長日照條件下2個熱帶玉米自交系的轉錄組差異分析
2022-06-15 22:59:18高媛王小利涂亮劉鵬飛郭向陽王安貴祝云芳吳迅陳澤輝
南方農業學報 2022年3期
高媛 王小利 涂亮 劉鵬飛 郭向陽 王安貴 祝云芳 吳迅 陳澤輝
摘要:【目的】深度挖掘不同光敏類型自交系間的差異表達基因,為揭示熱帶玉米種質光周期敏感性變異機理提供理論依據,同時為采用分子輔助方法改良熱帶玉米種質提供新途徑。【方法】以熱帶光鈍感自交系QR273和光敏感自交系T32為試驗材料,通過人工控制長日照條件(16 h光/8 h暗)對試驗材料進行處理,于四葉期采集幼嫩葉片提取總RNA,利用RNA-Seq技術對不同光敏類型自交系材料進行轉錄組測序,同時結合生物信息學分析,篩選出不同光敏類型自交系間的差異表達基因。【結果】在長日照條件下,從玉米自交系T32和QR273間共鑒定出6977個差異表達基因,其中T32較QR273的上調表達基因有3291個、下調表達基因有3686個。GO功能注釋分析結果顯示,有24個應答光刺激和應答輻射的基因在T32和QR273中表現出顯著差異;此外,有17個同時與應答光刺激、應答輻射、胚后期發育、光周期、生殖結構發育、生殖系統發育、生殖發育過程、光周期/開花和分生組織營養生長向生殖生長轉變相關的差異表達基因。KEGG信號通路富集分析發現有15個晝夜節律通路相關基因在T32和QR273中表現出顯著差異,其中,Zm00001e030377和Zm00001e023792基因在T32中的表達量高于QR273,而Zm00001e016746和Zm00001e011193基因在T32中的表達量低于QR273。【結論】長日照條件下光敏型與光鈍型熱帶玉米自交系間的轉錄組差異分析共鑒定出6977個顯著差異表達基因,包括24個參與應答光刺激和應答輻射的差異表達基因有,17個同時與應答光刺激、應答輻射、胚后期發育、光周期及分生組織營養生長向生殖生長轉變相關的差異表達基因,以及15個與晝夜節律通路相關的差異表達基因。這些基因通過差異性參與晝夜節律通路中的成花誘導,導致成花啟動途徑不完全相同,可能是熱帶種質光周期敏感性變異的關鍵遺傳基因。
關鍵詞: 熱帶玉米自交系;光周期;差異表達基因;長日照條件;轉錄組測序
中圖分類號: S513.01? ? ? ? ? ? ? ? ? ? ? ? ? ? ? 文獻標志碼: A 文章編號:2095-1191(2022)03-0654-11
Analysis of transcriptome differences between two tropical maize inbred lines planted under long sunshine
GAO Yuan WANG Xiao-li TU Liang LIU Peng-fei GUO Xiang-yang WANG An-gui ZHU Yun-fang WU Xun CHEN Ze-hui
(1Agricultural College, Guizhou University, Guiyang, Guizhou? 550025, China; 2Institute of Upland Crops, Guizhou Academy of Agricultural Sciences, Guiyang, Guizhou? 550006, China; 3Pratacultural Institute, Guizhou Academy of
Agricultural Sciences, Guiyang, Guizhou? 550006, China)
Abstract:【Objective】The differentially expressed genes (DEGs) among different photosensitive inbred lines were explored, so as to provide a theoretical basis for revealing the variation mechanism of photoperiod sensitivity of tropical maize germplasm and a new way of improving tropical maize germplasm by molecular assisted method. 【Method】Tropical germplasm light-insensitive inbred lines of QR273 and light-sensitive inbred line of T32 were used as test materials, which were planted under artificial control conditions of long-days (LD) treatment (16 h light/8 h dark). In the four leaf stage, young leaves were gathered, and their total RNA was extracted. The transcriptome sequencing of different photosensitive inbred lines was carried out by RNA-Seq technology. At the same time, combined with bioinformatics analysis, the DEGs among different photosensitive inbred lines were screened. 