氮肥類型對不同稻作模式水稻生長和產量的影響
2022-07-07 01:11:32姜碩琛張海維吳啟俠朱建強
中國土壤與肥料 2022年5期
姜碩琛,張海維,孔 盼,杜 斌,吳啟俠,朱建強
(長江大學農學院,濕地生態與農業利用教育部工程研究中心,湖北 荊州 434025)
水稻是全球重要的糧食作物之一,是我國2/3人口的主食[1]。預計到2025年,水稻產量需增加目前水平的60%,相當于每年至少增加1.2%的單產,才能滿足全球人口增長的需求[2]。在過去的半個世紀,雖然我國水稻產量急劇增加,但伴隨的是肥料的大量施用,尤其是氮肥[3]。不合理的氮肥施用往往會降低氮肥利用率[4],未被水稻利用的氮會通過地表徑流、滲漏以及氨揮發的形式而損失,引起環境污染[5]。同時低的氮肥利用率不利于水稻生長和產量增長[6]。因此,探究高效的氮肥施用方式,有助于提高水稻產量和保護生態環境。稻蝦共生模式是一種以傳統水稻種植為基礎,將水稻種植與小龍蝦養殖在稻田中相結合的新技術,是一種高效的稻田生態種養模式[7]。與傳統稻田模式相比,稻蝦共生模式增加了農民收益。對于水稻產量較低的低產田,由于收益過低,農戶不愿種植水稻,導致農田被擱置、荒廢,而將低產田改為稻蝦種養田后,由于小龍蝦產品增加了經濟效益,因此提高了農戶種糧積極性,保證了糧食的種植面積,該模式 同時滿足了農民的“錢袋子”和我國的“糧袋子”[8]。 為進一步增加稻蝦共生模式中水稻的產量,提高經濟效益和生態效益,探究高效的氮肥施用方式十分重要。
復合肥作基肥、尿素作追肥的氮肥管理方式是我國應用最早,且應用時間最長的施氮方法,雖然該方法養分高、增產快,但長期施用會導致土壤質量退化、農業面源污染等問題[9]。為保持土壤肥力和減少環境污染,有機肥和無機肥配合施用或使用緩/控釋肥,是滿足農業可持續發展和環境友好型的施氮方式。陸海飛等[10]研究發現,長期采用有機肥和無機肥配施可提高土壤細菌多樣性和酶活性,進而提高農田生態系統的生產力。劉紅江 等[11]研究發現,有機肥和無機肥配施時,增加有機肥的施用比例,可以降低地表徑流中總氮和鉀的流失量,有機肥和無機肥施用比例為5∶5時,可以達到保證產量并降低農田養分流失的效果。Zhang 等[12]研究表明,有機肥與無機肥配施可以增加土壤有機質含量、提高籽粒氮含量和水稻產量,雖然地表水NH4+含量和氨揮發有所增長,但由于其提高了土壤中氮的殘留量,從而減少了氮素的損失。