植物多糖的益生作用及其影響因素研究進展
2022-07-02 03:49:36楊斯惠曹亞楠任遠航彭鐮心時小東
食品科學 2022年11期
楊斯惠,向 月,曹亞楠,*,任遠航,彭鐮心,*,時小東
(1.成都大學 農業農村部雜糧加工重點實驗室,四川 成都 610106;2.四川省雜糧產業化工程技術研究中心,四川 成都 610106)
多糖是由超過10個單糖基經過-或-糖苷鍵聚合、脫水形成的天然高分子化合物,主要分布于動物、植物、藻類及微生物中,可以根據其化學成分、在植物體內的功能、來源、溶解性等進行分類。根據來源的不同,可將多糖劃分為動物多糖、植物多糖和微生物多糖(真菌多糖、細菌多糖)。植物多糖作為植物體的重要組成物質,與蛋白質、核酸、脂類并列為植物生命體四大基本物質。常見的植物多糖有淀粉、纖維素、葡聚糖、果膠、果聚糖等。隨著越來越多的科研工作者投入對植物多糖的研究中,其在加工、結構和生物活性方面受到關注。研究表明,植物多糖在抗氧化、抗腫瘤、抗肝損傷、降血糖、益生作用等方面具有顯著優勢。其中,益生作用是多糖生物活性的一個重要方面。
根據推測,人體內存在約500~1 000種細菌,這些微生物總共構成了100萬億個細胞,是自身細胞數量的10 倍,而大多數微生物在腸道生存。這些在人和動物腸胃中生活的微生物統稱為腸道菌群。健康個體的腸道菌群中,大多數微生物分布在厚壁菌門(Firmicutes)、擬桿菌門(Bacteroidetes)、放線菌門(Actinobacteria)、變形菌門(Proteobacteria)、螺旋體門(Spirochaetes)、梭桿菌門(Fusobacteria)、藍藻菌門(Cyanophyta)、疣微菌門(Verrucomicrobia)以及黏膠球形菌門(Lentisphaerae)這9 類菌門中,其中至少90%的細菌屬于厚壁菌門和擬桿菌門。絕大多數微生物群的定植始于生命早期,在新生兒出生時和出生后不久,新生兒接觸了母體和環境中的微生物,同時腸道菌群的建立啟動。嬰兒發育、腸道形態和飲食變化等因素影響嬰兒腸道菌群的變化,結果表現為兼性厭氧菌數量減少,嚴格厭氧細菌逐漸增加成為優勢菌種。哺乳期結束后,腸道菌群趨于穩定,健康人體內形成終生的菌群特征。正常機體內,腸道菌群能夠代謝宿主自身不能降解的多糖,增強腸道上皮屏障功能,減輕炎癥反應,調節免疫反應,抑制致病菌群生長的功能。然而當微生物受到環境、年齡、飲食、生活方式、抗生素等各類因素影響時,菌群動態平衡被打破,進而導致菌群混亂。而通過在飲食中補充多糖調節腸道微生物區系也逐步被人們認識到是有效方法。
據報道,植物多糖(淀粉除外)不能被人體直接消化,但可作為益生元通過抑制病原體和刺激免疫系統來維持機體胃腸道健康,被腸道菌群分解利用,生成具有生物活性的產物(如短鏈脂肪酸(short-chain fatty acids,SCFAs)),調節腸道微生物組成和活性,進而促進人體健康。而多糖的生理功能又與其單糖組成、分子質量、分子形狀和鏈構象等結構密切相關。當前隨著對植物多糖生物活性深入研究,其在醫學、食品、生命科學等領域廣泛運用,成為國內外研究熱點。本文系統論述植物多糖與其益生作用的關系以及影響植物多糖結構的因素,旨在為植物多糖的精準化開發利用提供參考。
1 植物多糖的益生作用
