不同樹齡對胡楊葉片水勢與光合特征的影響
2022-06-15 02:24:42史智欣武勝利管文軻岳永江何宇翔
新疆農業科學 2022年5期
史智欣,武勝利,管文軻,韓 瑋,岳永江,何宇翔
(1新疆師范大學地理科學與旅游學院/新疆干旱區湖泊環境與資源重點實驗室,烏魯木齊 830054;2. 新疆林業科學院,烏魯木齊 830000)
0 引 言
【研究意義】胡楊(PopuluseuphraticaOliv)是新疆塔里木河流域荒漠河岸林的主要建群樹種[1]。若胡楊幼樹根莖發育不完全,死亡率會較高,胡楊種群生存力下降,隨著胡楊胸徑增大,其根系分布、形態特征和生物量分配均需相應調整以適應較大的側根[2]。胡楊抗逆性主要通過葉片生理指標來體現,葉水勢則是衡量植物水分虧缺,判斷植物抗旱能力的一個重要生理指標,與氣孔導度、二氧化碳濃度、凈光合速率等光合因子存在密切的聯系。研究不同樹齡胡楊葉水勢以及其他光合因子,分析不同樹齡對胡楊葉片水勢與光合特性的影響,進一步研究極端干旱區內不同樹齡胡楊的抗逆機理,對塔里木河下游不同樹齡胡楊的人工更新復壯、規范化種植具有重要意義。【前人研究進展】塔里木盆地灰胡楊幼樹光能利用率、胞間CO2濃度、凈光合速率、蒸騰速率、氣孔導度均隨土壤水分遞減而降低[3],凈光合速率下降的主要原因是非氣孔限制因素。分析艾比湖自然保護區內,由近及遠距河道的3個樣地內胡楊的水分特征、蒸騰特征和耗水規律[4]發現,影響樣地一(水分充足)和樣地二(水分一般)狀態下胡楊凈光合速率和蒸騰速率的主要環境因子是光合有效輻射,生理因子為氣孔導度,水分利用效率;影響樣地三(水分缺乏)胡楊凈光合速率和蒸騰速率的環境因子為光合有效輻射、大氣溫度、相對濕度,生理因子是氣孔導度和水分利用效率。不同樹齡的紅豆杉[5]、檸條[6]、烏柳[7]差異光合特性和水分利用特征明顯,其中5年生紅豆杉幼苗的光合參數強于3年生和4年生紅豆杉;而隨著檸條樹齡的增加,其水勢和光合能力都顯著下降;處于不同生長階段的烏柳則表現出不同的生存適應對策,11年生和25年生烏柳的光合能力與水分利用效率均較強,37年生烏柳的光合能力與水分利用效率均較低,4年生烏柳的光合能力較低,水分利用效率卻較高。【本研究切入點】目前關于胡楊的研究大多還集中在水分利用策略、光合特性等方面,對不同樹齡下胡楊水勢與生理及生態因子的變化研究并不多見。需研究快速生長期不同樹齡對胡楊的葉片水勢日變化特征、光合日變化特征的影響。【擬解決的關鍵問題】研究不同灌水量對胡楊幼齡林葉水勢、土水勢的影響,分析胡楊的生理特征及其抗逆性,為極端干旱區胡楊的人工栽培、規范化種植、更新復壯提供理論基礎。