魏海燕等[13]研究發現,對于小穗型水稻,一次性基施緩/控施肥可提高水稻產量,但對于大穗型水稻,需要基施緩/控釋肥后,再追施尿素才能滿足水稻的氮肥需求。Wei等[14]研究表明用脲甲醛樹脂或聚合物包膜尿素作基肥,尿素作分蘗肥,可以提高水稻產量與稻米品質。Ding等[15]通過Meta分析比較了中國境內緩/控釋肥和有機肥等肥料對水稻產量的影響,結果顯示有機肥增產7.8%,緩/控釋肥增產7.4%。
雖然前人圍繞復合肥、有機無機互混肥和緩控釋肥對水稻產量的影響,已有較多研究,但針對稻蝦共生模式下,不同氮肥類型對水稻生長和產量影響的報道仍較少。在稻蝦共生和傳統中稻模式中,不同氮肥施用類型對水稻生長和產量影響的差異比較更是未曾報道。本研究選擇復合肥、緩/控釋肥、有機無機互混肥3種氮肥類型,以不施用氮肥作為空白對照,研究在常規中稻和稻蝦共生模式中,不同氮肥類型對水稻生長動態、光合特性、產量和氮肥偏生產力的影響,并為不同稻作模式下優化氮肥管理提供依據。
1 材料與方法
1.1 試驗材料
本試驗于2020年在湖北省荊州市長江大學農業科技示范基地(30°21′N,112°09′E)進行,該地屬北亞熱帶農業氣候帶,年平均氣溫16.5℃,≥10℃積溫5094.9~5204.3℃,年平均降水量1095 mm,年平均日照時數1718 h。中稻模式種植制度為水稻、冬小麥輪作,土壤類型為粘壤土,土壤(0~20 cm)有機質21.4 g/kg、全氮2.24 g/kg、全磷0.54 g/kg、全鉀3.67 g/kg、堿解氮89.6 mg/kg、 有效磷51.3 mg/kg、速效鉀114.6 mg/kg、pH值 6.8。稻蝦模式所用試驗田已進行了3年的稻蝦綜合種養管理,種植制度為稻蝦共生、冬閑田,土壤類型為潛育型水稻土,土壤(0~20 cm)有機質25.4 g/kg、全氮1.91 g/kg、全磷0.34 g/kg、 全鉀3.34 g/kg、堿解氮69.4 mg/kg、有效磷24.1 mg/kg、 速效鉀118.7 mg/kg、pH值7.3。緩/控釋肥(N∶P2O5∶ K2O=28∶15∶5),有機無機互混肥(有機肥∶N∶ P2O5∶K2O=15∶18∶10∶12),復合肥(N∶P2O5∶K2O= 15∶15∶15)由中國中化集團有限公司提供。供試品種為黃華占,由長江大學農學院提供。
1.2 試驗設計
