) ,以30 g/L質量濃度分別替換基礎培養基中的碳源和氮源。按照1.2 節所述方法測定發酵液中芽孢濃度,篩選利于HMB19198 菌株芽孢形成的最佳碳源和氮源。
1.5 Plackett-Burman 法分析培養基中影響菌株芽孢形成的主要因子
Plackett-Burman 法是一種近飽和的兩水平試驗設計方法,該方法能通過最少試驗次數快速篩選出影響某一性狀的主要因子[19]。本研究利用Plackett-Burman 法對利于HMB19198 菌株芽孢形成的碳、氮源進行優化,每個因素分別取低水平“?1”和高水平“1”兩個水平,采用 Minitab 軟件進行方差分析。具體試驗因子及水平見表2。
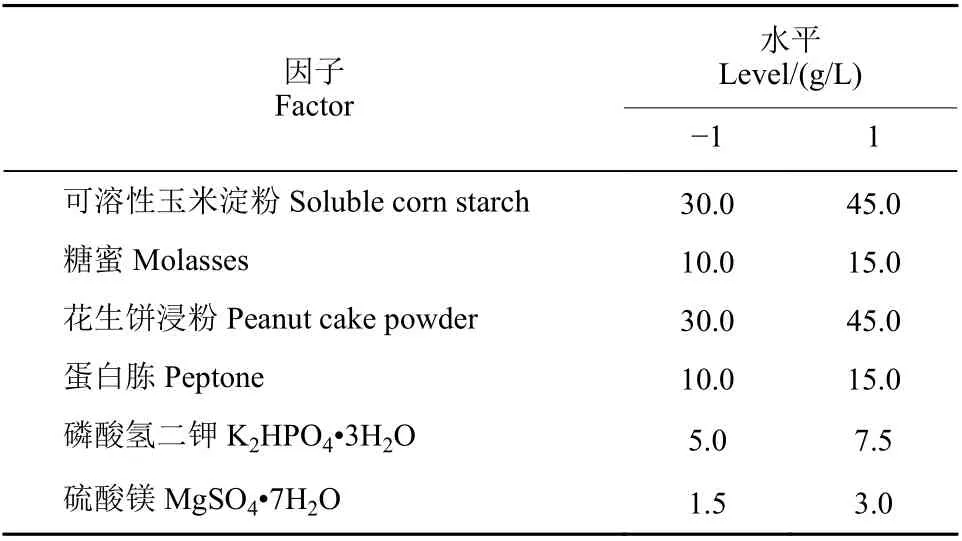
表2 Plackett-Burman 試驗因子與水平設計Table 2 Experimental design of the factors and levels of Plackett-Burman
1.6 響應曲面分析法確定發酵液芽孢濃度最高時的培養基配方
響應曲面分析法是一種尋找多因素系統中最佳條件的數學統計方法[19]。在Plackett-Burman 分析的基礎上,以可溶性玉米淀粉、糖蜜、花生餅浸粉、K2HPO4?3H2O 為參考因子,以發酵液中芽孢濃度為指標,采用Design-Expert 軟件按照Boxbehnken 中心組合設計4 因子、3 水平、29 次運行的響應曲面試驗。具體試驗因子及水平見表3。通過對結果數據進行二次回歸擬合,最后在一定的水平范圍內求出最佳值。
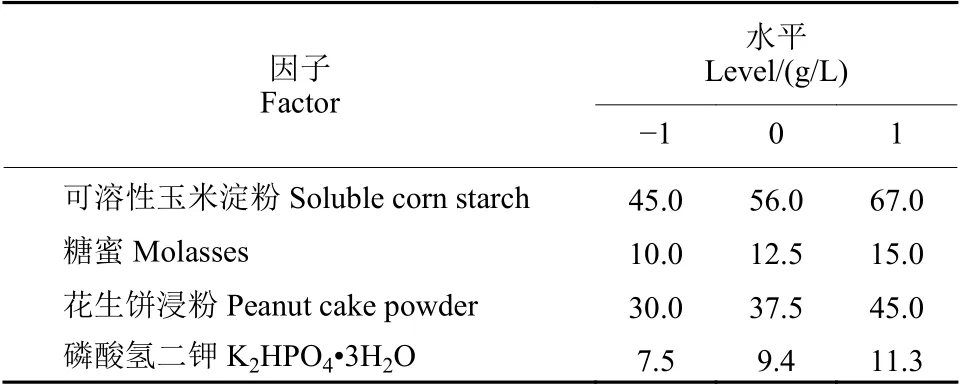
表3 響應曲面試驗因子與水平設計Table 3 Experimental design of the factors and levels of RSA
1.7 枯草芽孢桿菌HMB19198 菌株生長曲線測定
按照優化后的配方配制優化培養基,以基礎培養基為對照進行搖瓶培養。將過夜培養的HMB19198 菌株按照1 : 100 的比例分別接種到優化培養基和基礎培養基中,于37 ℃、160 r/min 下振蕩培養,每隔2 h 取樣統計發酵液中芽孢濃度,計算芽孢形成率,并繪制生長曲線。
1.8 優化培養基中發酵液所產泛革素濃度測定