【Result】Under LD conditions, a total of 6977 genes showed differentially expression between lines of T32 and QR273, of which 3291 genes showed up-regulated expression in T32 and 3686 genes showed down-regulated expression in T32. GO functional annotation analysis showed that 24 genes responding to light stimulation and radiation showed significant differences in T32 and QR273 and that there were 17 DEGs related to response to light stimulation and radiation, post embryonic development, photoperiod, reproductive structure development, reproductive system development, reproductive development process, photoperiod or flowering and meristem vegetative to reproductive growth. Enrichment analysis of KEGG signaling pathway showed genes related to circadian rhythm pathway had significant differences in expression between T32 and QR273. The expression of Zm00001e030377 and Zm00001e023792 genes in T32 was higher than that in QR273, while the expression of Zm00001e016746 and Zm00001e011193 genes in T32 was lower than that in QR273.? 【Conclusion】A total of 6977 DEGs are identified by transcriptome difference analysis between photo-sensitive and light-insensitive tropical maize inbred lines under long sunshine conditions, of which 24 DEGs involved in response to light stimulation and radiation, and 17 DEGs related to response to light stimulation, radiation, post embryonic development, photoperiod and the transformation from vegetative growth to reproductive growth of meristem, and 15 DEGs related to circadian rhythm pathway. These genes may be the genetic factors leading to the significant differences in photoperiod sensitivity variation among tropical maize germplasms.CB6E18A6-CA0D-42C0-9C0F-28C54F624F8F
Key words: tropical maize inbred lines; photoperiod; differentially expressed genes; long sunshine conditions; transcriptome sequencing
Foundation items: National Natural Science Foundation of China (32171981); Guizhou Province Science and Technology Department (QKHJC〔2020〕1Z09); Guizhou Province High-level Innovative Talent Cultivation Object Project (〔2018〕5629); Science and Technology Innovation Project of Guizhou Academy of Agricultural Sciences (〔2022〕03)
0 引言
【研究意義】玉米作為重要的糧食作物之一,在保障國家糧食安全方面具有重要意義(燕照玲等,2017;曹丹等,2021)。我國的玉米種植分為東華北、西北、黃淮海及南方玉米生態區等,但不同生態區域因光溫差異,所需要的玉米種質也各不相同(劉世夢倪和宋敏,2021)。在我國黃淮海以北地區,主要以塘四平頭、旅大紅骨、蘭卡斯特、改良瑞德等溫帶玉米種質為主(黎裕和王天宇,2010);在南方玉米生態區,由于病蟲害較嚴重,因此對具有較高抗性的Suwan、墨白、CIMMMYT、ETO、巴西等熱帶玉米種質的需求較大。熱帶玉米種質具有籽粒品質優、抗性強、遺傳多樣性豐富等優勢(劉穎等,2020;周海宇等,2020),在現代玉米育種中的應用越來越多。Suwan作為典型的熱帶玉米種質,以其選育自交系如S37和T32等所組配的雜交種在西南地區已得到大面積推廣應用(郭向陽等,2016);但熱帶玉米種質存在光周期敏感性強的缺陷,嚴重影響其在長日照條件下正常開花結實,從而限制在不同生態區的推廣應用(賈小平等,2020;史勇等,2021)。因此,探究長日照條件下熱帶玉米種質光周期變異的遺傳差異,鑒定出具有熱帶背景且光反應鈍感和敏感基因,對于深度揭示熱帶玉米種質光周期敏感性變異的遺傳機制具有重要意義。【前人研究進展】雄花分化作為植物正常開花的重要因素,是影響光周期敏感的主要因子,其分化時期可分為成花誘導、成花啟動和花器官發育等3個階段(劉莉和祝朋芳,2004),其中成花誘導是植物開花的必要步驟,備受植物學研究者的關注。