植物中非淀粉類多糖在唾液、胃液和腸液等消化液中不能被消化或完全消化,其中不能被宿主自身酶類催化水解的植物多糖進入腸道,成為人體內腸道微生物區系的主要碳源。利用碳水化合物活性酶類(carbohydrate-active enzymes,CAZymes)的交互降解作用是腸道菌群利用植物多糖的主要途徑。腸道菌群編碼的CAZymes主要包括糖苷水解酶(glycoside hydrolases,GHs)、糖基轉移酶(glycosyltransferases,GTs)、多糖裂解酶(polysaccharide lyases,PLs)、碳水化合物酯酶(carbohydrate esterases,CEs)和碳水化合物結合模塊(carbohydrate binding modules,CBM)。據報道,腸道菌群分解利用多糖的能力與其編碼的CAZymes數量密切相關,同一類型但源于不同菌種的CAZymes活力存在差異,因此,不同菌屬或菌株分解植物多糖的能力可能不同,不同的植物多糖來源還會影響降解微生物中基因表達的特異性。在結腸環境中,植物多糖在腸道菌群的作用下通過不同的降解系統和轉運系統被降解成單糖或低聚糖,隨后這些單糖或低聚糖被轉運至胞內進一步降解和發酵,產生氣體(氫氣、二氧化碳和甲烷)、熱量和SCFAs。SCFAs是腸道微生物群酵解植物多糖的產物中研究最多的。醋酸、丙酸和丁酸是糞便中最常檢測到的SCFAs,是不同細菌的代謝產物。許多細菌可以產生醋酸,但丙酸和丁酸通常是由特定的細菌產生的。在胃腸道環境中,丁酸的主要產生菌是厚壁菌門(如毛螺菌科和普氏桿菌),而丙酸是由類桿菌、陰性桿菌和一些梭狀芽孢桿菌產生的。SCFAs在人體內如何通過碳水化合物代謝途徑獲得以及發揮何種生理功能已有較為詳細的敘述。研究表明,植物多糖對腸道菌群組成具有調節作用,能夠促進有益細菌生長,產生SCFAs,降低糞便pH值,抑制有害菌生長(圖1)。
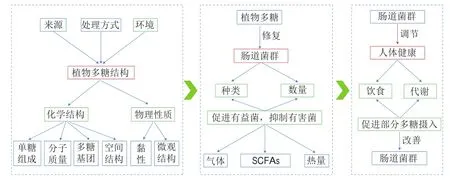
圖1 植物多糖對腸道菌群和人體健康的調節作用Fig.1 Regulatory effect of plant polysaccharides on intestinal flora and human health
由表1可知,竹筍多糖、枇杷葉多糖、黃芪多糖、黨參多糖、白術多糖、大粒車前子多糖、菊粉型果聚糖和枸杞多糖均能促進雙歧桿菌的增殖,果膠多糖和枇杷葉多糖可以顯著增加巨球藻屬和脫硫弧菌屬的數量,燕麥多糖和龍眼多糖則能促進干酪乳桿菌的增殖。這些益生菌相對豐度改變影響著代謝產物SCFAs的生成。植物多糖的來源具有多元性,而不同的植物多糖針對不同的益生菌作用不同,這可能與多糖高度復雜的結構、物理特性密切相關。

表1 植物多糖的益生作用Table 1 Prebiotic effects of plant polysaccharides

續表1
2 植物多糖結構與腸道菌群的關系
2.1 化學結構
植物多糖是結構形式復雜的高分子化合物。植物多糖可以分為一級結構、二級結構、三級結構和四級結構。一級結構包括單糖的組成、糖苷鍵的連接方式和順序、異頭碳的構型等;二級結構是指多糖主鏈之間通過氫鍵等次級鍵而形成的聚合體結構;類似于其他生物大分子,三級和四級結構是指在多糖的二級糖單位結構的基礎上,通過非共價作用在有序空間里產生的規則構象。近年來,從植物中提取得到的多糖在醫學、食品、生命科學等領域引起了廣泛關注,其具有多種生理活性如抗腫瘤、免疫調節、降血糖、益生作用等。這些生理活性受其分子質量、鏈構象、糖苷鍵類型等結構的影響很大。