將HMB19198 菌株分別以優化培養基和基礎培養基于37 ℃、160 r/min 下振蕩培養48 h,獲得菌株發酵液;發酵液于4 ℃、10000 r/min 下離心20 min,收集上清液;用6 mol/L 鹽酸溶液調節上清液pH 值至2.0,4 ℃下過夜,次日于10000 r/min下離心20 min,收集沉淀;沉淀干燥后加入甲醇抽提2 次,合并抽提液得到脂肽粗提物。
采用高效液相色譜 (HPLC) 梯度洗脫分離脂肽化合物。HPLC 分析系統為 AKTA Purifier 10 液相分析系統,色譜柱為 SOURCE5 RPC ST4.6/150柱。流動相A 為V(三氟乙酸) :V(乙腈) :V(水) =0.065 : 2﹕97.935,流動相B 為V(三氟乙酸) :V(乙腈) :V(水) = 0.05 : 80 : 19.95。梯度洗脫,49.86 min內流動相A 由100%到0。檢測波長215 nm,流速1 mL/min。本團隊前期研究已明確,保留時間在20~25 min 的物質為泛革素[13],因此,本研究將20~25 min 期間洗脫液的峰面積除以對應處理的芽孢濃度作為不同培養基發酵液中泛革素的濃度。
2 結果與討論
2.1 最適基礎培養基
利用8 種芽孢桿菌常用培養基對HMB19198菌株進行振蕩培養,測定不同培養基中HMB19198菌株的芽孢濃度。結果表明:2 號培養基中發酵液的芽孢濃度最高,達到3.34 × 109個/mL;其次是5 號培養基,芽孢濃度為2.97 × 109個/mL,1 號和6 號培養基結果與5 號培養基接近,分別為2.90 ×109和2.83 × 109個/mL;4 號培養基中發酵液的芽孢濃度最低,僅為9.00 × 108個/mL (圖1)。由于2 號培養基中的芽孢濃度最高,并且其組分簡單、原料來源豐富、價格低廉且取材方便,因此,確定選擇2 號培養基作為基礎培養基進行進一步優化。
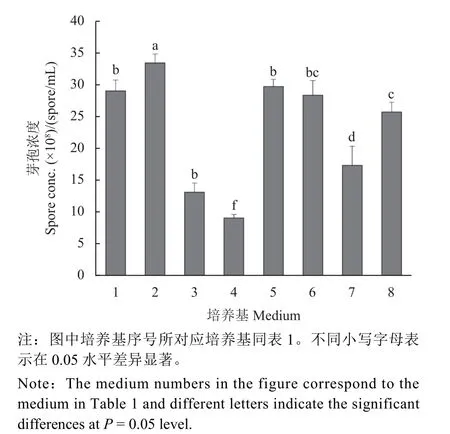
圖1 不同基礎培養基對枯草芽孢桿菌HMB19198菌株芽孢濃度的影響Fig. 1 Effect of different basic mediums on the spore concentration of B. subtilis strain HMB19198
2.2 最適宜芽孢形成的碳源及氮源確定