試驗采取裂區設計,主區為2個稻作模式,分別為常規中稻模式和稻蝦共生模式。兩種稻作模式的差異如下。(1)水管理差異,常規中稻模式水管理方式:移栽至分蘗末期保持田間3~5 cm水層,分蘗末期排水曬田控制無效分蘗,曬田結束至成熟前7 d保持2~4 cm水層,成熟前7 d排水曬田便于收獲。稻蝦共生模式水管理方式為:移栽至分蘗末期保持田間3~5 cm水層,分蘗末期排水曬田控制無效分蘗,曬田結束至成熟期前7 d保持25~35 cm水層以便小龍蝦活動,成熟前7 d排水曬田便于收獲。(2)小龍蝦養殖的差異,稻蝦共生模式下,移栽至曬田結束期間,小龍蝦一直生活在蝦溝中,田面四周設有高20 cm、寬20 cm的壟,防止小龍蝦進田破壞分蘗期的水稻。曬田結束復水后,沿田邊四周撒小龍蝦飼料40 g/m2,每天 17:00撒1次,持續7 d,引導小龍蝦進田,提高稻蝦共生度,此時部分小龍蝦生活在田間,7 d后通過地籠捕撈法(地籠規格為:網眼8 mm,框架12 cm×14 cm,長1.4 m,田間地籠密度為1個/50 m2)測得共生度為(0.23±0.04)只/m2。裂區為氮肥類型差異,設有不施氮肥(CK)、緩/控釋肥、復合肥、有機無機互混肥,3個施氮處理的氮肥用量均為N 120 kg/hm2,4個處理的P2O5和K2O用量分別為62.5和135 kg/hm2。具體施用方法為:緩/ 控釋肥中氮肥按照基肥∶蘗肥∶穗肥為8∶0∶2分施,復合肥和有機無機互混肥中氮肥按照基肥∶蘗肥∶穗 肥為5∶3∶2分施。3個施氮處理中基肥按照不同類型氮肥施用,蘗肥和穗肥通過尿素的形式施用。磷肥全部做基肥,鉀肥按照基肥∶穗肥為5∶5分施,4個處理中需補充的P2O5由磷酸氫二鈉代替,K2O由氯化鉀代替。具體試驗設計見表1、表2。 兩個稻作模式和4個氮肥類型共組成8個處理,每個處理重復3次,共24個小區,小區面積為300 m2。 田間試驗采取育秧移栽的方式,5月1日播種,6月1日移栽,株行距為40 cm×20 cm,每穴種植2株。其他田間管理按照當地農民高產管理,并且及時防治病蟲草害,避免產量損失。
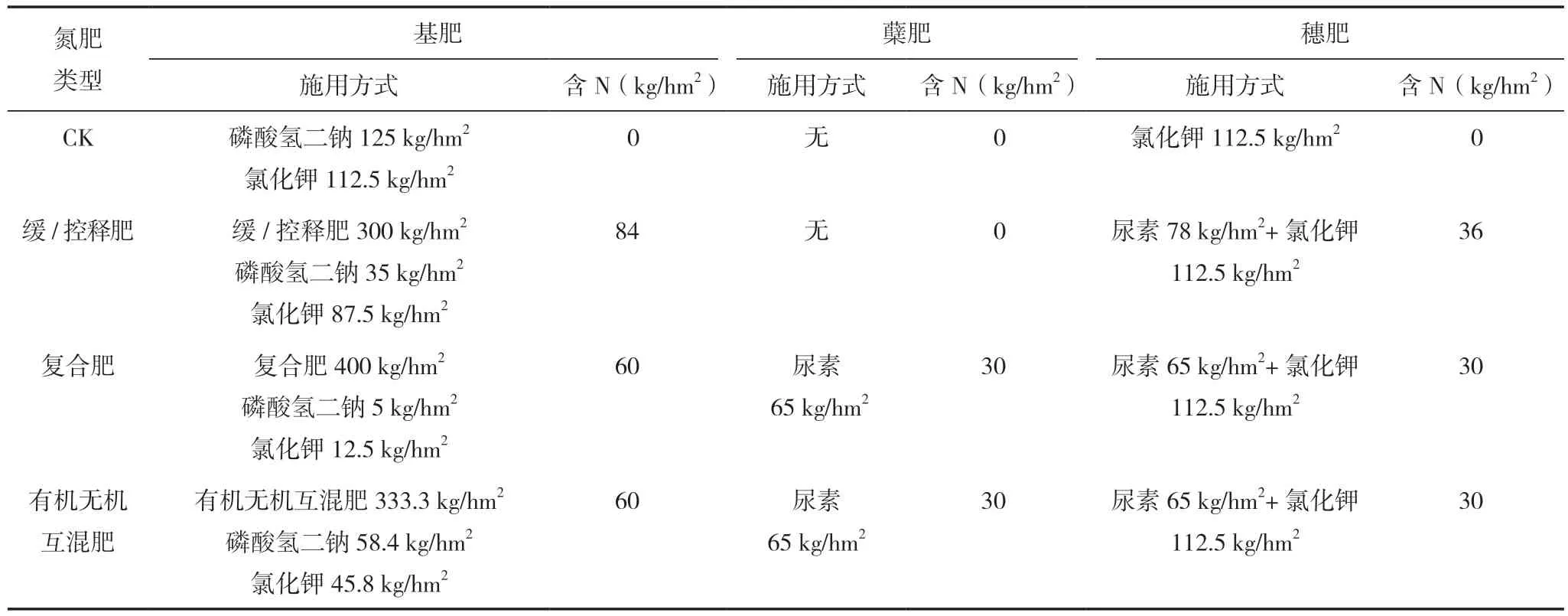
表1 試驗施肥設計
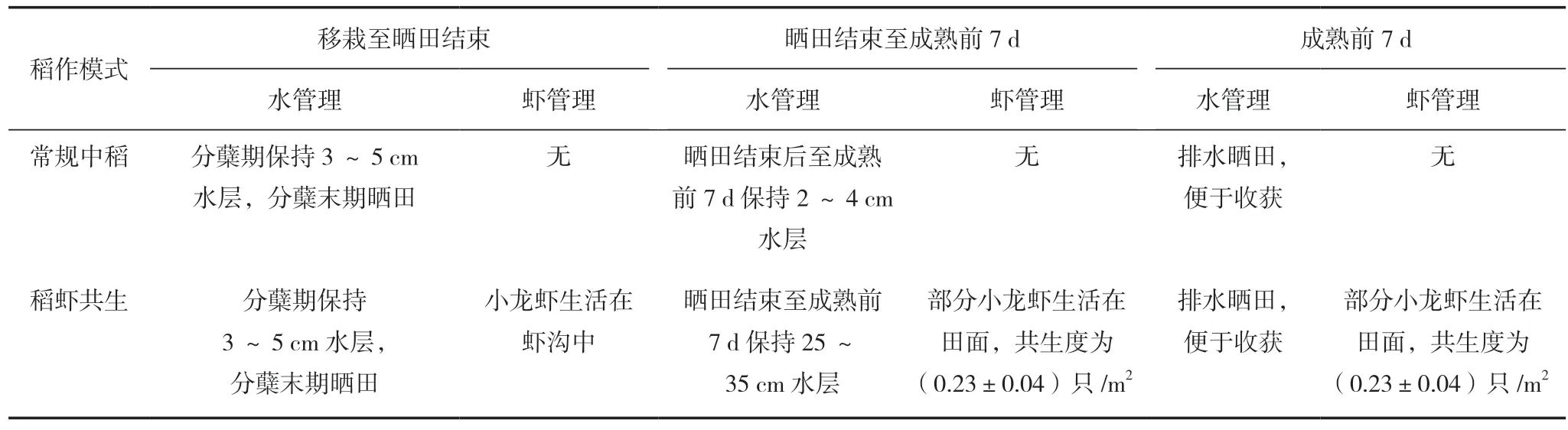
表2 試驗稻作模式設計
1.3 樣品采集與測定
1.3.1 生育期調查
記錄水稻移栽期、分蘗盛期、抽穗期(田間50%的植株抽穗)和成熟期。
1.3.2 干物質積累量和葉面積的測定