任永哲等(2006)通過比較不同日照條件下玉米的生長差異,發現在長日照條件下玉米四葉期的總葉片數顯著多于短日照條件;吳連成(2008)以熱帶玉米自交系CML288和溫帶玉米自交系黃早四為材料,評價不同日照條件下玉米的光周期敏感時期,結果發現短日照條件下熱帶玉米自交系CML288的光周期敏感時期在第4~7葉期,而長日照條件下CML288的光周期敏感時期在第4~14葉期;周海宇等(2020)研究表明,理想日照時長是促進植物正常花芽分化的重要環境因子。此外,有學者利用溫熱玉米種質的光周期反應差異來研究成花啟動的分子調控機制。研究表明,長日照條件下,編碼生物鐘組分的核心振蕩器基因ZmELF4與ZmTOC1共同正向調節開花促進基因ZmCCA1/ZmLHY,且ZmELF4和ZmTOC1基因在夜間呈現表達高峰后(Alabadí et al.,2001;張少方,2011;Gendron et al.,2012)即促進下游生物鐘輸出基因如ZmGI的表達,致使ZmGI基因在午間出現表達高峰(唐維等,2014;Lee and An,2015),同時轉錄激活ZmCOL/ZmHd1基因,使其在黃昏時達到峰值(Miller et al.,2008;楊爽,2010),從而促進植物開花。吳連成(2008)研究發現,8個光周期敏感基因均表現晝夜表達節律,其中ZmTFL1基因為開花抑制基因,能抑制玉米的成花誘導;Meng等(2011)利用溫熱玉米自交系對ZCN8基因的研究表明,ZCN8基因是唯一具有成花活性的基因,而開花抑制基因ZmCCT、ZmCCT9和ZmCCT10能負反饋調節ZCN8基因表達,進而抑制植物開花;Jin等(2018)以溫帶玉米自交系B73為材料對ZmCOL3基因的研究表明,ZmCOL3是玉米開花抑制因子,在不同光照條件下過量表達ZmCOL3基因可延遲開花約4 h,該基因的表達能反式激活ZmCCT基因的轉錄或干擾晝夜節律,從而抑制植物開花。可見,不同光敏感基因均表現出晝夜表達及互相調控的規律,其研究結果為深度揭示玉米光周期敏感性變異提供了科學依據。【本研究切入點】目前,Suwan玉米種質在現代玉米育種中的重要性日趨凸顯,但對其光周期敏感性變異的遺傳研究較少。【擬解決的關鍵問題】以熱帶玉米Suwan種質選育的骨干玉米自交系T32(光敏感)和QR273(光鈍感)為材料,基于RNA-Seq技術對長光照條件下的玉米材料進行轉錄組測序,深度挖掘不同光敏類型自交系間的差異表達基因,為揭示熱帶玉米種質光周期敏感性變異機理提供理論依據,同時為采用分子輔助方法改良熱帶玉米種質提供新途徑。
1 材料與方法
1. 1 試驗材料
供試材料T32和QR273均選自熱帶玉米種質Suwan 1號群體,由貴州省農業科學院旱糧研究所提供。T32表現出強光敏感特性(光敏指數:株高46.35%,穗位高50.67%),QR273表現出光鈍感特性(光敏指數:株高3.82%,穗位高22.22%)。AxyPrep總RNA小量制備試劑盒(AXYGEN)購自貴州春滿谷生物科技有限公司,GoScriptTM Reverse Transcription System (Promega)和GoTaq? qPCR Master Mix (Promega)試劑盒購自貴州明涵生物科技有限公司。主要儀器設備:移液器(德國Eppendorf公司),高速冷凍離心機(美國Beckman公司),NanoDrop One型超微量紫外分光光度計(美國Thermo公司),反轉錄儀(美國BIO-RAD公司),CFX96 Real-time PCR Detection System(美國BIO-RAD公司),DW-HL540型超低溫冷凍儲存箱(中科美菱科技股份有限公司),JN-GH-01型人工智能氣候室(江南寧波儀器廠)。CB6E18A6-CA0D-42C0-9C0F-28C54F624F8F
1. 2 試驗方法
2020年7月將T32和QR273種植于貴州省農業科學院草業研究所人工氣候室(溫度25 ℃,濕度40%~50%,光照強度4800~6000 lx),長光照條件為16 h光照/8 h黑暗處理。玉米四葉期時,取3片幼嫩葉片等量混合,3個重復,液氮速凍。將葉片樣品分成2份,分別用于轉錄組分析(委托武漢愛基百客公司完成)和實時熒光定量PCR驗證。總RNA提取參照AxyPrep總RNA小量制備試劑盒(AXYGEN)說明進行操作,然后根據GoScriptTM Reverse Transcription System (Promega)試劑盒說明反轉錄合成cDNA。
1. 3 實時熒光定量PCR驗證
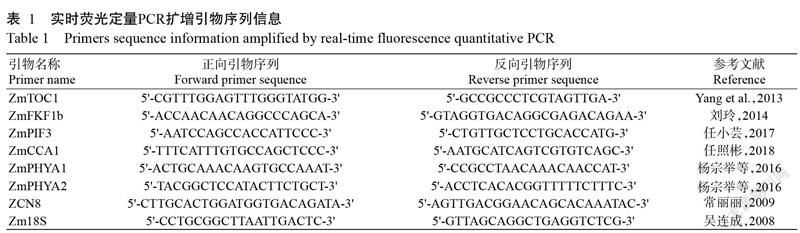
基于轉錄組測序分析結果,挑選7個候選基因進行實時熒光定量PCR驗證。反轉錄合成的cDNA經5倍稀釋后作為模板,以玉米管家基因Zm18S為內參基因,設3次重復。按照GoTaq? qPCR Master Mix試劑盒說明進行實時熒光定量PCR擴增,反應體系20.0 μL:正、反向引物(表1)各0.3 μL,cDNA模板2.0 μL,ddH2O 7.4 μL,SYBR Mix 10.0 μL。擴增程序:96 ℃預變性6 min;96 ℃ 30 s,55 ℃ 30 s,72 ℃ 30 s,進行45個循環;72 ℃讀取熒光值。采用2-ΔCt法換算目的基因相對表達量(王亞麗等,2021),并以Excel 2016進行統計分析及制圖。
2 結果與分析
2. 1 轉錄組測序質量分析結果
6份玉米葉片樣品轉錄組測序獲得的Clean reads介于47024638~56965124條,Clean bases均在6.90 Gb以上,Q30均高于93.00%,GC含量介于57.94%~60.61%(表2)。各樣品中比對到玉米參考基因組的Clean reads占總Clean reads的87.06%~88.47%,在玉米參考基因組中呈唯一比對的Clean reads占總Clean reads的83.38%~84.33%,而在玉米參考基因組中獲得完美匹配的Clean reads占總Clean reads的77.87%~78.68%,表明轉錄組測序數據質量較高,可用于后續研究。