1 材料與方法
1.1 材 料
研究區位于塔里木河下游尉犁縣境內(39°8'~41°39'N,85°02'~89°15'E),整個區域屬典型的溫帶大陸性干旱荒漠氣候[8],冬季干冷,夏季炎熱,日照總時長2 750~2 950 h,積溫(>10℃)4 040~4 300℃,年均降水量15~45 mm,蒸發量1 800~2 600 mm[9]。區內生態物種脆弱性和不穩定性較強,植物群落組成相對簡單,主要以帶狀和斑塊狀分布的胡楊為主要建群樹種,其次有檉柳(Tamarixramosissima)、蘆葦(Phragmitescommunis)、甘草(Glycyrrhizauralensis)、黑果枸杞(Lyciumruthenicum)等植物[10]。
于2019年5月30日~6月5日在塔里木河下游尉犁縣胡楊自然保護區前期綜合調查選取試驗樣地和不同樹齡胡楊幼樹。2019年6月9日~9月18日,采集數據和測定。
采用樹枝分叉法結合地徑和胸徑數據選取樹齡為3年生、5年生,樹勢一致、長勢良好且無病蟲害的胡楊幼樹各5株作為試驗樣株。
1.2 方 法
于2019年6月9日~9月18日期間,采用配備2 cm×3 cm標準透明葉室的Li-6400XT便攜式光合儀(LICOR,USA)和露點水勢儀(WP4,DECAGON,USA)分別測定胡楊氣體交換參數和葉水勢日動態值(ψ,MPa)。
胡楊葉片采用樹干中部同等大小的成熟葉片,2個樹齡胡楊各標定3片待測葉片,每個待測葉片重復3次測定,計算平均值。于每日06:00~20:00測定,每次測定間隔2 h。輸出生理因子參數有蒸騰速率(Tr,mmol/(m2·s) )、凈光合速率(Pn,μmol/(m2·s ) )、胞間CO2濃度(Ci,μmol/mol)、氣孔導度(Gs ,mol/(m2·s) ),環境因子參數為光合有效輻射(PAR,μmol/(m2·s ) )、大氣CO2濃度(Ca,μmol/mol)、相對濕度(RH,%)、氣溫(T,℃);葉水勢日變化每2 h測定1次,選取樹干中部同等大小的成熟葉片中間部位直徑約1 cm的小塊葉片,放置于露點水勢儀C-52樣品室中,將樣品室與露點水勢儀連接,測定葉水勢值(ψ,MPa)。水分利用效率(WUE,μmol/mmol)和氣孔限制值(Ls)計算公式如下:WUE=Pn/Tr;Ls=1-Ci/Ca。
1.3 數據處理
數據處理與統計分析采用Excel 2019和SPSS 25.軟件進行,繪圖采用Origin 9.0 Pro。其中,對不同樹齡胡楊葉水勢、光合參數間的數據運用單因素方差法進行比較,差異的顯著性水平采用Duncan新復極差法檢驗處理。