比較了12 種碳源對HMB19198 菌株芽孢濃度的影響。結果 (圖2A) 表明:在以復雜碳源為唯一碳源的培養基中,HMB19198 菌株發酵液的芽孢濃度較高,其中以可溶性玉米淀粉作為唯一碳源時芽孢濃度最高,達到4.52 × 109個/mL;其次是以糖蜜為唯一碳源的培養基,芽孢濃度為3.38 ×109個/mL;以麥麩粉、玉米粉和水解玉米粉為唯一碳源時,芽孢濃度分別為2.14 × 109、2.05 × 109和1.93 × 109個/mL。而在以糖類物質為唯一碳源的培養基中,HMB19198 菌株發酵液的芽孢濃度較低。由于芽孢桿菌攝取糖蜜的速度較快,可作為發酵前期的速效碳源,攝取玉米淀粉的速度較慢,將其作為發酵中后期的遲效碳源,兩者結合構成最適碳源組合。
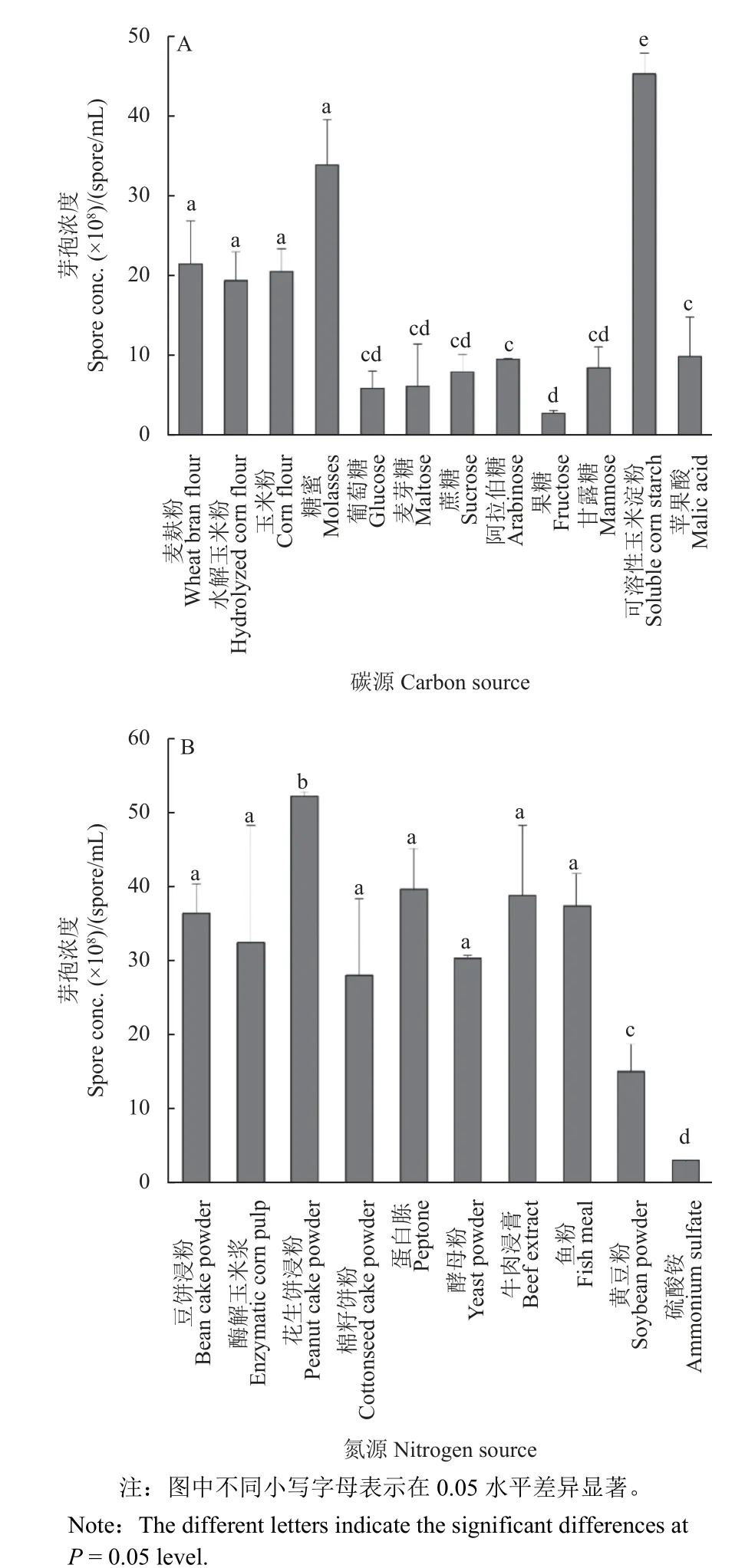
圖2 不同碳源 (A) 和氮源 (B) 對枯草芽孢桿菌HMB19198 菌株芽孢形成的影響Fig. 2 Spore concentrations of B. subtilis strain HMB19198 in basic medium supplemented with different carbon (A) and nitrogen (B) sources
在確定了以可溶性玉米淀粉和糖蜜為最適碳源的基礎上,進一步針對10 種氮源物質進行了優化。結果 (圖2B) 表明:在以花生餅浸粉為氮源的培養基中,HMB19198 菌株發酵液的芽孢濃度最高,為5.22 × 109個/mL;其次是蛋白胨和牛肉浸膏,芽孢濃度分別為3.03 × 109和3.88 × 109個/mL;而以無機氮源 (NH4)2SO4為氮源的培養基中芽孢濃度最低,僅為2.90 × 108個/mL。雖然以牛肉浸膏和蛋白胨作為氮源時芽孢濃度相近,但牛肉浸膏成本較高,故選取芽孢桿菌攝取速度較快的蛋白胨作為發酵前期的速效氮源,以花生餅浸粉作為發酵中后期的遲效氮源,兩者結合構成最適氮源組合。
2.3 基礎培養基中影響芽孢形成的主要因子
分別以可溶性玉米淀粉和糖蜜為碳源,以花生餅浸粉和蛋白胨為氮源,無機鹽為K2HPO4?3H2O和MgSO4?7H2O,利用Plackett-Burman 法對6 個因素 (表4) 進行優化。結果 (表5) 表明:可溶性玉米淀粉、花生餅浸粉、糖蜜、K2HPO4?3H2O對HMB19198 菌株芽孢形成影響顯著,是影響發酵液中芽孢濃度的主要因子。圖4 中各因子的模型預測結果顯示,可溶性玉米淀粉和K2HPO4?3H2O濃度與芽孢濃度呈正相關,糖蜜和花生餅浸粉濃度與芽孢濃度呈負相關。
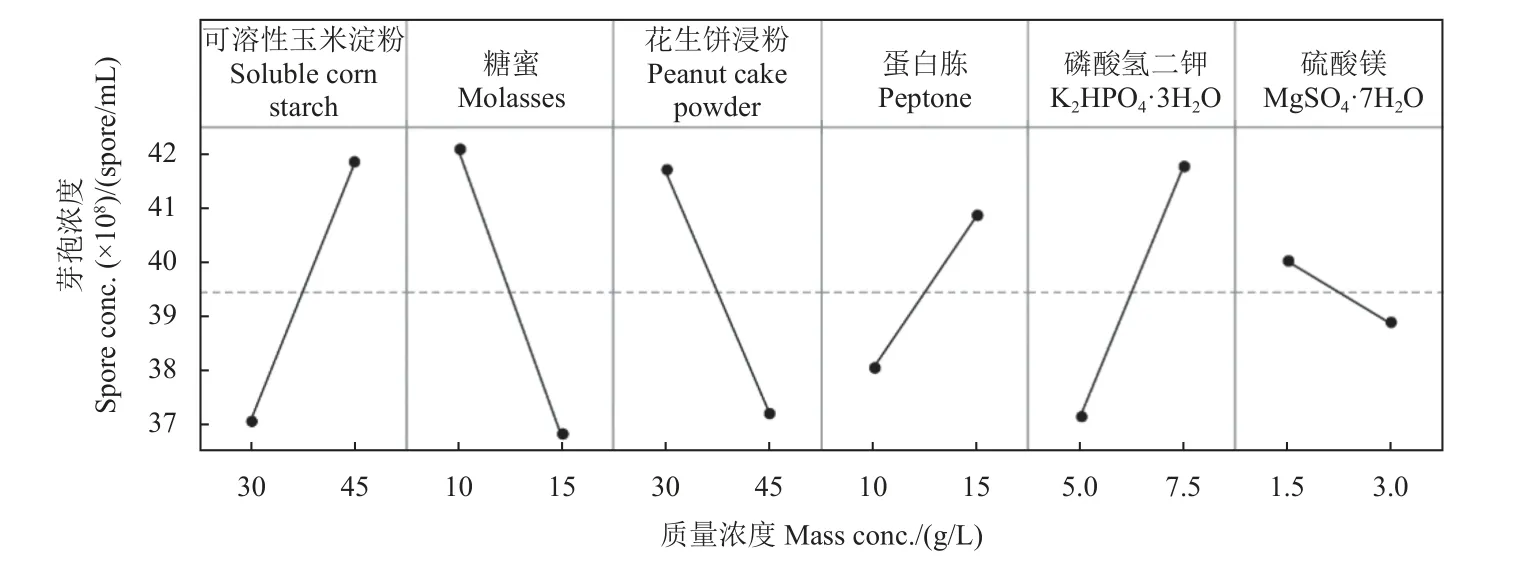
圖4 影響芽孢形成主要因子的預測模型Fig. 4 Predictive models of the main factors affecting spore formation