2.1.1 單糖組成
植物多糖的單糖組成與益生元活性具有相關性。單糖組成能夠影響多糖的鏈結構與高級結構,而高級結構是影響多糖生物活性強弱的一個重要因素。由表1可知,大部分具有益生作用的多糖都具有半乳糖、阿拉伯糖、葡萄糖、鼠李糖、半乳糖醛酸和甘露糖,還有少數具有木糖和果糖。王軼帆等報道了由不同單糖殘基組成的雜多糖——龍眼多糖與由相同單糖殘基組成的同多糖——燕麥多糖的結構特征及其益生活性比較,發現單糖組成更為復雜的龍眼多糖相較于主要由葡萄糖和少量甘露糖組成的燕麥多糖促益生菌增殖效果更佳,推測可能是由于雜多糖作為碳源被益生菌消耗較慢,有利于維持益生菌活菌數穩定。以半乳糖和半乳糖醛酸為基本構成單元的低聚半乳糖具有多種生物活性,同時具有較強的耐酸性和耐熱性,不會在高溫殺菌的加工過程中被破壞,也很少因人體胃酸分解而失去活性。其中,低聚半乳糖的益生活性與地錦草表現出的治療腹瀉等功效相符,李勝男推測由大量半乳糖組成的地錦草多糖可能也具有低聚半乳糖的益生活性。Zhu Rugang等報道了獼猴桃果實中果膠低聚糖(pectin oligosaccharides,POSs)與抗糖化潛力的構效關系,發現POSs的益生元活性與分支度((阿拉伯糖)∶(鼠李糖)和(半乳糖)∶(鼠李糖))以及中性糖含量呈正相關,特別是半乳糖和阿拉伯糖。從阿月渾子殼和竹筍中提取的由阿拉伯糖、半乳糖和木糖組成的多糖是潛在益生元也印證了多糖的單糖組成與益生元活性具有相關性這一觀點。
2.1.2 分子質量
分子質量在一定范圍內的多糖具有生物活性,分子質量過大或過低的多糖一般不具有明顯的生物活性。究其原因,是因為活性多糖通過跨越細胞膜進入生物體內發揮生物學活性,而分子質量的大小影響了多糖的溶解性和黏度,與多糖跨越細胞膜障礙相關,分子質量太大不利于實現這個過程。與分子質量278、354 kDa的阿拉伯木聚糖相比,分子質量66 kDa的阿拉伯木聚糖對于益生菌生長具有更好的增殖作用。Chen Guangjing等報道了不同提取方法對竹筍多糖益生元活性影響的研究,探究提取工藝對竹筍多糖的體外消化率、益生元活性和理化性質的影響。結果發現,超聲輔助提法得到的竹筍多糖(U-CPS)和酶解法得到的竹筍多糖(E-CPS)的分子質量均顯著低于另外3種多糖。同時體外研究發現,分子質量和粒徑較小的U-CPS和E-CPS可能更利于被細菌細胞轉運和代謝,刺激SCFAs的生成,從而產生更好的益生作用。Huang Fei等發現分別采用熱水、超細粉碎和超細粉碎輔助酶法提取得到的龍眼果肉多糖LP-H、LP-S和LP-SE對益生菌增殖有積極作用,且分子質量較小的LP-SE對益生菌增殖作用最強。Fu Yuan等利用7種方法(熱水提取、加壓水提取、高速剪切均質提取、微波輔助提取、超聲輔助提取、超聲波輔助酶法提取和超聲-微波輔助)提取枇杷葉多糖,其中超聲輔助提取和超聲-微波輔助提取的枇杷葉多糖具有較強的益生作用,這可能與其具有較低的分子質量和黏度有關。張宇等通過灌胃鹽酸林可霉素建立腸道微生態失調小鼠模型,然后利用不同分子質量的黃芪多糖進行治療,結果表明黃芪多糖分子質量在6~10 kDa范圍內有利于“扶持”有益菌,抑制有害菌,調節小鼠腸道微生態菌群平衡效果最佳。