2 結果與分析
2.1 不同樹齡胡楊葉片水勢變化特征
研究表明,3年生/5年生胡楊在快速生長期內日變化趨勢不同。其中,6月2種樹齡胡楊幼樹都表現出早晚高午間低的趨勢,06:00葉水勢分別為-0.12和-0.14 MPa,隨后在14:00出現全天最低值-0.29和-0.28 MPa,3年生胡楊幼樹于14:00后,出現迅速攀升勢態,并于20:00達到全天最大值-0.11 MPa,5年生胡楊則維持這一低值至16:00后又迅速上升至-0.12 MPa。7月,2個樹齡胡楊幼樹日變化差異顯著,且5年生幼樹變化幅度明顯大于3年生幼樹,3年生幼樹呈波浪式上升趨勢,5年生幼樹呈現明顯“V”字形。3年生幼樹水勢全天日變化最大值出現在20:00(-0.05 MPa),5年生則出現在06:00(-0.14 MPa)。2個樹齡最低值都出現在14:00為-0.17和-0.39 MPa,20:00葉水勢回升至-0.05和-0.14 MPa。8月,2個樹齡幼樹日變化大致相同,3年生幼樹葉水勢在早晨先呈現出緩慢下降趨勢,并于14:00達到全天峰值低谷-0.17 MPa后又逐步攀升至全天最大值-0.07 MPa;5年生胡楊日變化較快,呈明顯“V”字形,最大值和最小值亦出現于20:00和14:00,分別為-0.12和-0.28 MPa。圖1
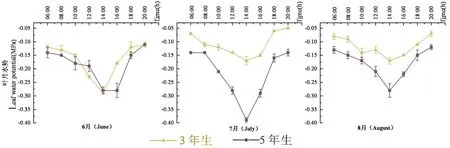
圖1 胡楊葉水勢日變化Fig.1 Daily variation of leaf water potential of P.euphratica in the study area
2.2 不同樹齡胡楊光合參數日變化
2.2.1 外部環境因子日變化特征
研究表明,光合有效輻射整體呈先上升后下降的變化趨勢,08:00光合有效輻射急劇上升,14:00開始下降。光合有效輻射在06:00出現最小值(196.96 μmol/(m2·s ),中午14:00出現最大值(1 217.02 μmol/(m2·s );06:00大氣溫度為1 d中最低時段,為21.65℃,14:00出現最大值,為44.21℃,全天的均溫為35.27℃,日溫差的變化幅度為1~12.83℃。大氣CO2濃度于06:00~12:00急速下降,12:00~20:00呈現波浪式上升趨勢,波動范圍在366~408 μmol/mol,最大值出現在早上06:00,主要因為植物在夜間進行呼吸作用,富集CO2,故08:00大氣CO2濃度較高。空氣相對濕度最大值出現在06:00(59.51%),而后于12:00降至全天最低值(25.84%)后開始緩慢上升,1 d中,空氣相對濕度呈先下降后上升的態勢。