2. 2 差異表達基因篩選結果
長日照條件下,在T32和QR273間共篩選獲得6977個差異表達基因(圖1),其中,T32較QR273的上調表達基因有3291個,下調表達基因有3686個。
2. 3 差異表達基因GO功能注釋分析結果
GO功能注釋分析結果(圖2)顯示,在T32與QR273間的6977個差異表達基因分別注釋到生物學過程(Biological process)、細胞組分(Cellular component)及分子功能(Molecular function)三大功能分類,其中,生物學過程主要涉及細胞過程(Cellular process)、代謝過程(Metabolic process)及應答刺激(Response to stimulus)等,細胞組分主要涉及細胞(Cell)、細胞部分(Cell part)及細胞器(Organelle)等,分子功能主要涉及催化活性(Catalytic activity)、粘合物(Binding)、運輸活性(Transporter activity)及轉錄調節活性(Transcription regulator activity)等。以光周期相關差異表達基因所參與且排名前20的生物學過程進行富集分析,結果(圖3)顯示,在T32與QR273間,參與應答光刺激和應答輻射的差異表達基因最多,為24個;參與胚胎后發育的差異表達基因有22個;參與生殖結構發育、生殖系統發育及生殖發育過程的差異基因有20個;參與營養生長向生殖生長轉變的差異基因有15個。經Peak關聯基因分析,結果(圖4)顯示,有17個同時與應答光刺激、應答輻射、胚后期發育、光周期、生殖結構發育、生殖系統發育、生殖發育過程、光周期/開花和分生組織營養生長向生殖生長轉變相關的差異表達基因;有6個與晝夜節律相關的差異表達基因;有2個同時與應答光刺激、應答輻射、胚后期發育、光周期、生殖結構發育、生殖系統發育、生殖發育過程、光周期/開花、分生組織營養生長向生殖生長轉變和晝夜節律相關的差異表達基因;有2個同時與應答光刺激、應答輻射和晝夜節律相關的差異表達基因;有2個同時與應答光刺激、應答輻射和胚后期發育相關的差異表達基因;有1個同時與應答光刺激、應答輻射、胚后期發育、光周期、生殖結構發育、生殖系統發育和生殖發育過程相關的差異表達基因。
2. 4 差異表達基因KEGG信號通路富集分析結果
KEGG信號通路富集分析結果(圖5)顯示,在T32與QR273間的6977個差異表達基因分別富集到層次結構(Brite hierarchies)、細胞過程(Cellular processes)、環境信息處理(Environmental information processing)、遺傳信息處理(Genetic information processing)和新陳代謝(Metabolism)五大通路途徑,其中,層次結構通路主要涉及家族:信號和細胞過程(Families:Signaling and cellular processes)、蛋白家族:代謝(Protein families:Metabolism)和蛋白家族:遺傳信息處理(Protein families:Genetic information processing),細胞過程主要涉及運輸和分解代謝(Transport and catabolism)、細胞群落原核生物(Cellular community-prokaryotes)及細胞生長死亡(Cell growth and death)等,環境信息處理主要涉及信號轉導(Signal transduction)和膜運輸(Membrane transport)2個通路途徑,遺傳信息處理主要涉及翻譯(Translation)、轉錄(Transcription)及復制和修復(Replication and repair)等通路途徑,新陳代謝主要涉及能量代謝(Energy metabolism)、碳水化合物代謝(Carbohydrate metabolism)及氨基酸代謝(Amino acid metabolism)等通路途徑。同時,以光周期相關的差異表達基因所參與的通路途徑進行富集分析,結果(圖6)顯示,光周期相關差異表達基因主要富集在8個通路途徑中,其中有15個晝夜節律差異表達基因、3個泛素介導的蛋白水解基因及3個內質網蛋白加工差異表達基因達極顯著差異水平;此外,有5個分別參與α-亞麻酸代謝、氨基酸和核苷酸糖代謝、苯丙素生物合成、淀粉和蔗糖代謝、植物激素信號轉導的基因在T32和QR273中無顯著差異。CB6E18A6-CA0D-42C0-9C0F-28C54F624F8F
經Peak關聯基因分析結果(圖7)顯示,在T32與QR273間的6977個差異表達基因中有11個與晝夜節律相關的差異表達基因;有3個與晝夜節律和編碼泛素介導的蛋白質水解通路途徑相關的差異表達基因參;有3個參與植物激素信號轉導的差異表達基因;有1個編碼α-亞麻酸代謝的差異表達基因;有1個同時參與在氨基糖和核苷酸代謝及淀粉和蔗糖代謝的差異表達基因;有1個同時參與泛素介導的蛋白質水解和植物激素信號轉導的差異表達基因;有1個參與苯丙烷生物合成的差異表達基因。
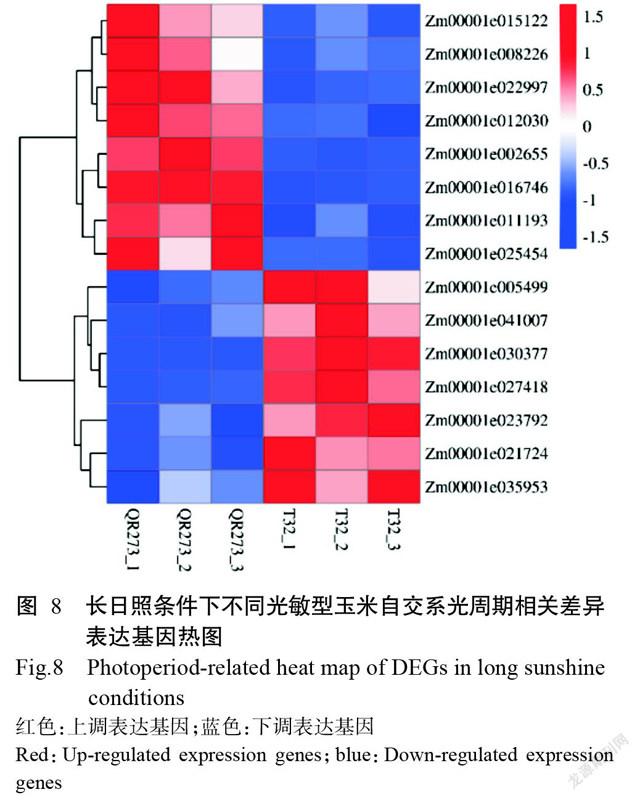
2. 5 光周期相關基因表達差異分析結果
基因表達差異分析(logFC:2個樣品間的表達量比值)結果顯示,15個光周期相關差異表達基因在玉米自交系T32與QR273間的表達同樣存在顯著差異(圖8)。其中,下調表達基因為Zm00001e015122 (TOC1;logFC:1.77354)、Zm00001e008226(TOC1;logFC:1.26039)、Zm00001e022997(TOC1;logFC:1.69316)、Zm00001e012030(PHYC2;logFC:1.03377)、Zm00001e002655(COP1;logFC:3.24929)、Zm0000- 1e016746(HY5;logFC:1.48346)、Zm00001e011193(FKF1;logFC:1.21325)和Zm00001e025454(PIF3;logFC:2.45599);上調表達基因為Zm00001e005499(PHYA;logFC:1.57792)、Zm00001e041007(HY5;logFC:1.44814)、Zm00001e030377(COP1;logFC:5.91744)、Zm00001e027418(FT;logFC:3.96988)、Zm00001e023792(COP1;logFC:1.11024)、Zm0000- 1e021724(CCA1、LHY;logFC:1.61528)和Zm0000- 1e035953(PRR3、PRR5、PRR7;logFC:1.42903)。