2.1.3 多糖基團
多糖基團數量、位置和種類的改變會導致生物活性的改變。相關研究表示,黃秋葵花多糖能夠顯著促進大腸桿菌發酵液中乙酸和己酸的產生。而進行羧甲基化、硫酸酯化和乙酰化結構修飾后,與黃秋葵花多糖相比,修飾多糖顯著抑制大腸桿菌發酵液中乙酸、丁酸和己酸的產生,這可能是因為修飾多糖不能被大腸桿菌酵解發酵產生SCFAs。王亞莉以葡聚糖硫酸鈉誘導的小鼠潰瘍性結腸炎為模型,研究松花粉多糖和硫酸酯化松花粉多糖對葡聚糖硫酸鈉組小鼠腸道菌群的影響,發現兩種多糖在調節腸道菌群平衡方面都起到積極作用。王鑫以菜籽為原料,經提取、分離和純化得到均一多糖R/MIN1-a,然后對R/MIN1-a分別進行硫酸化修飾、乙酰化修飾和羧甲基化修飾,研究R/MIN1-a和修飾后多糖對腸道益生菌體外增殖活性的影響。結果表明,與R/MIN1-a相比,硫酸化修飾產物對嗜酸乳桿菌、嬰兒雙歧桿菌、青春雙歧桿菌和兩歧雙歧桿菌的增殖作用更為明顯。Nuerxiati等對廣布紅門蘭多糖進行硫酸化、乙酰化、磷酸化和羧甲基化修飾,發現不同的改性多糖對不同的益生菌增殖效果不同;在保加利亞乳桿菌增殖效果的比較中,磷酸化多糖表現最佳,乙酰化多糖對青春雙歧桿菌的生長促進效果最好,硫酸化多糖則保持適度穩定的益生菌能力。
2.1.4 空間結構
特定的空間結構是發揮多糖生物活性的必要條件。作為影響多糖生物活性強弱的一個重要因素,多糖的空間結構復雜,是糖鏈之間或者糖鏈內部氫鍵、范德華力、色散力和疏水性等非共價鍵共同作用的結果,并具有多種構象,例如單螺旋、雙螺旋、三螺旋和無規卷曲等。在多糖空間結構的分析測定上,常用的方法有剛果紅實驗、X射線衍射和原子力顯微鏡分析等。一般認為,多糖具有規則的空間結構,則生物活性也強。研究發現,多糖生物活性的基礎是-(1→3)--葡聚糖的單股螺旋構象。例如,-(1→3)--葡聚糖發揮免疫增強活性主要依賴于單螺旋構象和位于螺旋外表面的親水性基團的存在,三股螺旋結構對其活性的發揮來說并不是必須的。而Maeda等認為,三股螺旋構型對有些食用菌-(1→3)-葡聚糖的免疫刺激和抗腫瘤活性有重要作用。雖有較多文獻報道多糖空間結構與活性的關系,也取得了重要進展,但由于植物多糖的復雜性和高度多態性,許多精細結構仍未得到準確解析,相關研究結果仍存爭議。植物多糖益生作用雖已被證實,但其空間結構與益生活性的關系鮮有報道,制約了植物多糖的精準化開發利用。
2.2 物理性質
2.2.1 黏度
多糖的黏度由溶解度和分子質量所決定。而黏度與非淀粉多糖中的單糖組成和鍵型沒有相關性。在較低濃度下,多糖會與水分子直接結合,黏度隨之增加;在較高濃度下,多糖相互結合和共價或非共價鍵連接形成網狀物,更易于增加腸道內容物的黏度。一般來說,分子質量越大、分子結構越復雜的多糖增加消化道中液體黏度的作用愈明顯。而研究表明,水溶性高和黏度低的多糖可以更容易、快速和完全地被益生菌利用。
2.2.2 微觀結構
多糖的微觀結構受單糖組成的影響,隨單糖組成復雜程度、數量和連接方式的改變,因此也會有所不同。用掃描電子顯微鏡觀察多糖的形態可以有效地分析多糖的結構和形態,包括大小、分子形狀和孔隙率。Xu Shuying等分析了人類腸道菌群對海壇紫菜多糖的微生物分解代謝,通過掃描電子顯微鏡觀察發現,海壇紫菜多糖的結構可能由較不易被腸道微生物降解利用的外層和容易被腸道細菌降解的內層組成,從而增加了SCFAs的水平。