快速生長期內,相對濕度和大氣CO2濃度的變化與光合有效輻射、大氣溫度不同,相對濕度在06:00出現峰值,而此時的光合有效輻射和大氣溫度較低,隨著光合有效輻射和大氣溫度升高,相對濕度逐漸下降。大氣CO2濃度隨著光合有效輻射和大氣溫度升高而降低后又緩慢上升。圖2
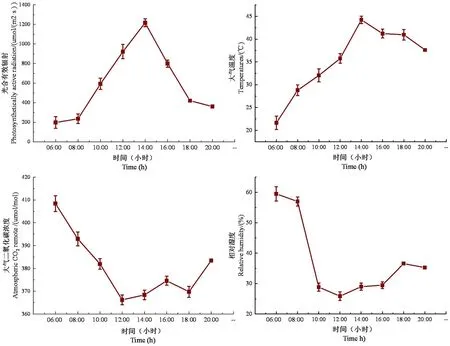
圖2 快速生長期內研究區環境因子日變化Fig.2 Daily variation of environmental factors in the study area during rapid growth period
2.2.2 不同樹齡胡楊光合參數日變化特征
研究表明,2個樹齡胡楊凈光合速率在6~8月均呈現單峰曲線,早晚低,午間高趨勢,這與葉水勢變化趨勢相反。在06:00 2個樹齡胡楊凈光合速率出現全天最低值,中午12:00,6月5年生胡楊達到最大峰值21.69 μmol/(m2·s ),8月5年生胡楊達到最大峰值15.35 μmol/(m2·s ),其他樹齡均在10:00達到最大峰值。
在快速生長期內,2個樹齡胡楊氣孔導度日變化有所差異,其中3年生胡楊呈現為雙峰曲線,5年生胡楊則呈現出單峰曲線,但2種樹齡胡楊氣孔導度振幅第1主峰值均出現在08:00~10:00,10:00后氣孔導度逐漸下降。氣孔開閉行為在不同樹齡,不同時間段均存在差異。8月3年生胡楊在15:00出現低谷值,16:00以后光合有效輻射減弱,胞間CO2回升,氣孔關閉明顯,氣孔導度迅速下降。氣孔導度整體表現為5年生>3年生,6月>7月>8月,且5年生達到快速生長期最大為0.96 mol/(m2·s),比同齡幼樹分別高出45%和63%。
3年生胡楊在6、7、8月均為單峰曲線,5年生胡楊在6、7月表現為雙峰曲線,在8月則變為單峰曲線。相較于凈光合速率,蒸騰速率在12:00~14:00出現峰值,比凈光合速率晚2 h出現,6月,14:00 2個樹齡出現全天最大峰值,且5年生胡楊達到快速生長期最大值22.35 mmol/(m2·s);但在7月,5年生胡楊蒸騰速率于20:00開始緩慢上升。蒸騰速率日均值在10.05~13.85 mmol/(m2·s),日均值較高的是3年生胡楊,且3年生胡楊在8月日變幅最大為0.44 mmol/(m2·s),7月3年生和5年生胡楊蒸騰速率差異極顯著(P<0.01)。