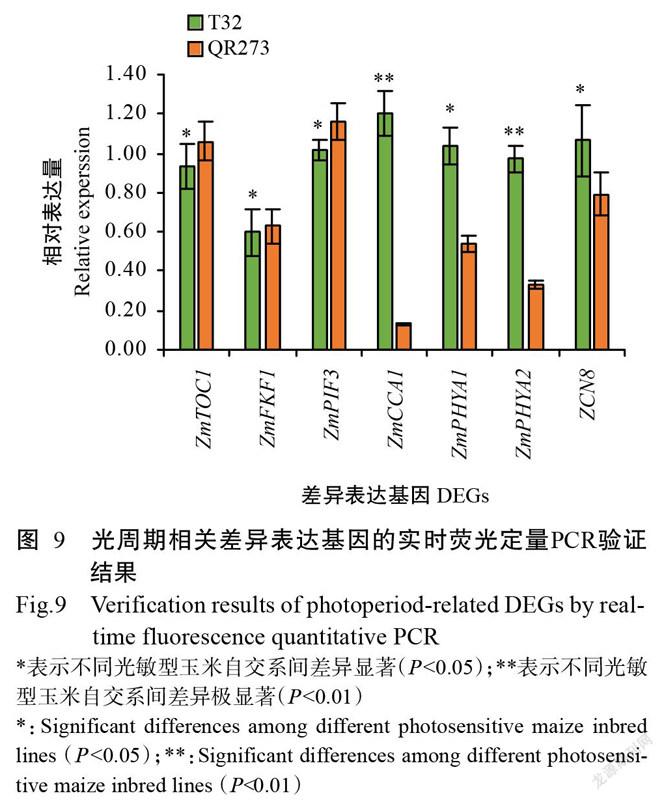
2. 6 實時熒光定量PCR驗證結果
實時熒光定量PCR驗證結果(圖9)顯示,在長日照條件下,ZmTOC1、ZmFKF1和ZmPIF3基因在T32中的相對表達量顯著低于QR273(P<0.05,下同),而ZmPHYA1和ZCN8基因在T32中的相對表達量顯著高于QR273,ZmCCA1和ZmPHYA2基因在T32中的相對表達量極顯著高于QR273(P<0.01),與轉錄組測序結果一致,說明轉錄組測序數據可靠性較高。
3 討論
玉米四葉期是營養生長期,也是成花誘導的開始時期,與成花啟動的分子調控機制不完全相同。本研究結果表明,在長日照條件下,從光敏感自交系T32和光鈍感自交系QR273間共檢測到6977個差異表達基因,主要參與生物學過程、細胞組分和分子功能,其中參與生物學功能的差異表達基因最多。KEGG信號通路富集分析結果顯示,這些差異表達基因主要參與層次結構、細胞過程、環境信息處理、遺傳信息處理及新陳代謝等五大信號通路途徑,其中參與代謝活動的差異表達基因最多。GO功能注釋分析發現,有24個參與應答光刺激和應答輻射的基因在T32與QR273中表現出顯著差異,且有17個同時與胚后期發育、光周期、分生組織營養生長向生殖生長轉變的差異表達基因,與周金龍(2016)、任芹勇(2019)的研究結果相似。此外,本研究結果與T32和QR273的田間表現較一致,即T32表現為在長日照條件下光敏感,不能正常開花結實;QR273則表現為光鈍感,在長日照條件下仍然能正常開花結實(劉穎等,2020),故推測T32與QR273間的差異表達基因是導致其光周期差異的重要因子。
ZmCCA1基因是玉米生物鐘核心振蕩器的關鍵基因,是擬南芥CCA1基因的同源基因,在清晨的表達水平達峰值,能與LHY基因相互作用形成負反饋環,共同抑制TOC1基因表達;TOC1基因又是CCA1和LHY基因的促進因子,夜晚時分CCA1和LHY基因的表達量降低,對TOC1基因的抑制作用減弱,因此,TOC1表達量的增加從而在凌晨促進CCA1和LHY基因表達,形成負反饋調節環(邢國芳等,2011)。ZmGI基因是玉米生物鐘重要的輸出基因,長日照條件下能與FKF1基因形成復合體在光照結束時達峰值,泛素化降解CDFs后轉錄激活下游轉錄調控基因,從而促進植物開花(劉玲,2014)。PRR亞族屬于CCT結構域基因家族,影響植物的開花,CCA1和LHY基因同時轉錄激活PRR9、PRR7和PRR5基因,而在PRR5基因突變體中PRR9和PRR7基因的表達量顯著降低,同時負調控CDF1基因,并抑制CCA1和LHY基因表達,致使生物鐘縮短,PRR9基因或PRR7基因突變則導致生物鐘延長(李劍峰等,2019)。本研究結果表明,多數光周期相關基因在光鈍感自交系和光敏感自交系間的表達存在顯著差異,且表達方向不同。其中Zm00001e015122、Zm00001e008226、Zm00001e022997、Zm00001e012 030、Zm00001e002655、Zm00001e016746、Zm0000-1e011193和Zm00001e025454基因在T32中呈下調表達,Zm00001e00-5499、Zm00001e041007、Zm0000-1e030377、Zm00001e027418、Zm00001e023792、Zm00001e021724和Zm00001e035953基因在T32中呈上調表達,可能與每個基因在應答光刺激過程中所扮演的角色不同存在關聯。如COP1作為一種E3泛素化連接酶,能介導擬南芥光信號轉導中HY5等正向調控因子的泛素化降解,從而影響植株幼苗的光形態建成、花青素合成及開花等生物學功能。Zm00001e030377和Zm00001e023792基因均屬于COP1家族基因,編碼COP1,在光敏感自交系T32中的相對表達量顯著高于QR273,從而延遲開花。酵母雙雜試驗結果證明,COP1負調控HY5,而HY5是堿性亮氨酸拉鏈(bZIP)轉錄因子家族成員之一,可促進植物光形態建成;Zm00001e016746基因編碼HY5轉錄因子,且在T32中的相對表達量顯著高于QR273,從而延遲開花,與駱倩(2014)、崔連花等(2021)的研究完全一致。F-box蛋白FKF1屬于ZTLs藍光受體家族,能與GI相互作用形成復合體,抑制CDF1結合在CO蛋白上,從而促進開花。Zm0000-1e011193基因編碼FKF1蛋白,在T32中的相對表達量低于QR273,故延遲開花,與楊敏等(2021)的研究結果一致。這與T32在長日照條件下不能正常結實,而QR273在長日照條件下可正常結實的表型完全一致,為進一步揭示熱帶玉米自交系光周期成花誘導分子調控機制及后續的熱帶玉米種質光周期遺傳改良提供了參考依據。CB6E18A6-CA0D-42C0-9C0F-28C54F624F8F
4 結論
長日照條件下光敏型與光鈍型熱帶玉米自交系間的轉錄組差異分析共鑒定出6977個顯著差異表達基因(上調表達基因3291個,下調表達基因3686個),包括24個參與應答光刺激和應答輻射的差異表達基因有,17個同時與應答光刺激、應答輻射、胚后期發育、光周期及分生組織營養生長向生殖生長轉變相關的差異表達基因,以及15個與晝夜節律通路相關的差異表達基因。這些基因通過差異性參與晝夜節律通路中的成花誘導,導致成花啟動途徑不完全相同,可能是熱帶種質光周期敏感性變異的關鍵遺傳基因。
參考文獻:
曹丹,張紅梅,楊海鵬,趙歡,劉小紅. 2021. 高溫脅迫下不同玉米材料間的基因差異表達分析[J]. 甘肅農業大學學報,56(1):42-49. [Cao D,Zhang H M,Yang H P,Zhao H,Liu X H. 2021. A differential expression of genes between different maize materials under high temperature stress[J]. Journal of Gansu Agricultural University,56(1):42-49.] doi:10.13432/j.cnki.jgsau.2021.01.007.
常麗麗. 2009. 玉米光周期敏感相關基因ZmTFL1的克隆及功能驗證[D]. 鄭州:河南農業大學. [Chang L L. 2009. Cloning and characterization of ZmTFL1 gene related to photoperiod sensitivity in maize[D]. Zhengzhou:Henan Agricultural University.] doi:10.7666/d.y1574484.
崔連花,詹為民,楊陸浩,王少瓷,馬文奇,姜良良,張艷培,楊建平,楊青華. 2022. 2個玉米ZmCOP1基因的克隆及其轉錄豐度對不同光質處理的響應[J]. 作物學報,48(6):1312-1324. [Cui L H,Zhan W M,Yang L H,Wang S C,Ma W Q,Jiang L L,Zhang Y P,Yang J P,Yang Q H. 2022. Molecular cloning of two maize (Zea mays) ZmCOP1 genes and their transcription abundances in response to different light treatments[J]. Acta Agronomica Sinica,48(6):1312-1324.] doi:10.3724/SP.J.1006.2022. 13040.
郭向陽,陳澤輝,胡興,祝云芳,王安貴,陳建軍. 2016. 熱帶玉米Suwan群體的遺傳特性及育種潛勢[J]. 玉米科學,24(4):41-45. [Guo X Y,Chen Z H,Hu X,Zhu Y F,Wang A G,Chen J J. 2016. Genetic characteristics and bree-ding potential of the good tropical maize Suwan groups[J]. Journal of Maize Sciences,24(4):41-45.] doi:10. 13597/j.cnki.maize.science.20160408.
賈小平,袁璽壘,李劍峰,王永芳,張小梅,張博,全建章,董志平. 2020. 不同光溫條件谷子光溫互作模式研究及SiCCT基因表達分析[J]. 作物學報,46(7):1052-1062. [Jia X P,Yuan X L,Li J F,Wang Y F,Zhang X M,Zhang B,Quan J Z,Dong Z P. 2020. Photo-thermal interaction model under different photoperiod-temperature conditions and expression analysis of SiCCT gene in foxtail millet(Setaria italica L.)[J]. Acta Agronomica Sinica,46(7):1052-1062.] doi:10.3724/SP.J.1006.2020.94144.
黎裕,王天宇. 2010. 我國玉米育種種質基礎與骨干親本的形成[J]. 玉米科學,18(5):1-8. [Li Y,Wang T Y. 2010. Germplasm base of maize breeding in China and formation of foundation parents[J]. Journal of Maize Sciences,18(5):1-8.] doi:10.13597/j.cnki.maize.science.2010.05. 005.
李劍峰,李婷,賈小平. 2019. PRRs家族功能基因的研究進展[J]. 植物遺傳資源學報,20(6):1399-1407. [Li J F,Li T,Jia X P. 2019. Advances on unlocking the functional basis of PRRs family genes[J]. Journal of Plant Genetic Resources,20(6):1399-1407.] doi:10.13430/j.cnki.jpgr. 20190403001.
劉莉,祝朋芳. 2004. 光周期與植物成花誘導[J]. 遼寧農業科學,(3):26-27. [Liu L,Zhu P F. 2004. Relationship between photoperiod and plant floral induction[J]. Liaoning Agricultural Sciences,(3):26-27.] doi:10.3969/j.issn.1002- 1728.2004.03.010.CB6E18A6-CA0D-42C0-9C0F-28C54F624F8F
劉玲. 2014. 玉米光周期途徑中藍光響應節律基因ZmFKF1的分子進化與關聯分析[D]. 成都:四川農業大學. [Liu L. 2014. Molcular evolution and association analysis of ZmFKF1,a clock-controled gene in maize[D]. Chengdu:Sichuan Agricultural University.]