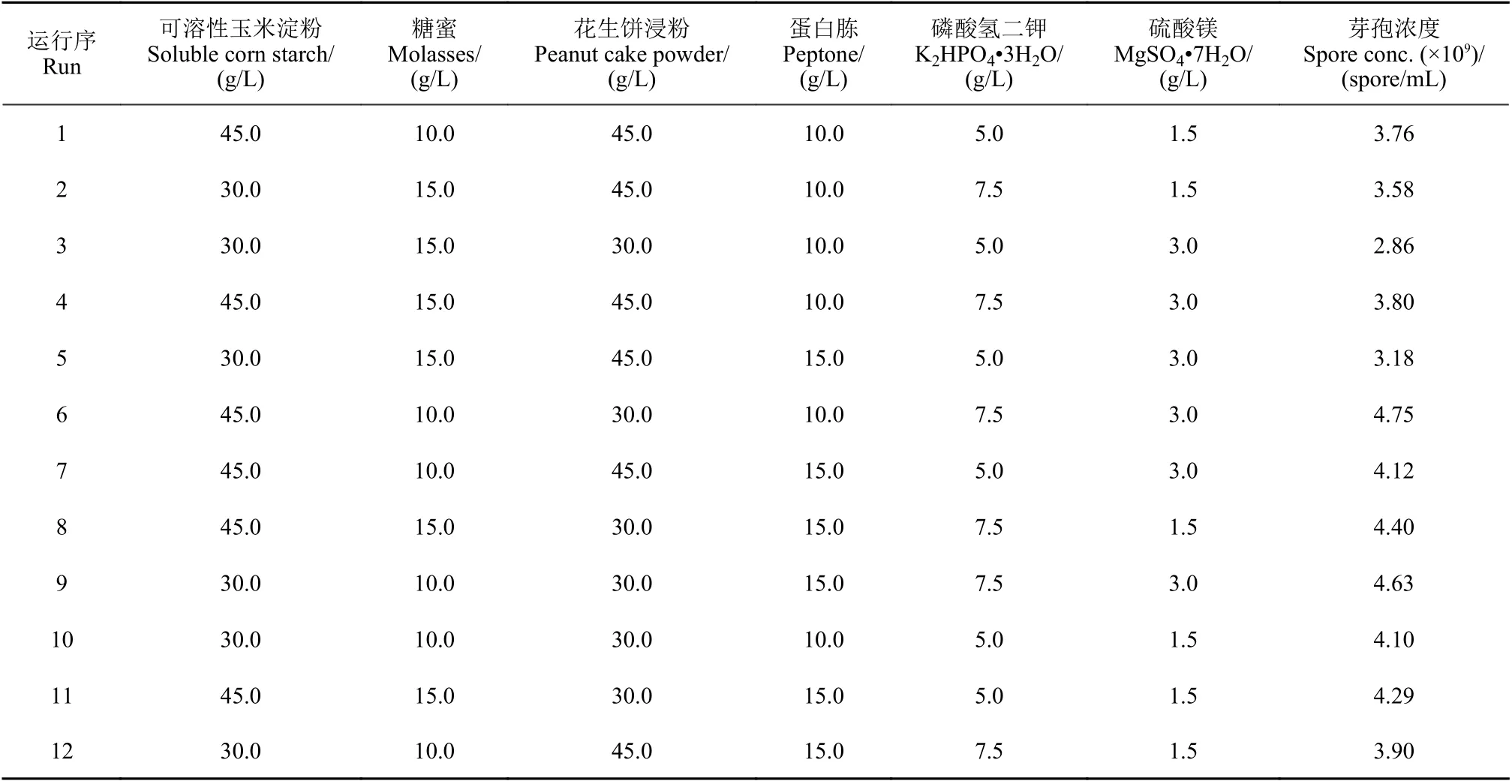
表4 Plackett-Burman 試驗設計與芽孢濃度響應Table 4 Experimental designs of Plackett-Burman and corresponding spore concentrations
2.4 培養基中主要影響因子的最佳濃度及驗證
以Plackett-Burman 分析所得4 種主要影響因子為基礎,以可溶性玉米淀粉45 g/L、糖蜜10 g/L、花生餅浸粉30 g/L 及K2HPO4?3H2O 7.5 g/L 作為中心點,進行響應曲面分析,試驗設計及結果見表5。
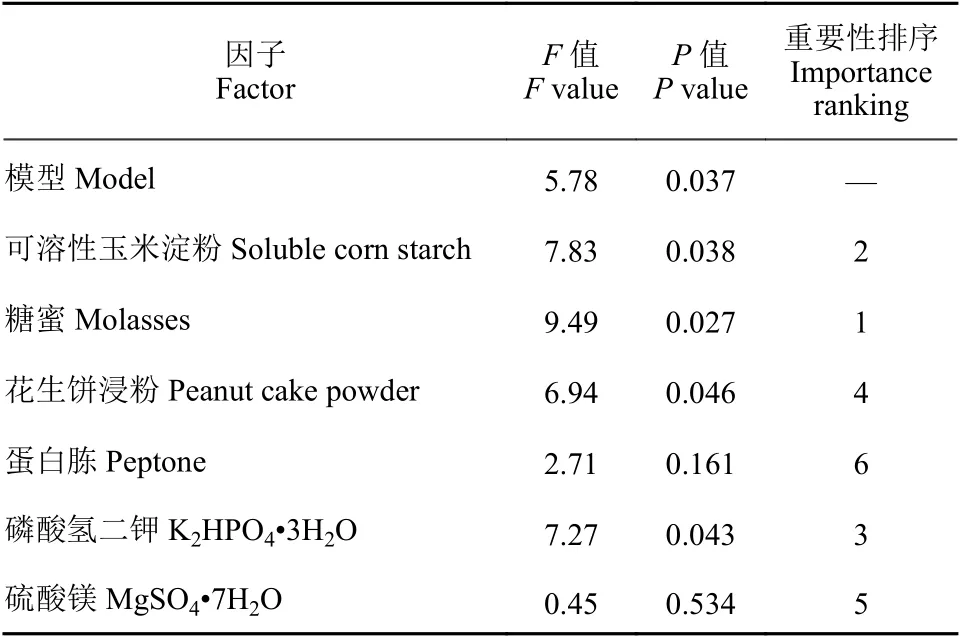
表5 Plackett-Burman 試驗統計分析Table 5 The regression analysises of Plackett-Burman experiments
利用Design Expert 10 軟件設計響應曲面試驗(表6),對數據分析并繪制響應曲面圖和等高線圖 (圖5) ,回歸分析結果見表7。統計結果顯示,A (可溶性玉米淀粉)、A × C (可溶性玉米淀粉和花生餅浸粉交互) 和B × C (糖蜜和花生餅浸粉交互)對發酵液中芽孢濃度影響影響顯著。
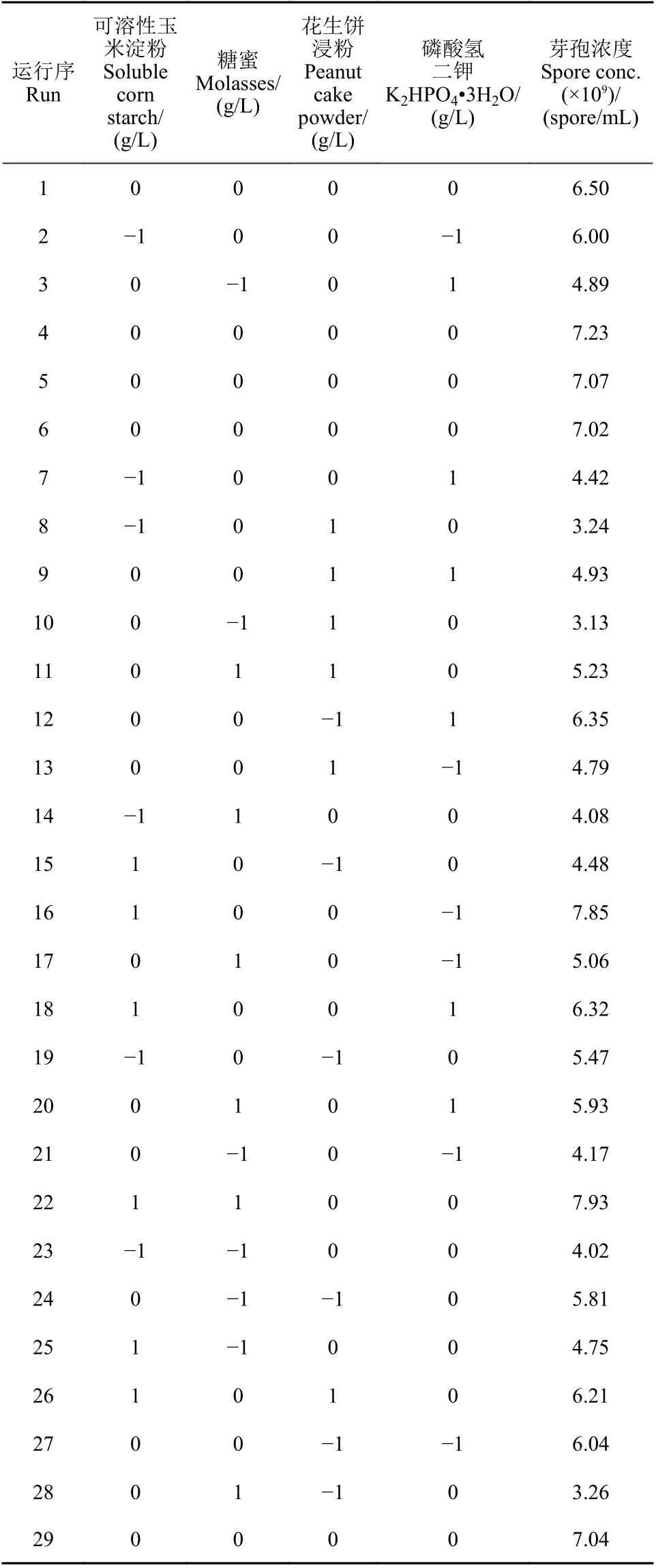
表6 響應曲面分析試驗設計及結果Table 6 Experimental designs and results of RSA
當確定糖蜜14.1 g/L、K2HPO4?3H2O 9.2 g/L為最佳值時,分析了可溶性玉米淀粉和花生餅浸粉的最佳比例。由圖5a 可見:在可溶性玉米淀粉為低水平時,隨著花生餅浸粉質量濃度升高,發酵液中芽孢濃度逐漸降低;在可溶性玉米淀粉為高水平時,隨著花生餅浸粉質量濃度升高,芽孢濃度呈先升高后降低趨勢。在花生餅浸粉為低水平時,隨著可溶性玉米淀粉濃度升高,發酵液中芽孢濃度變化較小;在花生餅浸粉為高水平時,隨著可溶性玉米淀粉濃度升高,芽孢濃度也逐漸升高。據此,確定兩者最佳配比為可溶性玉米淀粉67 g/L,花生餅浸粉41.6 g/L。圖5b 顯示,可溶性玉米淀粉與花生餅浸粉兩個因素的交互作用顯著。
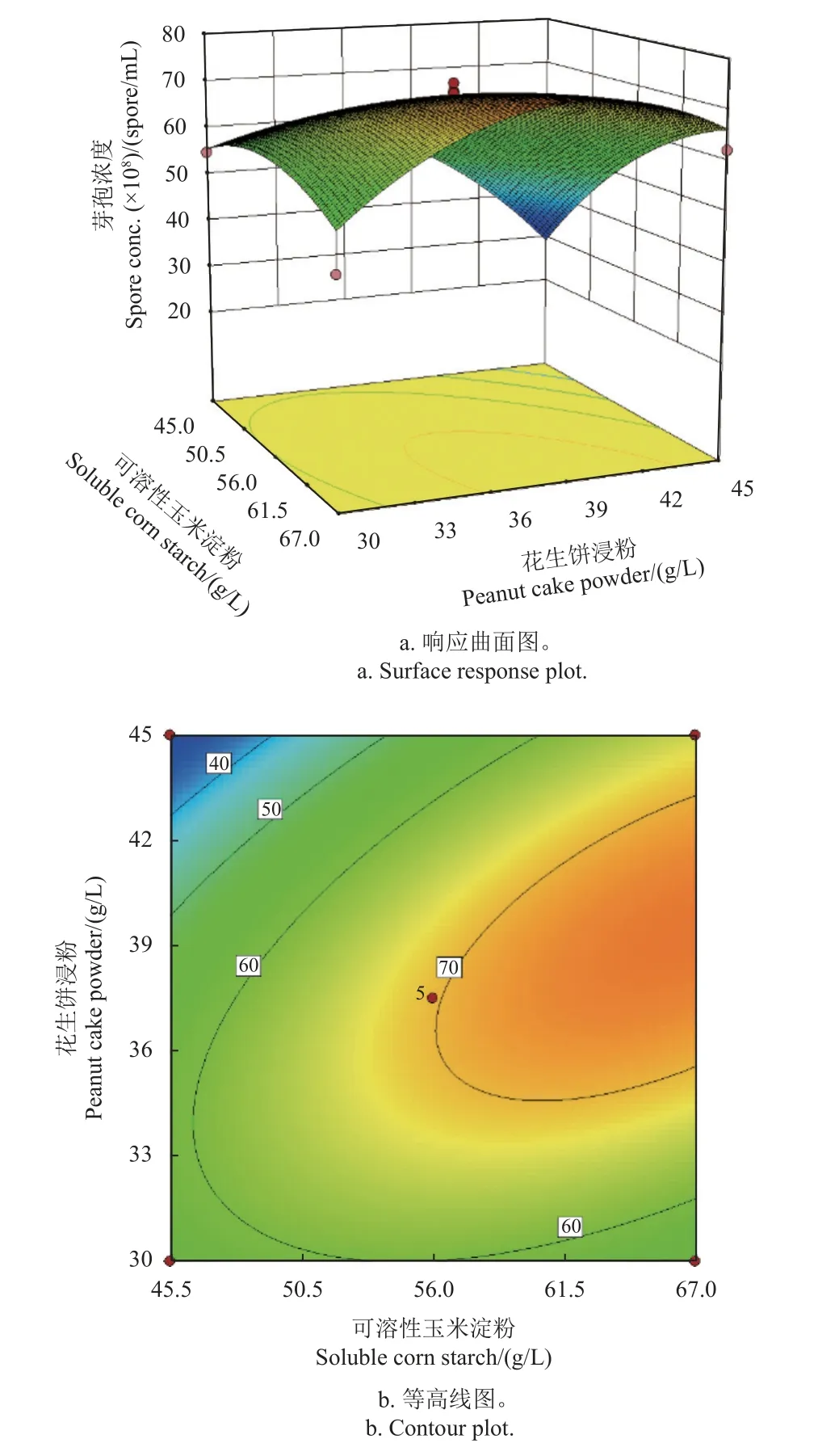