3 影響植物多糖結構的因素
植物多糖益生作用受到學者們廣泛關注,以植物多糖為原料的益生產品開發成為熱點。盡管其益生機制尚未明晰,植物多糖結構與物理性質關系到其活性強弱。多糖的結構受到來源、處理方式、環境、溫度和其他因素的影響(圖1)。認識改變植物多糖結構的關鍵因素并合理利用,可為植物多糖開發利用提供重要參考。
3.1 提取方法
針對植物多糖的提取,常用的方法有熱水浸提、酸提法、堿提法、醇提法、超聲波提法、酶解法等。
熱水浸提是最為經濟、便捷和常見的多糖提取方法,能夠較好保存分子結構,但提取耗時長,效率低,得到的多糖大多是中性多糖,還有可能降解多糖。酸提法原理是在一定溫度、時間和pH值條件下,細胞膨脹并破壞細胞壁,植物多糖從中釋放。酸提法具有提取率高、純度高、耗時短的優點;缺點是具有特異性,不適用于一般植物多糖的提取,并破壞糖苷鍵。堿提法原理與酸提法相似,適合提取含有糖醛酸的多糖及酸性多糖,有利于提取率的提高。酶解法是通過酶對細胞壁通透性的改變,減少溶劑提取時來自細胞壁、細胞膜和細胞間質的阻力,使其軟化、膨脹、破裂,從而使內容物流出。此法具有反應條件溫和、工藝簡單、生物活性高等優點,但與傳統水提醇沉法相比,成本較高。
張紅運研究了不同提取工藝對大豆種皮多糖結構的影響,利用微波輔助提取法和熱水浸提法提取多糖,發現微波輔助提取有利于大分子多糖的提取,但具有多糖分子質量分布范圍較廣、均一性差等缺點,同時還會降低分支鏈分子質量。馬智玲分別采用酸法、酶法和鹽法對馬鈴薯果膠進行提取,結果表明,不同提取方法下,果膠的單糖組成和分子質量不同,提取劑的性質和沉析方式也會對果膠的結構造成影響。Chen Guangjing等探究了提取工藝對竹筍多糖理化性質的影響。結果發現,不同提取工藝下,5種多糖的單糖組成相同,但各單糖的含量不同;超聲輔助提法得到的竹筍多糖和酶解法得到的竹筍多糖的分子質量均顯著低于另外3種多糖。Huang Fei等采用熱水、超細粉碎和超細粉碎輔助酶法提取龍眼果肉多糖,結果表明,3種提取方法對多糖的化學組成和結構均有一定影響。與熱水、超細粉碎法相比,超細粉碎輔助酶法提取多糖的表觀黏度、粒徑、分子質量最低。
3.2 分離、純化
多糖經過提取后一般還夾雜著蛋白質、色素、小分子雜質等物質。對于多糖結構和生物活性的研究,蛋白質的存在會造成干擾。因此,選用對多糖結構和生物活性不造成破壞的脫除方法很重要。常用Sevag法、三氟三氯乙烷法、陰離子交換樹脂法、酶解法等除去蛋白質。Sevag法是脫除蛋白質的經典方法,原理為蛋白質在氯仿等有機溶劑中發生變性。Sevag法的特點為條件溫和、效率低、操作瑣碎和得率不高等。與Sevag法相比,三氟三氯乙烷法脫除蛋白的效率較高,但溶劑易揮發,不宜大規模生產應用。陰離子交換樹脂法可調節溶液pH值使蛋白質形成陰離子,采用陰離子交換樹脂作為流動相,具有去除效果好、成本低、條件溫和等特點。常用活性炭吸附法、乙醇洗滌法、過氧化氫法等除色素。除小分子雜質則主要是用透析法和超濾法。分離除雜后,利用各種多糖的溶解度不同及電荷密度差異進行純化得到單一的多糖物質。常用的純化方法有分級純化法、凝膠柱層析、超濾法等。任益平以藜麥為原料采用超聲波輔助提取、AB-8大孔樹脂吸附、DEAE纖維素分離和Sephadex G-100凝膠柱純化后得到4個組分多糖,再通過傅里葉變換紅外光譜分析、核磁共振波譜分析、剛果紅實驗和微觀結構分析研究分離純化對多糖結構的影響。結果表明,在分離純化的過程中,分子間氫鍵或范德華力受到破壞,分子內部的相互作用隨之減小,而分離純化有利于晶體結構和三螺旋結構的形成。