2個樹齡胡楊胞間CO2濃度日變化06:00~10:00迅速下降并在中午時降至最低,于14:00后開始逐漸回升,變化趨勢大致呈“U”字形,與凈光合速率呈負相關,其中6月3年生、8月3年生、6月5年生胡楊幼樹在16:00降至全天最低峰值,其他樹齡則在14:00之前降至最低值。7、8月氣孔限制值變化趨勢與凈光合速率類似,6月2個樹齡均在16:00出現第2次峰值而此時的凈光合速率并未出現波動,且氣孔限制值全天最低值出現在20:00,凈光合速率最低值出現在06:00。氣孔限制值和胞間CO2濃度呈下降趨勢,氣孔導度與二者呈負相關,8月除5年生氣孔導度有微弱上升,其他樹齡氣孔導度均在下降。不同時間段內不同樹齡的光合日變化有所差異,2個樹齡的光合參數,發現主要是氣孔關閉,氣孔限制值增大導致的光合抑制。
2個樹齡胡楊水分利用效率趨勢均為雙峰,主峰值均出現于08:00~10:00,(8月5年生相較于其他樹齡峰值提前1 h)次峰值顯著低于主峰值,日均值變化在0.97~1.65 μmol/mmol。5年生胡楊水分利用效率顯著高于3年生,且在8月達到快速生長期最大值,最大峰值為4.89 μmol/mmol。快速生長期內水分利用效率為8月5年生>7月3年生>7月5年生>6月3年生>6月5年生>8月3年生。圖3
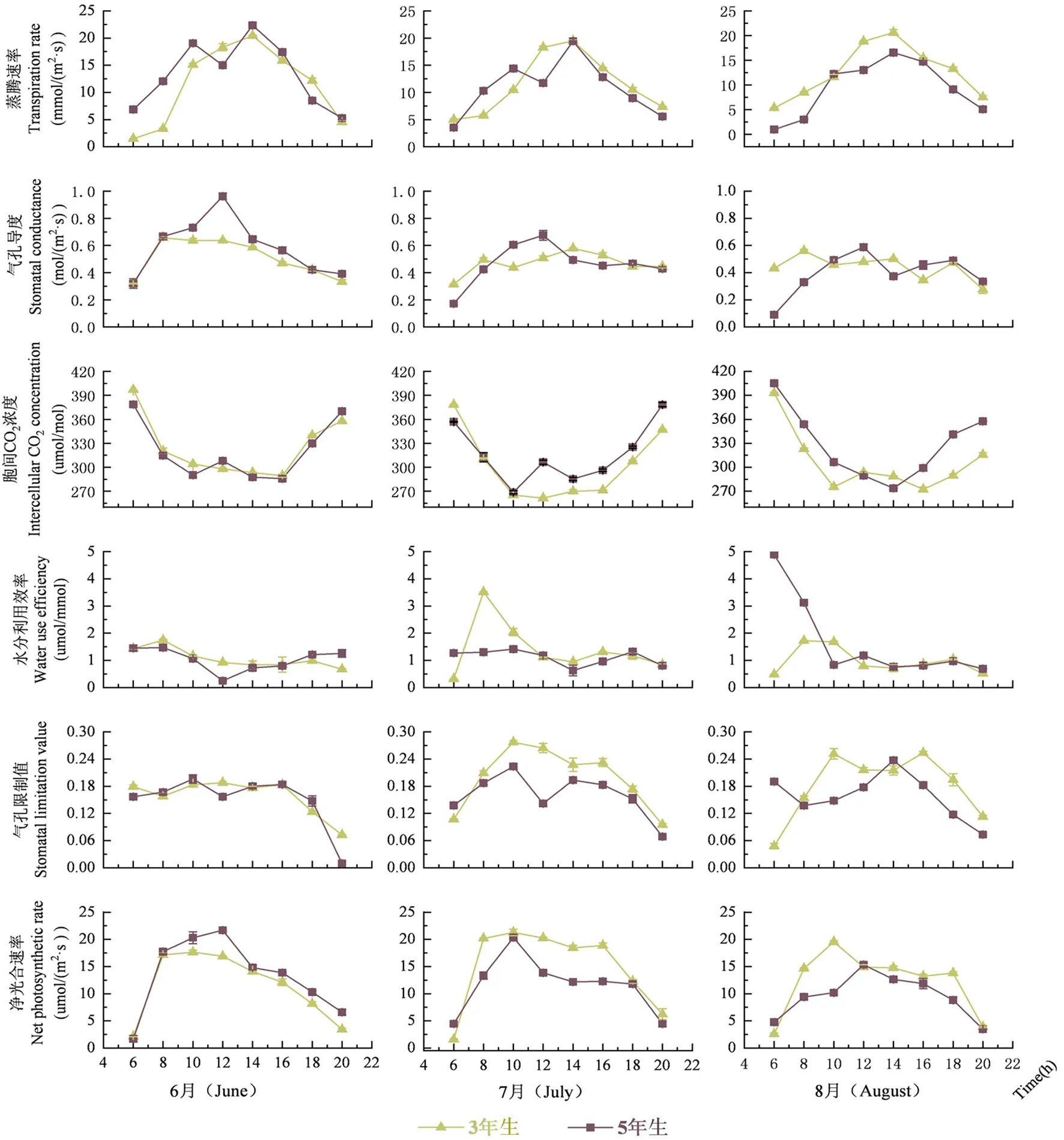
圖3 快速生長期內不同樹齡胡楊光合特性指標日變化Fig.3 Diurnal variation of photosynthetic characteristics of P.euphratica at different ages during rapid growth period
2.3 不同樹齡胡楊葉片水勢相關性
2.3.1 與外部環境因子相關性
研究表明,快速生長期內葉水勢與大氣溫度,光合有效輻射值呈負相關,與大氣CO2濃度,相對濕度呈正相關。其中6月5年生和7月5年生胡楊葉水勢與大氣溫度呈極顯著相關,6月3年生,8月3年生葉水勢與大氣溫度達到顯著相關,7月3年生,8月5年生葉水勢與大氣溫度沒有相關性;快速生長期3年生胡楊葉水勢與光合有效輻射相關性顯著,5年生胡楊光合有效輻射和葉水勢沒有相關性。6月5年生,7月5年生胡楊大氣CO2濃度與葉水勢呈現相關性,6月3年生,8月5年生相對濕度與葉水勢顯著相關。表1
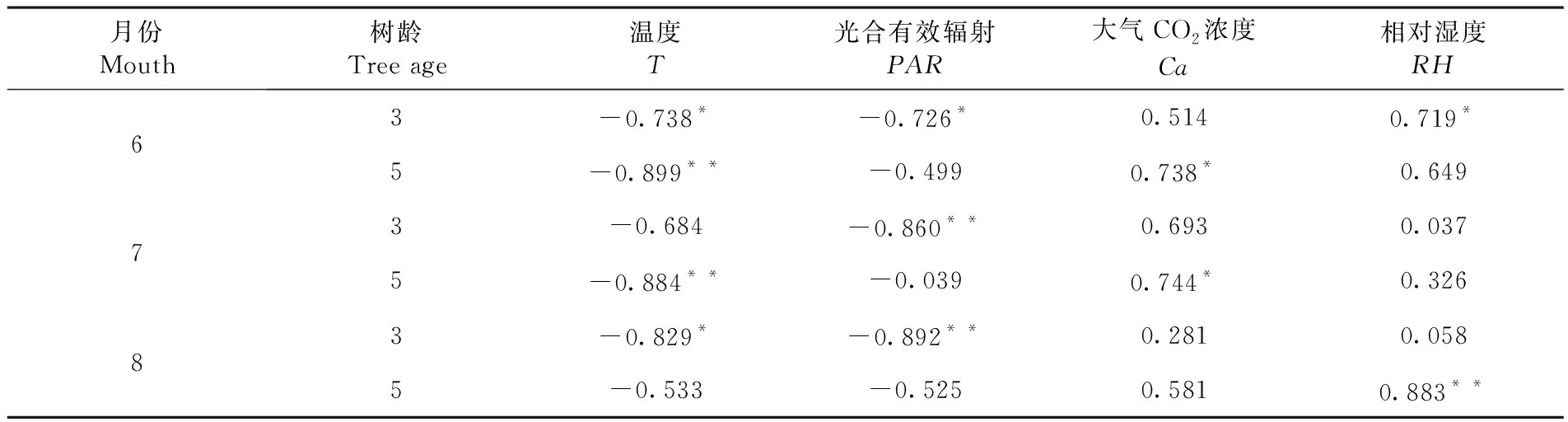