劉世夢倪,宋敏. 2021. 品種改良對玉米單產的貢獻率分析[J]. 河南農業大學學報,55(2):364-371. [Liu S M N,Song M. 2021. Analysis on the contribution rate of varie-ty improvement to corn yield[J]. Journal of Henan Agricultural University,55(2):364-371.] doi:10.16445/j.cnki.1000-2340.20210317.002.
劉穎,吳迅,郭向陽,何兵,王安貴,劉鵬飛,高媛,涂亮,祝云芳,陳澤輝. 2020. 溫熱玉米T 32/齊319 F2:3家系光周期敏感性分析[J]. 種子,39(12):7-12. [Liu Y,Wu X,Guo X Y,He B,Wang A G,Liu P F,Gao Y,Tu L,Zhu Y F,Chen Z H. 2020. Photoperiodic sensitivity analysis of F2:3 family crossed by T32 and Qi319 from tropical regions[J]. Seed,39(12):7-12.] doi:10.16590/j.cnki.1001-4705. 2020.12.007.
駱倩. 2014. 擬南芥PIF3-LIKE1與phyB和COP1互作調控光形態建成的分子機制研究[D]. 上海:上海交通大學. [Luo Q. 2014. COP1 and phyB physically interact with PIL1 to regulate its stability and photomorphogenic development in Arabidopsis[D]. Shanghai:Shanghai Jiao Tong University.]
任芹勇. 2019. 谷子Ci846植物學特性學及轉錄組分析[D]. 呼和浩特:內蒙古農業大學. [Ren Q Y. 2019. Botany characteristics and transcription group analysis in foxtail millet Ci846[D]. Hohhot:Inner Mongolia Agricultural University.] doi:10.27229/d.cnki.gnmnu.2019.000579.
任小蕓. 2017. ZmPIFs基因的克隆、表達及AtPIFs基因的抗旱功能研究[D]. 揚州:揚州大學. [Ren X Y. 2017. Cloning and expression of ZmPIFs and study on the drought resistant function of AtPIFs[D]. Yangzhou:Yangzhou University.]
任永哲,陳彥惠,庫麗霞,常勝合,高偉,陳曉. 2006. 玉米光周期反應及一個相關基因的克隆[J]. 中國農業科學,39(7):1487-1494. [Ren Y Z,Chen Y H,Ku L X,Chang S H,Gao W,Chen X. 2006. Response to photoperiodical variation and the clone of a photoperiod-related gene in maize[J]. Scientia Agricultura Sinica,39(7):1487-1494.] doi:10.3321/j.issn:0578-1752. 2006.07.026.
任照彬. 2018. 玉米光周期敏感性調節因子ZmCCA1互作蛋白的篩選及調控路徑的初步鑒定[D]. 鄭州:河南農業大學. [Ren Z B. 2018. Preliminary identification of pathway and interaction proteins of photoperiod sensitive factor ZmCCA1 in maize[D]. Zhengzhou:Henan Agricultu-ral University.] doi:10.27117/d.cnki.ghenu.2018.000039.
史勇,金維環,任真真,郭莎,董世鳳,溫巖朋,陳彥惠. 2021. 玉米光周期敏感調節機制的研究進展[J]. 玉米科學,29(1):92-96. [Shi Y,Jin W H,Ren Z Z,Guo S,Dong S F,Wen Y P,Chen Y H. 2021. Research advances in regulation mechanism of photoperiod sensitivity in maize[J]. Journal of Maize Sciences,29(1):92-96.] doi:10.13597/j.cnki.maize.science.20210114.
唐維,劉亞菊,張允剛,王欣,后猛,閆會,馬代夫,李強. 2014. 植物GI基因生理功能研究進展[J]. 分子植物育種,12(5):1044-1049. [Tang W,Liu Y J,Zhang Y G,Wang X,Hou M,Yan H,Ma D F,Li Q. 2014. Advances on physio-logical functions of GI gene in plant[J]. Molecular Plant Breeding,12(5):1044-1049.] doi:10.13271/j.mpb.012.001 044.CB6E18A6-CA0D-42C0-9C0F-28C54F624F8F
王亞麗,陳煜東,王益軍. 2021. 高世代回交玉米矮稈種質的轉錄組分析[J]. 江蘇農業學報,37(2):280-288. [Wang Y L,Chen Y D,Wang Y J. 2021. Transcriptome analysis on the advanced backcross population of maize dwarf germplasm[J]. Jiangsu Journal of Agricultural Sciences,37(2):280-288.] doi:10.3969/j.issn.1000-4440.2021.02. 002.
吳連成. 2008. 光周期影響玉米開花轉換的機理研究[D]. 鄭州:河南農業大學. [Wu L C. 2008. Study on the mechanism of floral transition under different photoperiods in maize[D]. Zhengzhou:Henan Agricultural University.] doi:10.7666/d.y1574711.
邢國芳,杜偉建,張雁明,韓浩坤,韓淵懷. 2011. 玉米晝夜節律鐘基因CCA1的克隆及表達分析[J]. 山西農業大學學報(自然科學版),31(4):332-337. [Xing G F,Du W J,Zhang Y M,Han H K,Han Y H. 2011. Cloning and expression analysis of circadian clock gene CCA1 in maize[J]. Journal of Shanxi Agricultural University(Natural science edition),31(4):332-337.] doi:10.3969/j.issn. 1671-8151.2011.04.008.