于水稻分蘗盛期、抽穗期和成熟期,每小區調查100穴植株的莖蘗數,計算單穴平均莖蘗數,據此每小區取代表性植株5穴,測量葉片的長與寬,按公式計算葉面積=長×寬×0.75,并計算葉面積指數(LAI)。將5穴水稻地上部分分成莖葉和穗(水稻分蘗盛期僅分為莖葉),在105℃下殺青30 min,80℃烘干至恒重,測定各部分干物質重。
1.3.3 光合勢
光合勢(LAD)=1/2×(L1+L2)×(t2-t1)。式中,L1和L2為前后2次測定的葉面積,t1和t2為前后2次測定的間隔時間(d),LAD單位為m2/(d·hm2)。
1.3.4 群體生長率
群體生長率(CGR)=(W2-W1)/[A(t2-t1)]。式中,W1和W2為前后2次測定的干物質積累量,t1和t2為前后2次測定的間隔時間(d),A為土地面積,CGR單位為g/(d·m2)。
1.3.5 SPAD值
于晴天9:00~11:30,用SPAD-502葉綠素計測定葉片SPAD值。在分蘗盛期和抽穗期,每個小區根據單穴平均莖蘗數選擇5穴水稻,測定時選擇水稻劍葉(分蘗盛期選擇頂部全展葉)測定1/2處及其上下3 cm、葉寬1/4或3/4的位置,每穴水稻重復測定3次,3次平均值作為該穴水稻SPAD值。
1.3.6 凈光合速率
于晴天9:00~11:30,用LI-6400便攜式光合作用測定系統測定葉片凈光合速率。在分蘗盛期和抽穗期,每個小區根據單穴平均莖蘗數選擇5穴水稻,測定時選擇水稻劍葉(分蘗盛期選擇頂部全展葉)的中部,每葉片重復測定3次,3次平均值作為該穴水稻的凈光合速率。
1.3.7 產量及其構成和氮素偏生產力
在水稻成熟期,每個小區根據單穴平均莖蘗數選擇5穴水稻,測定每穗穎花數,用水漂法區分飽粒(沉入水底者)和空癟粒,計算飽粒結實率和飽粒千粒重和收獲指數。每個小區選擇40 m2水稻測定實際產量,稻蝦共生模式根據種植水稻的有效面積進行實量,將稻谷烘干后折算到含水率為14%的千粒重和產量。氮肥偏生產力(kg/kg)=施氮區產量/氮肥施用量。
1.4 數據分析
數據通過Excel 2010進行預整理,通過SPSS 21.0進行數據分析,使用方差分析檢驗不同處理間的差異(P<0.05),用鄧肯的多區間法進行多重 比較。
2 結果與分析
2.1 稻作模式和氮肥類型對水稻生育進程的影響