目前,有較多的文獻報道了關于多糖提取方法對結構影響的研究,但針對分離和純化工藝是否對多糖結構具有影響的研究相對較少。例如,在多糖脫蛋白的研究中,一般以蛋白質清除率、多糖損失率、多糖得率作為指標,并未深入探討脫蛋白方法與多糖結構之間的關系。有學者認為,醋酸鉛法、三氯乙酸法可能對多糖結構造成破壞,Sevag法、醋酸鉛法、三氯乙酸法、氫氧化鈉法可能破壞多糖活性。分級純化法可能會影響多糖結構和活性,金屬絡合物法可能影響多糖活性。但分離純化工藝具體如何影響多糖結構的機理機制,進而影響多糖生物活性還鮮有報道。
3.3 化學修飾
多糖分子的修飾是針對性地利用目標基團替代多糖分子上不同的基團(一般為羥基、羧基、氨基)進行化學修飾,使多糖結構發生變化,單糖構成比例、分子質量和形貌特征等也隨之改變,從而改善多糖生物活性。目前,常用的化學修飾方法主要有硫酸化、磷酸化、羧甲基化、化學降解修飾和乙酰化等。
硫酸化修飾是利用硫酸基團取代羥基生成多糖硫酸化衍生物的過程。常用的方法有濃硫酸法、氯磺酸-吡啶法和三氧化硫-吡啶法。朱麗丹采用氯磺酸-吡啶法,以離子液體為介質對米糠多糖進行硫酸酯化結構修飾,通過傅里葉紅外光譜分析發現,在809.52~836.66 cm處的吸收峰是C-O-S伸縮振動,伸縮振動峰的位置和硫酸根基團在糖環上取代位置有關。氯磺酸-吡啶法對多糖進行硫酸化修飾通常伴隨著多糖的降解。但是,在硫酸化修飾涼粉草多糖過程中,多糖分子質量增加,表明硫酸化衍生物的成功制備沒有伴隨多糖降解,這可能緣于甲酰胺的緩沖作用和硫酸化改性的適度反應條件。Xiao Heng等在利用不同反應條件對海蒿子多糖進行硫酸化修飾后,發現改性導致多糖的單糖組成發生改變,葡萄糖含量顯著降低,而其他單糖(巖藻糖、阿拉伯糖、半乳糖、木糖、甘露糖、半乳糖醛酸和葡萄糖醛酸)含量略微增加。
磷酸化修飾是一種常見的結構修飾方法之一,指多糖中的羥基被磷酸基團取代。常用的磷酸化材料有磷酸、三氯氧磷、磷酸氫二鈉、磷酸二氫鈉和三聚磷酸鈉等。何淼等利用多聚磷酸鹽法修飾山豆根多糖,紅外光譜定性分析發現,1 153.00 cm處的吸收峰為P=O的伸縮振動;1 021.29 cm處為P-O-C的伸縮振動;899.36 cm處為P-O-R的伸縮振動。在三氯氧磷修飾麥冬多糖研究中,張力妮等發現麥冬多糖化學修飾后空間結構發生改變,呈現三螺旋結構,紅外光譜研究發現,在1 035 cm和1 120 cm處有P-O-C的伸縮振動吸收峰。
羧甲基化修飾是將多糖分子上引入羧甲基基團,具有成本低、制備簡便、試劑易得等特點,是多糖結構修飾常用的一種方法。其原理是將多糖與一氯乙酸在堿性的條件下相互反應,從而將羧甲基引入原多糖殘基上的某些羥基上。目前常用的方法有溶媒法和水媒法。吳慧玉采用羧甲基化、硫酸酯化和乙酰化對黃秋葵多糖進行結構修飾,發現經不同方法修飾得到的多糖其分子的不對稱性、鏈構象、分子厚度、分子間距及表面形貌發生改變。
3.4 物理修飾