燕照玲,段俊枝,馮麗麗,陳海燕,齊紅志,楊翠蘋,施艷,任銀玲,劉毓俠. 2017. 玉米抗病毒基因工程研究進展[J]. 南方農業學報,48(12):2136-2144. [Yan Z L,Duan J Z,Feng L L,Chen H Y,Qi H Z,Yang C P,Shi Y,Ren Y L,Liu Y X. 2017. Maize antiviral genetic engineering:A review[J]. Journal of Southern Agriculture,48(12):2136-2144.] doi:10.3969/j.issn.2095-1191.2017.12.04.
楊敏,胥華偉,王翠玲,楊護,魏岳榮. 2021. 利用CRISPR/Cas9技術研究玉米ZmFKF1在開花過程中的作用[J]. 中國農業科學,54(4):696-707. [Yang M,Xu H W,Wang C L,Yang H,Wei Y R. 2021. Using CRISPR/Cas9-media-ted targeted mutagenesis of ZmFKF1 delayed flowering time in maize[J]. Scientia Agricultura Sinica,54(4):696-707.] doi:10.3864/j.issn.0578-1752.2021.04.003.
楊爽. 2010. 玉米光周期敏感相關基因ZmCOL的克隆及功能驗證[D]. 鄭州:河南農業大學. [Yang S. 2010. Cloning and characterization of ZmCOL gene related to photope-riod sensitivity in maize[D]. Zhengzhou:Henan Agricultural University.] doi:10.7666/d.y1728556.
楊宗舉,閆蕾,宋梅芳,蘇亮,孟凡華,李紅丹,白建榮,郭林,楊建平. 2016. 玉米光敏色素A1與A2在各種光處理下的轉錄表達特性[J]. 作物學報,42(10):1462-1470. [Yang Z J,Yan L,Song M F,Su L,Meng F H,Li H D,Bai J R,Guo L,Yang J P. 2016. Transcription characteristics of ZmPHYA1 and ZmPHYA2 under different light treatments in maize[J]. Acta Agronomica Sinica,42(10):1462-1470.] doi:10.3724/SP.J.1006.2016.01462.
張少方. 2011. 玉米光周期敏感基因ZmELF4的克隆及功能驗證[D]. 鄭州:河南農業大學. [Zhang S F. 2011. Cloning and characterization of photoperiod sensitive gene Zm-TFL1 in maize[D]. Zhengzhou:Henan Agricultural University.] doi:10.7666/d.y1999417.
周海宇,程偉東,謝小東,周錦國,覃蘭秋,謝和霞,譚賢杰,江禹奉. 2020. 14份玉米自交系的光周期敏感性分析[J]. 西南農業學報,33(9):1877-1883. [Zhou H Y,Cheng W D,Xie X D,Zhou J G,Qin L Q,Xie H X,Tan X J,Jiang Y F. 2020. Photoperiod sensitivity analysis of 14 maize inbred lines[J]. Southwest China Journal of Agricultural Sciences,33(9):1877-1883.] doi:10.16213/j.cnki.scjas. 2020.9.001.CB6E18A6-CA0D-42C0-9C0F-28C54F624F8F
周金龍. 2016. 玉米光周期敏感近等基因系的轉錄組分析[D]. 鄭州:河南農業大學. [Zhou J L. 2016. Transcriptome analysis of maize photoperiod-insensitive near-isogenic[D]. Zhengzhou:Henan Agricultural University.] doi:10.7666/d.D01117137.
Alabadí D,Oyama T,Yanovsky M J,Harmon F G,Más P,Kay S A. 2001. Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock[J]. Science,293(5531):880-883. doi:10.1126/science. 1061320.
Gendron J M,Pruneda-Paz J L,Doherty C J,Gross A M,Kang S E,Kay S A. 2012. Arabidopsis circadian clock protein,TOC1,is a DNA-binding transcription factor[J]. Proceedings of the National Academy of Sciences of the United States of America,109(8):3167-3172. doi:10. 1073/pnas.1200355109.
Jin M L,Liu X G,Jia W,Liu H J,Li W Q,Peng Y,Du Y F,Wang Y B,Yin Y J,Zhang X H,Liu Q,Deng X,Li N,Cui X Y,Hao D Y,Yan J B. 2018. ZmCOL3 a CCT gene represses flowering in maize by interfering with the circadian clock and activating expression of ZmCCT[J]. Journal of Integrative Plant Biology,60(6):465-480. doi:10.1111/jipb.12632.
Lee Y S,An G. 2015. OsGI controls flowering time by modulating rhythmic flowering time regulators preferentially under short day in rice[J]. Journal of Plant Biology,58(2):137-145. doi:10.1007/s12374-015-0007-y.
Meng X,Muszynski M G,Danilevskaya O N. 2011. The FT-like ZCN8 gene functions as a floral activator and is involved in photoperiod sensitivity in maize[J]. The Plant Cell,23(3):942-960. doi:10.1105/tpc.110.081406.
Miller T A,Muslin E H,Dorweiler J E. 2008. A maize CONSTANS-like gene,conz1,exhibits distinct diurnal expression patterns in varied photoperiods[J]. Planta,227(6):1377-1388. doi:10.1007/s00425-008-0709-1.
Yang Q,Li Z,Li W Q,Ku L X,Wang C,Ye J R,Li K,Yang N,Li Y P,Zhong T,Li J S,Chen Y H,Yan J B,Yang X H,Xu M L.2013. CACTA-like transposable element in ZmCCT attenuated photoperiod sensitivity and accelerated the post domestication spread of maize[J]. Proceedings of the National Academy of Sciences of the United States of America,110(42):16969-16974. doi:10.1073/pnas. 1310949110.
(責任編輯 蘭宗寶)CB6E18A6-CA0D-42C0-9C0F-28C54F624F8F