由表3可見,常規中稻模式水稻全生育期(113~122 d)短于稻蝦共生模式(129~140 d)。常規中稻模式下,未施氮處理的全生育期比施氮處理少7~9 d,其中移栽期至抽穗期比施氮處理多2~3 d,抽穗期至成熟期比施氮處理少10~12 d。稻蝦共生模式下,未施氮處理的水稻全生育期比施氮處理多8~11 d,其中,移栽期至抽穗期天數比施氮處理多6~8 d,抽穗期至成熟期天數比施氮處理多1~3 d。3個施氮處理在各生育期的天數差異小于2 d。由此可得:稻蝦共生模式的生育期長于常規中稻模式,常規中稻模式下未施氮處理的生殖生長期和全生育期短于3個施氮處理,稻蝦共生模式下未施氮處理的營養生長期和全生育期長于3個施氮處理。
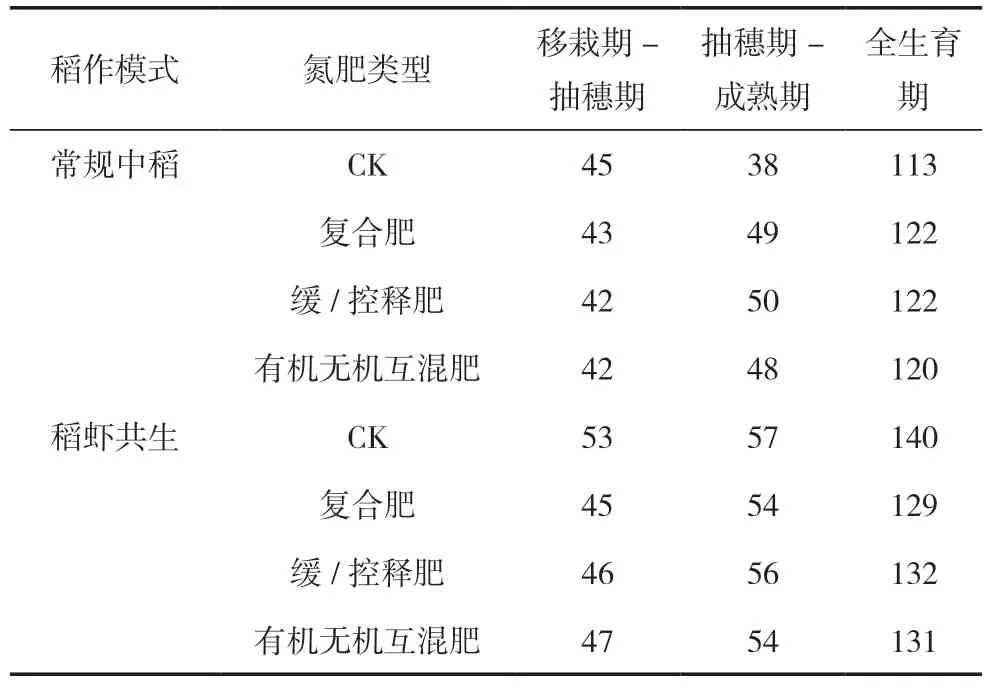
表3 不同處理間水稻的生育進程(d)
2.2 稻作模式和氮肥類型對水稻各時期葉面積指數和光合勢的影響
由表4可知,稻作模式對抽穗期LAI有顯著影響,對抽穗期至成熟期LAD有極顯著影響,氮肥類型和稻作模式交互作用對抽穗期和成熟期LAI、分蘗期至成熟期LAD有極顯著影響。傳統中稻模式抽穗期LAI比稻蝦共生模式高5.67%,抽穗期至成熟期LAD比稻蝦模式低5.46%。有機無機互混肥的抽穗期和成熟期的LAI、分蘗期至抽穗期和抽穗期至成熟期的LAD最高,分別比CK高88.00% 和73.75%、54.88%和91.63%,分別比復合肥高9.30% 和3.58%、8.34%和6.04%,分別比緩/控釋肥高5.62%和3.27%、7.01%和10.88%。傳統中稻模式下,以上4個指標在3種氮肥類型間無顯著差異,但施氮處理顯著高于未施氮處理;稻蝦共生模式下,以上4個指標施氮處理顯著高于未施氮處理,此外,抽穗期LAI和分蘗期至抽穗期LAD以有機無機互混肥最高,抽穗期至成熟期LAD以復合肥和有機無機互混肥較高。以上結果表明:傳統中稻模式LAI高于稻蝦共生模式,但LAD反之。有機無機互混肥的LAI和LAD最高,且在稻蝦共生模式中更加明顯。
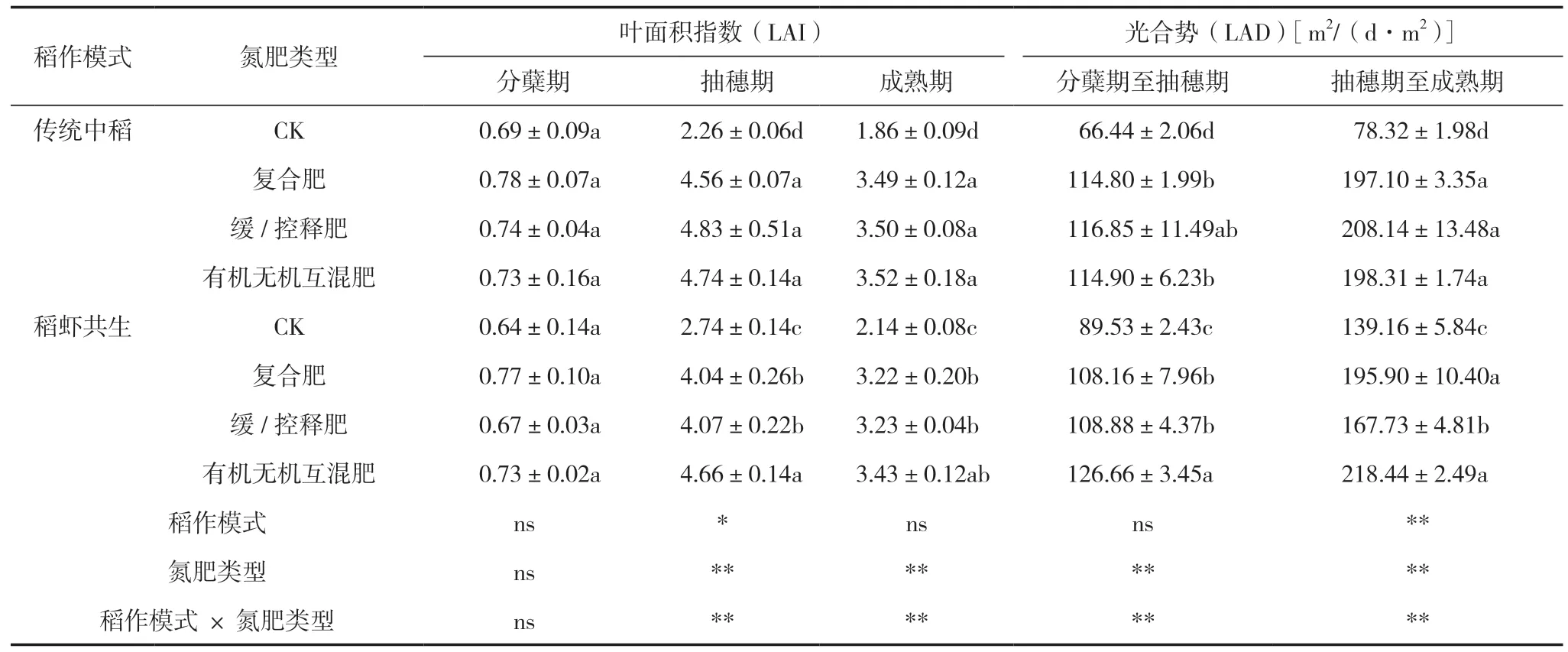
表4 不同處理間水稻各時期的葉面積指數和光合勢
2.3 稻作模式和氮肥類型對水稻各時期干物質積累量和生長速率的影響
由表5可知,稻作模式對抽穗期和成熟期干物質積累量(DW)、分蘗期至抽穗期生長速率有極顯著影響,對抽穗期至成熟期生長速率有顯著影響,氮肥類型對抽穗期和成熟期DW、分蘗期至抽穗期和抽穗期至成熟期生長速率有極顯著影響,稻作模式和氮肥類型的交互作用對抽穗期DW有極顯著影響,對分蘗期至抽穗期生長速率有顯著影響。傳統中稻模式的抽穗期和成熟期DW、分蘗期至抽穗期和抽穗期至成熟期生長速率分別比稻蝦共生模式高7.89%和8.54%、21.58%和21.30%。