圖5 可溶性玉米淀粉與花生餅浸粉雙因素最適濃度及配比模擬結果Fig. 5 Simulation results of the optimum concentrations and ratios of soluble corn starch and peanut cake powder
當確定可溶性玉米淀粉67 g/L、K2HPO4?3H2O 9.2 g/L 為最佳值時,分析了糖蜜和花生餅浸粉的最佳比例。由圖6a 可見:在糖蜜為低水平時,隨著花生餅浸粉質量濃度升高,發酵液中芽孢濃度逐漸降低;在糖蜜為高水平時,隨著花生餅浸粉質量濃度升高,發酵液中芽孢濃度呈先升高后降低趨勢。而無論花生餅浸粉為高水平或低水平,隨著糖蜜濃度升高,發酵液中芽孢濃度均呈先升高后降低趨勢。綜上,確定兩者的最佳配比為糖蜜14.1 g/L,花生餅浸粉41.6 g/L。圖6b 顯示,糖蜜和花生餅浸粉兩個因素的交互作用顯著。
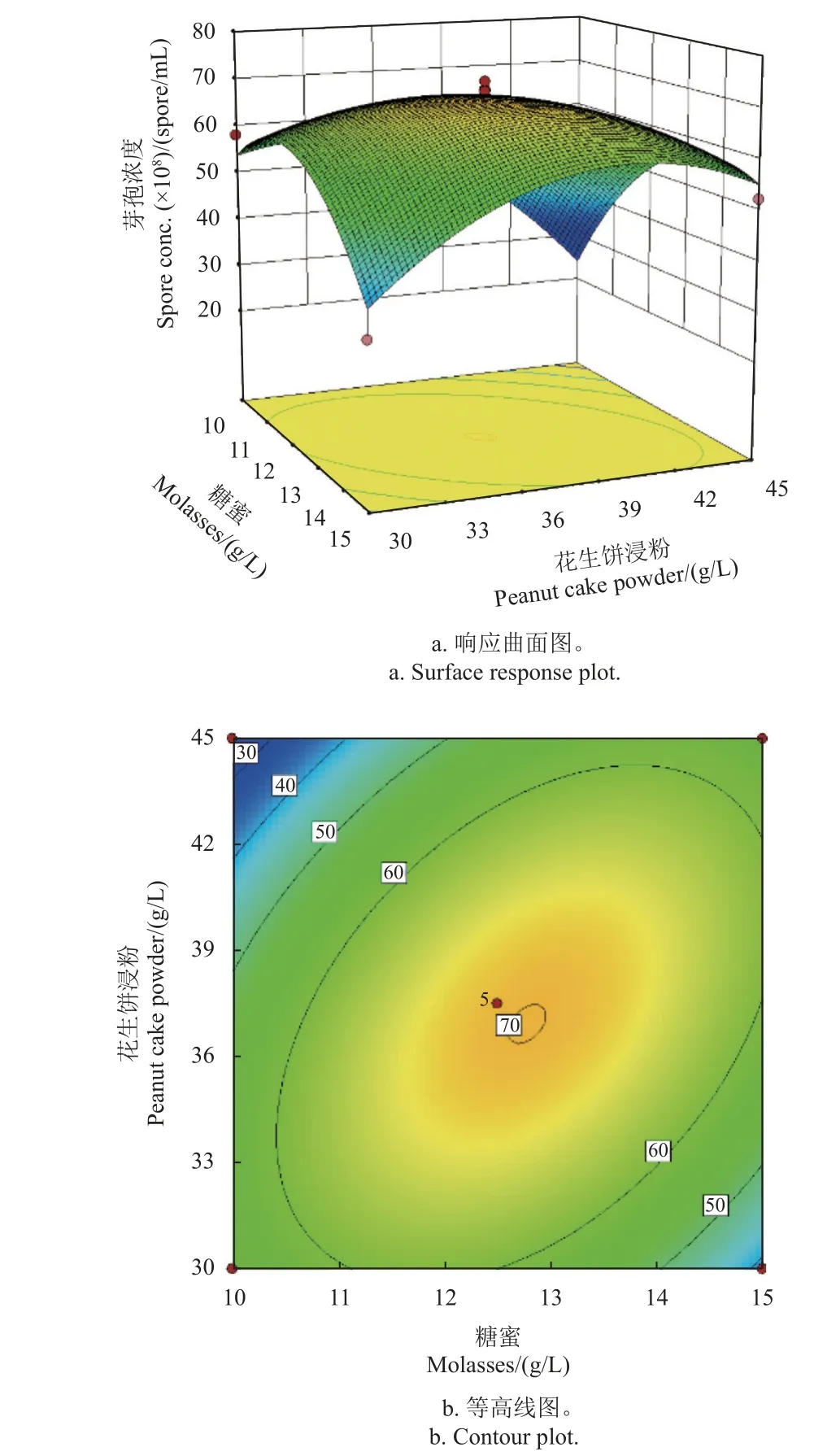
圖6 糖蜜與花生餅浸粉雙因素最適濃度及配比模擬結果Fig. 6 Simulation results of the optimum concentrations and ratios of molasses and peanut cake powder
圖5 和圖6 的響應面均為開口向下的凸型曲面,代表響應值存在極高值。利用Design Expert 10 軟件對表7 進行進一步分析計算,得到模型預測的最佳點:即當可溶性玉米淀粉質量濃度為67.0 g/L、糖蜜為14.1 g/L、花生餅浸粉為41.6 g/L、K2HPO4?3H2O 為 9.2 g/L 時,預測HMB19198 菌株發酵液中芽孢濃度最高,為7.83 × 109個/mL。
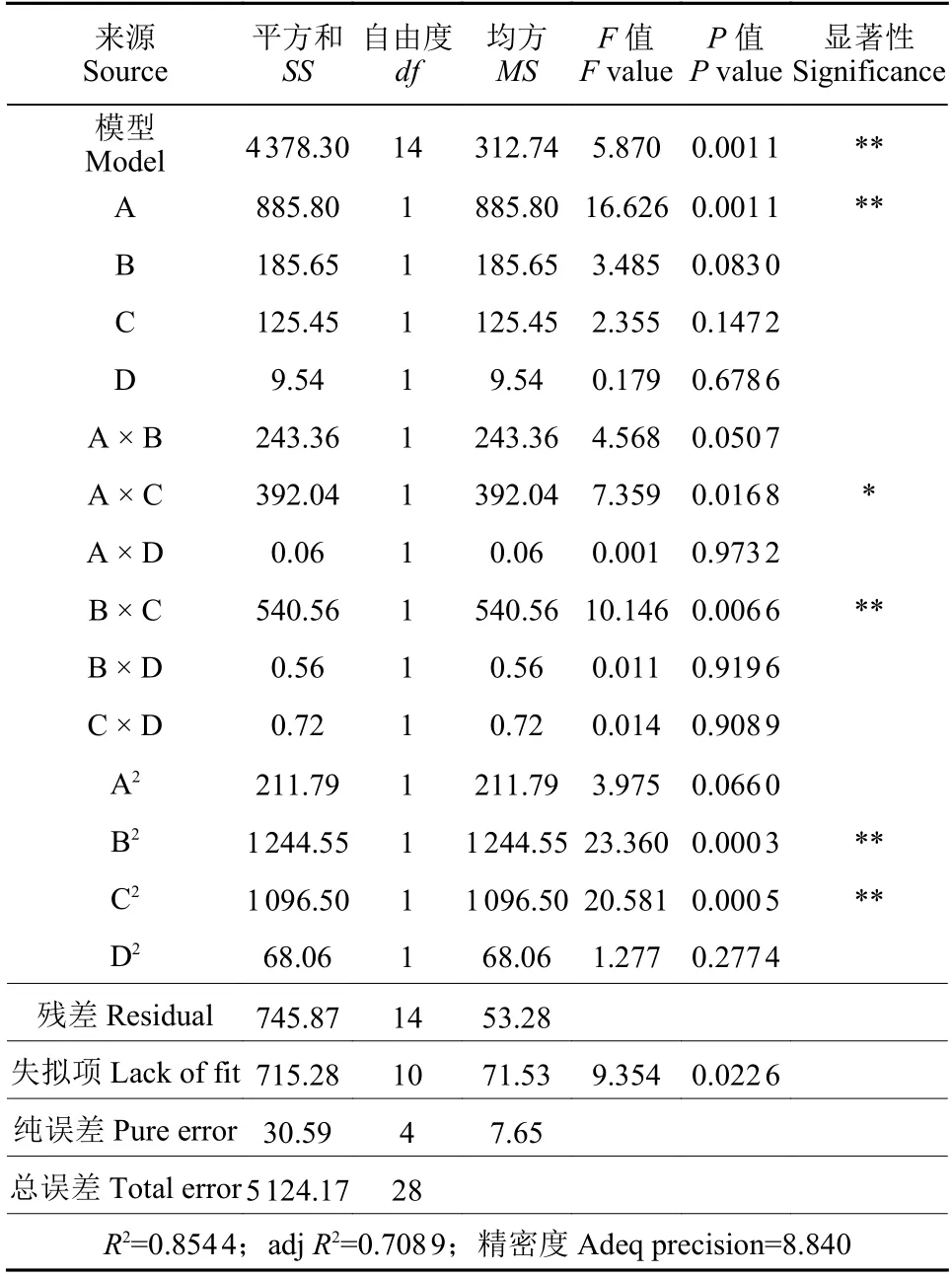
表7 響應曲面試驗回歸分析結果Table 7 The regression analysis of RSA
2.5 模型驗證
按照響應曲面模型預測所得最佳組合,即可溶性玉米淀粉67.0 g/L、糖蜜14.1 g/L、花生餅浸粉41.6 g/L、蛋白胨10 g/L、K2HPO4?3H2O 9.2 g/L及MgSO4?7H2O 1.5 g/L,配制得到優化培養基,將HMB19198 菌株在優化后的培養基中振蕩培養并統計芽孢濃度。結果表明,培養24 h 后,HMB19198菌株發酵液中芽孢濃度為6.92 × 109個/mL,與預測值7.83 × 109個/mL 比較接近,表明模型預測結果準確。