表1 葉水勢與環境因子相關系數Table 1 Correlation coefficient between leaf water potential and environmental factors
2.3.2 與內部生理因子相關性
研究表明,6月5年生幼樹葉水勢與蒸騰速率顯著相關,與胞間CO2濃度極顯著相關,而3年生胡楊僅與蒸騰速率達到顯著相關;7月3年生胡楊葉水勢與胞間CO2濃度極顯著正相關,與蒸騰速率、凈光合速率、氣孔限制值極顯著負相關,與水分利用效率沒有明顯相關性,而5年生胡楊僅與蒸騰速率顯著相關;8月2個樹齡胡楊葉水勢都與蒸騰速率顯著相關,與凈光合速率極顯著相關,其中3年生胡楊又與氣孔限制值顯著相關,5年生胡楊與胞間CO2濃度顯著相關,與氣孔限制值極顯著相關。表2
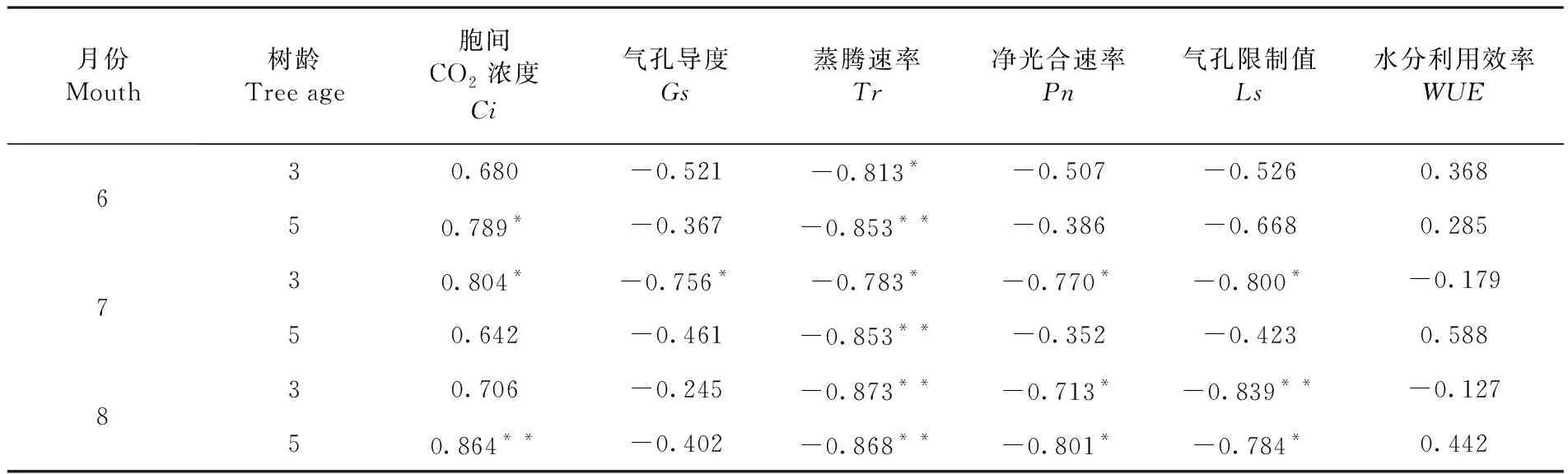
表2 葉水勢與光合生理因子相關性Table 2 Correlation between leaf water potential and physiological factors
3 討 論
植株水勢能夠反映土壤、植被和大氣對植物體內水分可利用性的綜合影響[11]。水勢變化特征既可以反映胡楊對環境的適應,又能反映環境對胡楊的影響程度[12]。受內部、外部光合因子的雙重影響,2個樹齡胡楊葉水勢日變化表現不同,說明樹齡對胡楊葉水勢與光合特性均有一定影響。尹立河等[13]研究表明,生長季小葉楊的葉水勢日變化曲線均呈單峰型;羅歡等[14]對荒漠河岸林胡楊的研究表明,氣候因子是影響胡楊葉水勢的關鍵性因子,胡楊葉水勢的變化與輻射照度的變化呈負相關關系。在極端干旱區衡量植物水分生產能力、篩選抗旱植物的重要指標是水分利用效率[15]。研究發現,3年生胡楊幼樹水分利用效率顯著低于5年生,與Donovan[16]發現幼齡灌木水分利用效率低于成熟灌木,Cavender等[17]關于成熟紅櫟的水分利用效率高于幼樹的研究結果一致。試驗發現3年生胡楊幼樹葉水勢卻明顯大于5年生,且水分利用效率與葉水勢呈現反比線性關系,樹齡較大的胡楊對水分利用并不靈敏,這可能是由于大齡胡楊可以更有效的利用其根系從母體和深層土壤獲得更多的水分以保持較高的水分利用效率,從而保持葉水勢日動態波動并不如3年生明顯,與鮑婧婷等[7]研究發現植物年齡越小,水分利用效率通常越低,且較低的水分利用效率利于建植初期植物的根系生物量的增加,這樣促使幼齡植物的根系達到較深土層,從而吸收有限的水分,促進生長的結果較為一致。