緩/控釋肥的抽穗期和成熟期DW、分蘗期至抽穗期的生長速率最高,分別比CK高91.73%和102.93%、 163.23%,分別比復合肥高15.13%和6.00%、21.79%,分別比緩/控釋肥高8.55%和3.56%、11.48%。復合肥的抽穗期至成熟期生長速率最高,兩種稻作模式數值平均后,比CK、緩/控釋肥和有機無機互混肥分別高127.79%、6.95%和3.41%。傳統中稻模式和稻蝦共生模式下,抽穗期DW和分蘗期至抽穗期生長速率大小順序均為緩/控釋肥>有機無機互混肥>復合肥>CK,傳統中稻模式成熟期DW大小順序為緩/控釋肥>有機無機互混肥>復合 肥>CK,稻蝦共生模式下成熟期DW大小順序為 緩/控釋肥>復合肥>有機無機互混肥>CK。綜上可見,傳統中稻模式DW和生長速率高于稻蝦共生模式,緩/控釋肥DW和分蘗期至抽穗期生長速率最高,復合肥抽穗期至成熟期生長速率最高。
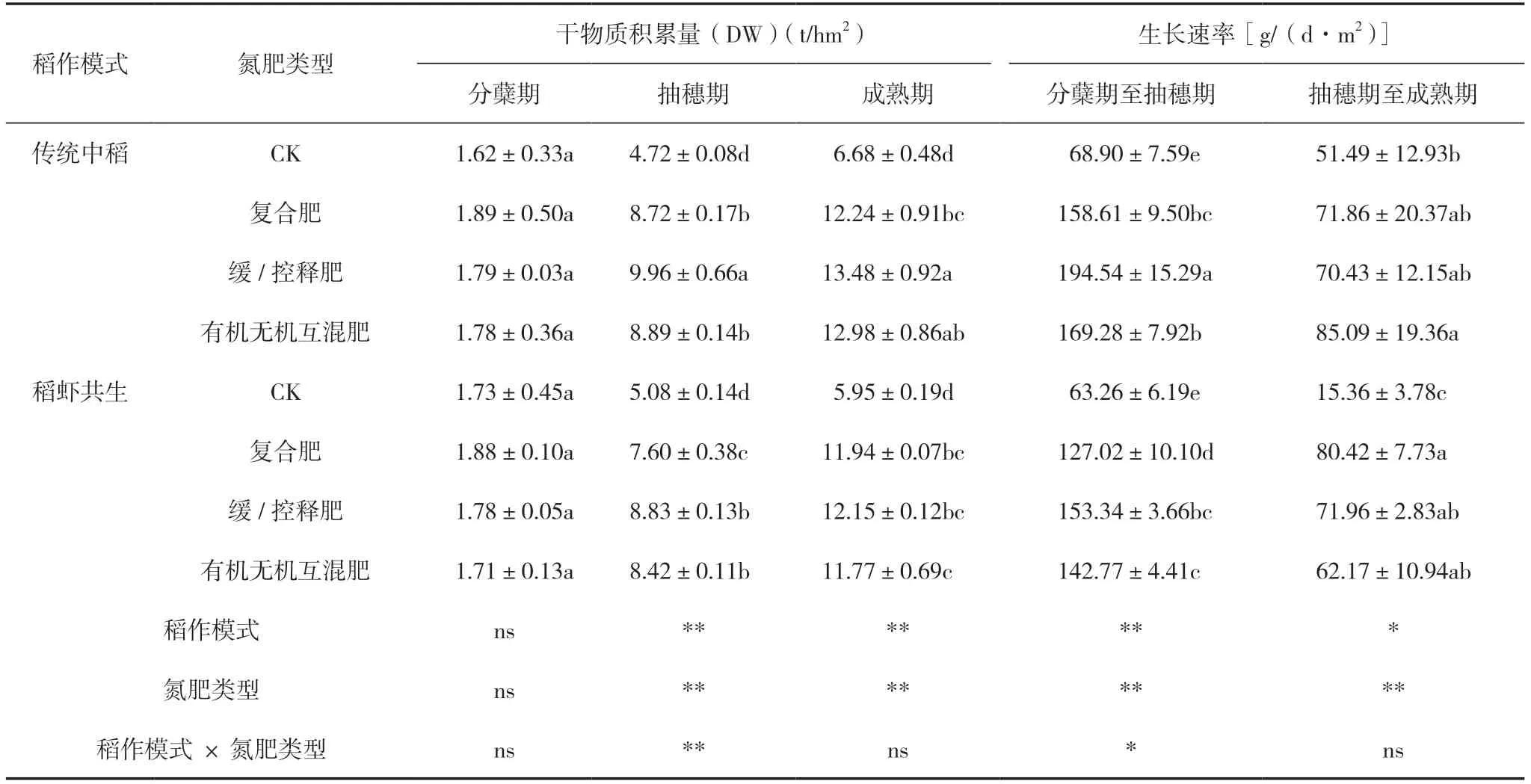
表5 不同處理間水稻各時期的干物質積累量和生長速率
2.4 稻作模式和氮肥類型對水稻各時期SPAD值和光合速率的影響
由表6可知,稻作模式對抽穗期SPAD值和光合速率有極顯著影響,氮肥類型對抽穗期SPAD值以及分蘗期和抽穗期光合速率有極顯著影響。傳統中稻模式抽穗期SPAD值和光合速率分別比稻蝦共生模式高5.19%和7.93%。有機無機互混肥的抽穗期SPAD值和光合速率最高,分別比CK高22.27%和21.21%,分別比復合肥高0.43%和1.73%,分別比緩/控釋肥高10.43%和6.83%。復合肥的分蘗期光合速率最高,比CK、緩/控釋肥和有機無機互混肥分別高17.74%、8.53%和1.68%。以上結果顯示,傳統中稻模式的SPAD值和光合速率高于稻蝦共生模式,有機無機互混肥抽穗期光合能力更強,而復合肥分蘗期光合能力更強。
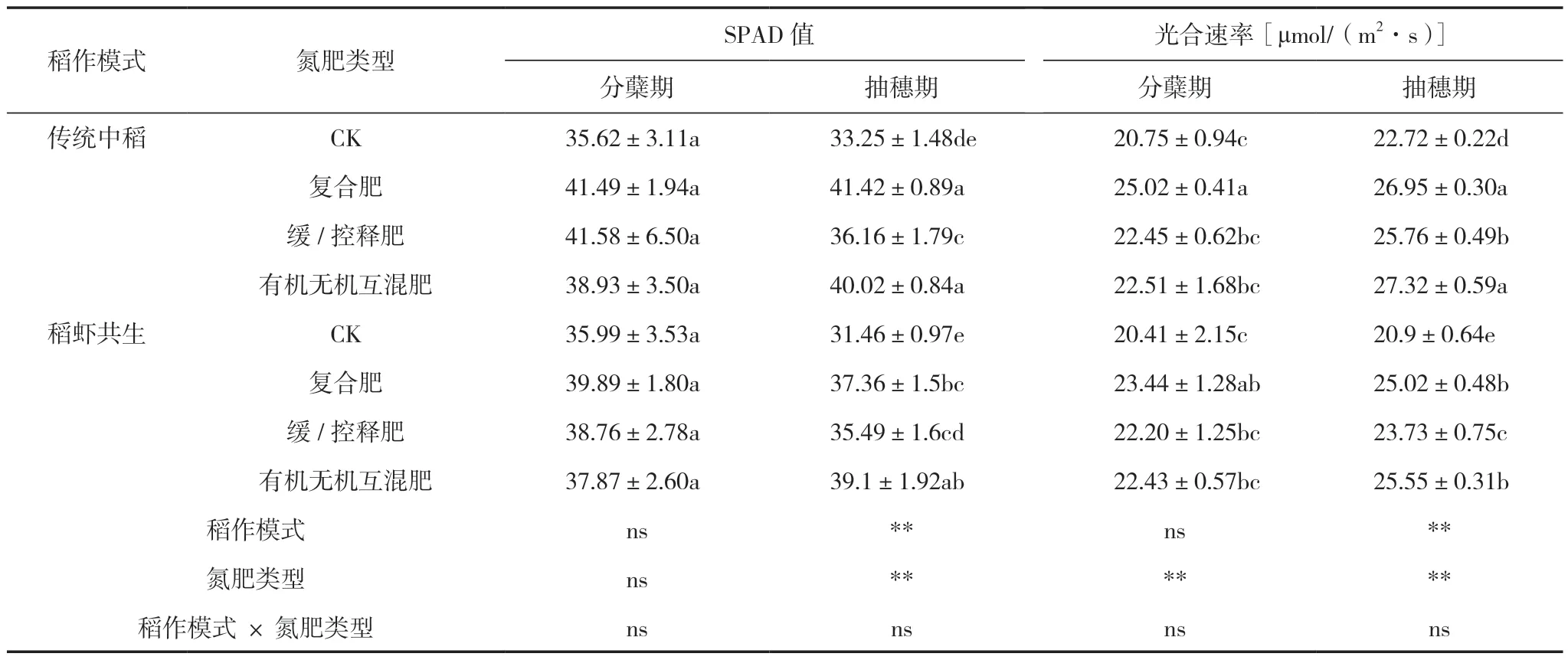
表6 不同處理間水稻各時期的SPAD值和光合速率
2.5 稻作模式和氮肥類型對水稻產量、產量構成因子和氮肥偏生產力的影響