2.6 菌株生長曲線
HMB19198 菌株在優化培養基中培養6 h 后進入對數生長期,16 h 開始生長趨緩,20 h 時芽孢濃度達到最大值1.10 × 1010個/mL;該菌株在基礎培養基中培養12 h 進入對數生長期,26 h 時芽孢濃度達到最大值7.08 × 109個/mL (圖7)。研究表明,與基礎培養基相比,在優化后培養基中,HMB19198菌株發酵周期可縮短40%,芽孢濃度提高了54.8%。
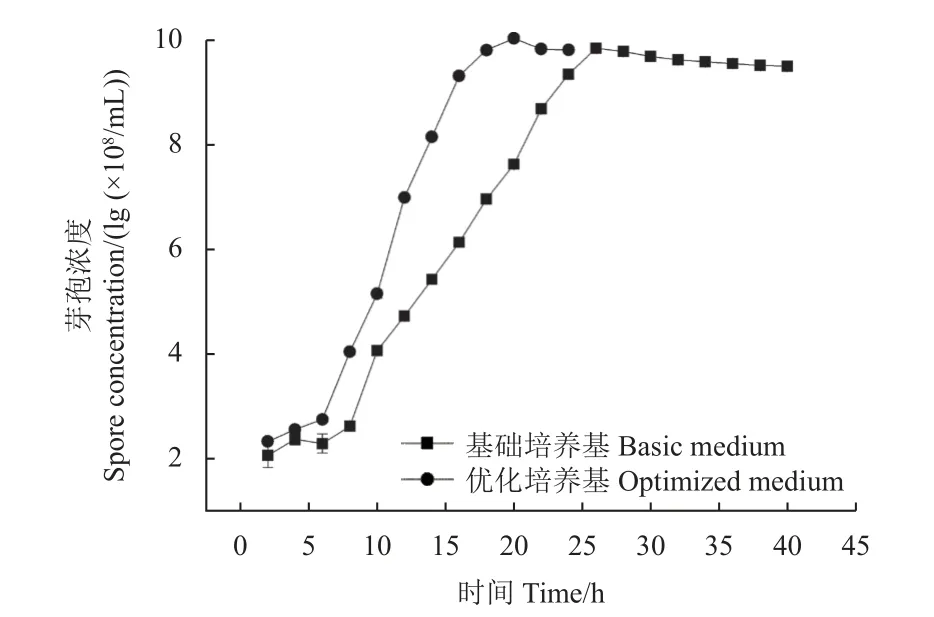
圖7 枯草芽孢桿菌HMB19198 菌株的生長曲線Fig. 7 Growth curve of B. subtilis strain HMB19198
2.7 優化培養基中發酵液產生的泛革素濃度
HMB19198 菌株分別在優化培養基和基礎培養基中振蕩培養后,提取2 種培養基中的脂肽類抗生素粗提液,利用HPLC 進行分析。結果表明,與基礎培養基相比,優化后培養基中泛革素的產量提高了39.4% (表8) 。

表8 培養基優化前后發酵液中泛革素濃度Table 8 Relative contents of fengycin in fermentation mediums before and after optimization
3 結論與討論
芽孢是芽孢桿菌產生的休眠體,含水量極低,抗逆性強,能耐受高溫、紫外線等。以芽孢為主要成分的微生物農藥制劑貨架期較長,是開發微生物源殺菌劑的主要成分[20]。
碳源物質是培養基中的重要組分之一,負責為細胞提供能源、構成碳架及合成代謝產物[21]。常見碳源物質主要包括碳水化合物、脂肪、有機酸等。以生產芽孢為目的的芽孢桿菌發酵培養基通常以廉價且易獲取的農副產品及各種糖類為碳源,如玉米粉、淀粉、糖蜜、麩皮及蔗糖等[22-23]。本研究通過對不同碳源物質進行優化,發現以可溶性玉米淀粉為碳源能提高HMB19198 菌株發酵液的芽孢濃度。可溶性玉米淀粉是植物多糖類碳源,在細菌發酵過程中可被降解為葡萄糖,進而被細胞吸收和轉化。葡萄糖是細菌優先利用的碳源,但過量的葡萄糖也會抑制細菌的生長和代謝代謝[24-25]。由于淀粉降解為葡萄糖存在一個緩慢的過程,保證了培養基中葡萄糖不會大量積累,因而避免了過量葡萄糖所造成的負面效應[26]。尹鳳嬌等[27]通過單因素試驗,篩選出了最適合解淀粉芽孢桿菌B. amyloliquefaciensY4 菌株芽孢形成的碳源為可溶性淀粉,并通過Plackett-Burman 分析,明確了可溶性淀粉是影響發酵液中芽孢濃度的主要因子。