葉水勢可以反映植物進行光合能力的大小[18],反之,植物光合作用的產能亦能揭示出胡楊葉水勢的強弱。丁俊祥[19]、曹生奎[20]等研究發現植物葉水勢主要受光合有效輻射和氣孔導度影響較明顯。對比2個樹齡胡楊光合參數發現,氣孔開閉行為在不同樹齡,不同時間段均存在差異,3年生胡楊表現為雙峰曲線,5年生胡楊則為單峰曲線,2個樹齡胡楊氣孔導度的峰值集中于08:00~10:00出現,10:00后氣孔導度輕微震蕩,整體呈現下降趨勢。氣孔導度是蒸騰過程中水蒸氣從體內排到體外的主要出口,也是光合作用和呼吸作用與外界氣體交換的通道,影響著蒸騰、光合、呼吸等作用過程[21]。隨著樹齡的增大,氣孔發育愈加健全,氣孔的開閉行為對外界生態因子的影響愈加明顯,氣孔開閉行為引起的胞間CO2濃度、氣孔限制值和凈光合速率的變化。植物光合作用存在非氣孔限制因素和氣孔限制因素[22],當胞間CO2濃度降低,若此時氣孔限制值上升,則表示由于氣孔導度下降導致的大氣CO2供應受限引發的凈光合速率下降,導致光合作用減弱的主要原因是氣孔限制因素;當胞間CO2濃度升高,若此時氣孔限制值下降,則是由于氣孔導度升高導致的光合機構和葉功能受到損害引起的光合作用減弱[23]。研究發現,2個樹齡胡楊幼樹對光合箝制的表現各有不同,快速生長期內,2個樹齡凈光合速率均為單峰曲線,無明顯光合午休現象,但3年生氣孔導度為雙峰曲線,5年生為單峰曲線,胞間CO2濃度與氣孔限制值表現亦有差異。在中午12:00~14:00,2個樹齡胞間CO2均為下降趨勢(除7月5年生),此時氣孔限制值呈現下降,氣孔導度關閉,說明此時的光合箝制不單純受外界環境影響,與自身生理、樹齡等面對干旱環境時所做出的抗逆策略關系更為突出,從蒸騰速率變化可以看出,6~7月,5年生幼樹蒸騰速率變化為雙峰(其他時間2個樹齡均為單峰),有明顯光合午休現象,光合有效輻射降低,摒棄生理、樹齡等因素分析氣體交換參數整體趨勢發現,2個樹齡胡楊幼樹均為氣孔限制因素造成光合作用減弱。
4 結 論
4.12個樹齡胡楊在快速生長期內葉水勢日動態均呈現出早晚高,午間低趨勢,樹齡越小對應的葉水勢越高,樹齡不同,其光合因子變化亦有所差異,其中葉水勢與水分利用效率呈負相關關系。
4.2快速生長期2個樹齡胡楊葉水勢均與蒸騰速率呈極顯著正相關(P<0.01),與外部環境因子相關性有所差異。其中 5年生胡楊均與環境因子大氣CO2濃度顯著相關,與大氣溫度極顯著相關;3年生胡楊均與環境因子大氣溫度顯著相關,與光合有效輻射極顯著相關。
4.3樹齡不同,其光合因子變化亦有所差異,5年生胡楊胞間CO2濃度、氣孔導度、水分利用效率日均值顯著高于3年生(P<0.05)。
4.4快速生長期內2個樹齡胡楊幼樹均表現由氣孔限制因素造成光合作用減弱。