由表7可知,稻作模式對產量有極顯著影響,氮肥類型對產量、單位面積有效穗數、千粒重和氮肥偏生產力有極顯著影響。傳統中稻模式的產量比稻蝦共生模式產量高14.06%。緩/控釋肥的產量和單位面積有效穗數最高,分別比CK高99.96%和99.46%,分別比復合肥高7.96%和4.95%,分別比有機無機互混肥高6.41%和20.03%,緩/控釋肥的氮肥偏生產力最高,比復合肥和有機無機互混肥分別高17.29%和13.70%。有機無機互混肥的千粒重最高,分別比CK、復合肥和緩/控釋肥高6.82%、6.01%和5.31%。綜上所述,傳統中稻模式產量高于稻蝦共生模式,緩/控釋肥產量、單位面積有效穗數和氮肥偏生產力最高,有機無機互混肥千粒重最高。

表7 不同處理間水稻的產量、產量構成因子和氮肥偏生產力
3 討論
氮肥類型顯著影響水稻產量和氮肥利用率[16],稻蝦共生模式是在傳統中稻模式的基礎上引入小龍蝦養殖的一種新技術,具有提高經濟效益和生態效益的廣泛前景[8],研究不同氮肥類型對稻蝦共生模式和傳統中稻模式下水稻生長和產量的影響,具有較高的研究意義。本研究中,稻蝦共生模式的水稻生育期長于常規中稻模式(表3),在水稻分蘗結束至成熟期,稻蝦共生模式的田間水位(25~35 cm)高于傳統中稻模式(2~4 cm),長期處于深水環境引起的澇漬會限制水稻根系發育和根系呼吸作用,影響養分的吸收和碳水化合物的合成,從而延長水稻生育期[17],同時稻蝦共生模式田間有小龍蝦的活動,小龍蝦的排泄物不斷為水稻追肥,導致水稻貪青遲熟[18]。稻蝦共生模式下未施氮處理生育期更長,這與較高的非結構碳水化合物儲備可縮短澇漬引起的延緩生長有關[19],通過水稻生長的各項指標(表4~6)可以看出,未施氮處理下水稻營養結構相對較差,因此在澇漬逆境下生長更慢,生育期長于施氮處理。常規中稻模式下,未施氮處理的生殖生長期和全生育期短于3個施氮處理,這與Wei等[20]結論一致,由于未施氮處理導致土壤缺乏氮元素,使水稻葉綠素含量和光合速率降低,引起水稻早衰[21]。值得注意的是,同樣未施氮肥,稻蝦共生模式下生育期延長了27 d,其中營養生長期延長了8 d,灌漿期延長了19 d。缺氮使生殖生長提前到來[22],而稻蝦共生模式下水體和土壤中氮含量始終高于傳統中稻模式,同時澇漬延緩了水稻生長[17],二者是未施氮處理在稻蝦共生模式中營養生長期延長的主要原因。傳統中稻模式下氮素缺乏引起水稻早衰,縮短了灌漿時間;另一方面稻蝦共生模式水體和土壤氮含量較高,雖然氮含量與灌漿速率呈正相關[20],但未施氮處理始終維持低氮供應,使水稻不會缺氮早衰[21],也不會高氮而快速灌 漿[23],導致灌漿時間被延長,但目前沒有相關報道證明持續供應低氮會延長灌漿時間,需要試驗進一步證明。此外,大量研究證明干濕交替會提高灌漿效 率[24-26],與稻蝦共生模式相比,傳統中稻模式相對存在干濕交替的效果。未施氮處理的水稻在傳統中稻模式下灌漿期縮短,而稻蝦共生模式下灌漿期延長,導致二者灌漿時間相差19 d。