糖蜜是食糖加工過程的副產品,含有較高濃度的蔗糖,易被細胞吸收。本研究發現,糖蜜也能提高HMB19198 菌株發酵液的芽孢濃度,但糖蜜的濃度與芽孢形成率呈負相關,說明高濃度的糖蜜會抑制HMB19198 菌株芽孢的形成,因此,糖蜜可作為芽孢桿菌發酵前期的能量來源。
氮源物質的主要功能是構成細胞結構 (氨基酸、蛋白質、核酸等) 及合成含氮代謝產物。根據被微生物利用的速度,氮源物質可分為遲效氮源(黃豆粉、花生餅浸粉、棉籽餅粉等) 和速效氮源(蛋白胨、硫酸銨等) 。遲效氮源通常為蛋白質含量較高的農副產品,其中花生餅浸粉含氮量為8.0%,高于豆餅粉 (6.71%) 、棉籽餅粉 (6.56%) 和黃豆粉 (5.2%)[28]。本研究發現,以花生餅浸粉為氮源能顯著提高HMB19198 菌株的芽孢形成率。原因可能除花生餅浸粉蛋白質含量較高外,還因其含有較多的氨基酸、多肽及某些生長因子,能促進HMB19198 菌株的生長和代謝。
碳/氮比 (C/N ratio) 是微生物發酵培養基的重要參數,直接影響發酵過程中菌體的生長和代謝,碳/氮比過高或過低均不利于芽孢桿菌的發酵[29]。適合芽孢桿菌發酵的培養基碳/氮比通常為5~10;碳/氮比偏高 (12~20) 通常會導致芽孢產量降低,但可刺激代謝產物的分泌;而碳/氮比過高(> 20) 會導致芽孢及代謝產物產量同時降低;碳/氮比過低 (< 5) 則會導致菌體自溶[30]。本研究通過響應曲面法,分析了可溶性玉米淀粉和糖蜜組成的碳源與不同比例花生餅浸粉氮源對HMB19198菌株芽孢形成的影響。結果發現,可溶性玉米淀粉和糖蜜濃度處于較高水平、花生餅浸粉濃度在中等水平,即碳/氮比為15.8 時,發酵液中芽孢濃度最高。本研究優化后培養基的碳/氮比略高于上述碳/氮比最適區間 (5~10) ,其原因可能是由于芽胞桿菌發酵的最適碳/氮比還受其他因素影響。不同碳源與氮源組合的最適碳/氮比通常不同[31],另外,不同菌種對碳/氮比的偏好也不同。相較于芽孢桿菌,假單胞菌的碳/氮比最適區間為7~14;酵母菌在碳/氮比為4 時菌體生長旺盛,而在碳/氮比為3 時,可代謝產生較多的谷氨酸[31]。Jin 等[15]發現,用菜籽粕代替蛋白胨作為氮源,在相同碳源情況下可使枯草芽孢桿菌 3-10 菌株的發酵水平提高20%,并使脂肽類抗生素伊枯草菌素 (iturins)的產量提高了8 倍。除此之外,碳/氮比對芽孢產量的影響也會隨氧傳遞系數的增大而逐漸降低。培養基中固體含量較多或質地較粘稠都會導致氧傳遞系數降低,通氣性變差,此時就需要較高的碳/氮比以維持細菌旺盛的生長[32]。