研究表明,過大的分子質量、復雜的空間結構和高聚合度是阻礙多糖發揮生物活性的主要原因。而對多糖結構采用物理修飾方法,可以達到降低多糖的分子質量、改變多糖空間結構、降低聚合度的目的,有利于增強多糖生物活性。與化學修飾相比,物理修飾具有反應條件溫和、環保、操作簡便等優點。目前,常用的物理修飾方法有超聲波降解修飾、離子輻射修飾和高壓均質降解等。
超聲波降解修飾具有降解過程和降解程度易控制、易獲得目標分子質量、無雜質引入、綠色等優點在多糖結構修飾中得到了廣泛應用,其原理主要是利用低頻(1 MHz)、高強度(3 W/cm)的超聲波通過增加質點振動能量來切斷多糖大分子中的糖苷鍵以降低多糖分子質量,增加水溶性,從而提高一些大分子多糖(如纖維素、殼聚糖、葡聚糖)的生物活性。Yang Haihong等以黑加侖果實為原料,利用超聲波處理降解黑加侖果實多糖,發現多糖分子質量隨超聲時間的延長和功率的增加而減小,使用超聲功率300、500 W和700 W超聲降解100 min后,黏均分子質量從6.12h10Da分別下降到9.40h10、8.00h10、4.65h10Da,數均分子質量也從8.13h10Da分別降至3.39h10、1.74h10、4.57h10Da。
離子輻射修飾具有反應易控、無需添加物、環保、綠色等優點。其原理是采用γ射線、X射線及電子束等電離輻射誘導多糖發生物質聚合、交聯、接枝或降解等物理化學變化以改變多糖的分子結構提高其生物活性。目前,離子輻射修飾在殼聚糖、海藻酸鈉、纖維素等多糖降解上都有報道。Henniges等利用γ射線照射木屑后,其纖維素的聚合度發生了降低。
高壓均質降解利用高壓均質機來實現多糖降解的一種新型純物理降解方法,具有純物理作用、產品純度高、簡便、環保等優點。高壓均質降解技術在多糖中的應用,目前報道較多的是針對殼聚糖的降解。研究發現,高壓均質降解殼聚糖是通過機械作用切斷殼聚糖分子鏈,降低分子質量,降解效果隨循環次數的增加而增加,對產物的結構和脫乙酰度基本不改變。Xie Fan等采用高壓均質處理(200 MPa、20 min)對紫肉馬鈴薯多糖進行改性,結果發現其單糖組成發生改變,阿拉伯糖的含量升高,而葡萄糖的含量則降低。Avi等為了研究高壓均質化對果膠的影響,以柑橘果膠和蘋果果膠為原料進行高壓均質化處理,結果表明,高壓均質可以使柑橘果膠發生明顯解聚,顯著降低其分子質量,但對蘋果果膠幾乎沒有影響,這可能與蘋果果膠中性糖含量更加豐富、結構更加致密有關。
改變各種植物多糖結構的方法整理見表2。

表2 改變植物多糖結構的主要方法Table 2 Main chemical and physical methods to modify the structure of plant polysaccharides
4 結 語
自然界植物種類繁多,植物多糖結構豐富。植物多糖作為腸道菌群的能量來源,可作為潛在益生元開發差異化的健康產品,具有廣闊的應用前景。植物多糖結構復雜,目前構效關系尚未完全明晰,制約了植物多糖精準化利用。隨著多糖結構解析的深入,相關研究人員應結合多組學技術,逐漸明確腸道微生物對植物多糖的利用過程;建立具有益生活性的植物多糖快速篩選技術;同時應關注植物多糖與蛋白、脂肪、維生素以及其他功能性成分如何協同調節腸道菌群、促進人體健康。
猜你喜歡
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