培養基中的礦物質元素雖然不參與細胞物質的組成,但卻是很多酶的輔助因子。鉀離子與細胞滲透壓和透性相關,在培養基中可起到一定的緩沖劑作用并維持pH 值穩定。本研究發現,K2HPO4?3H2O 濃度與HMB19198 菌株的芽孢濃度呈正相關,且9.2 g/L 的K2HPO4?3H2O 最適合HMB19198 菌株芽孢的形成。不同菌株對鉀離子濃度的需求可能不同。林陳強等[33]的研究表明,培養基中添加適量的鉀離子能促進地衣芽孢桿菌CHB6 菌株的生長和芽孢形成,優化后的培養基中芽孢濃度可達到4.8 × 109個/mL,芽孢形成率為90.6%。張麗霞[34]通過分析K、Mg、Mn、Fe、Zn、Cu、Mo 及Ca 等礦物質養料對芽孢桿菌生長的影響,確定了K 對枯草芽孢桿菌發酵液芽孢濃度影響最大,進一步明確了K2HPO4是最佳的K供體,在K 質量濃度為0.7 g/L 時,發酵液中芽孢濃度最高,較優化前提高了29%。K2HPO4在提供K 的同時還可以提供P 元素,P 是核酸和蛋白質的必要成分,也是細胞膜的重要組分[21],同時P 還是微生物平衡生長的限制因素。
泛革素是枯草芽孢桿菌及其近源種產生的一種脂肽類抗生素,在抑制病原菌生長和防治作物病害中發揮著重要作用[35-36]。本團隊前期研究[13]證明,泛革素是HMB19198 菌株產生的主要抑菌活性物質,在抑制番茄灰霉病菌生長和防治番茄灰霉病中具有重要作用。因此,提高發酵液中泛革素的含量能提高HMB19198 菌株的防病效果。Yassen 等[7]的研究表明,發酵液中泛革素含量受培養基中碳源和氮源影響較大。Islam 等[6]比較了枯草芽孢桿菌C9 菌株在不同碳、氮源組合下的泛革素產量,發現以甘露醇為碳源、以黃豆粉為氮源能提高泛革素的產量。因此,通過發酵培養基優化提高泛革素產量是一種有效途徑。本研究篩選得到了利于HMB19198 菌株芽孢形成的優化培養基,發現在優化培養基中,HMB19198 菌株的泛革素產量提高了39.4%。芽孢桿菌中脂肽類抗生素的產生一般發生在對數生長后期至芽孢形成前期,一些基因同時調控著芽孢的形成和脂肽類抗生素的產生,因此,提高芽孢的形成率往往伴隨著脂肽類抗生素產量的提高[37]。本研究也發現,采用優化后的培養基不僅提高了HMB19198 菌株發酵液中芽孢濃度,同時還提高了泛革素的產量。