本研究發現,傳統中稻模式LAI、DW、生長速率、SPAD和光合速率高于稻蝦共生模式(表4~6),可能原因是稻蝦共生模式的長期深水灌溉,限制了水稻根系發育和根系呼吸作用,降低了水稻的光合作用,減少了碳水化合物的合成,限制了營養結構的搭建[17,27-29],而LAD反之,這是因為稻蝦共生模式生育期長于傳統中稻模式(表3)。有機無機互混肥LAI最高(表4),說明有機無機互混肥可促進葉生長,提高冠層覆蓋面積,這與Bi等[30]和Wang等[31]的結論一致,在稻蝦模式中更明顯,原因可能是有機肥作為水生生物的食物之一,有機肥的使用豐富了水生生物食物來源,同時水生生物是小龍蝦的食物,可促進小龍蝦生長,水生生物和小龍蝦的糞便是水稻營養的另一個來 源[32],促進水稻的生長。緩/控釋肥DW在分蘗期至抽穗期合成速率最高(表5),這與Hou等[33]的研究結論一致,說明緩/控釋肥的養分釋放速度與水稻生長所需營養物質速度更同步。復合肥抽穗期至成熟期生長速率最高(表5),可能是復合肥的養分釋放過快,使水稻抽穗期地上部分干物質積累較低,抽穗期再次施肥后,抽穗期至成熟期水稻養分吸收和干物質積累更高。有機無機互混肥抽穗期光合能力更強(表6),原因可能是有機肥提高了水稻葉綠素含量和光合能力[34],這與侯紅乾 等[34]研究結論一致。復合肥分蘗期光合能力更強,這與復合肥的養分釋放速度最快有關。
我國對稻蝦共生模式的研究結果普遍表明,稻蝦共生模式改善了土壤結構,增加了土壤養分、微生物的活性以及群落功能多樣性,提高了水稻產 量[5,35-36]。本研究中,傳統中稻模式產量高于稻蝦共生模式(表7),這與前人結論相反,蔡晨 等[35]和佀國涵等[36]的研究沒有提及稻蝦共生模式提高土壤肥力后對水稻產量的影響,佀國涵 等[36]研究采用稻蝦共作的方式提高了水稻產量,但稻蝦共作模式是中稻收獲后灌水養蝦,水稻并未受到深水灌溉的澇漬脅迫。本研究通過調查湖北省多地農戶在稻蝦綜合種養模式中的日常管理后,選擇農戶最常用的水、蝦管理方式進行試驗,長期的深水灌溉應該是稻蝦共生模式水稻生長受限、產量降低的主要原因。因此,本文建議稻蝦共生模式在稻蝦共生期,田間水位不宜超過25 cm。緩/控釋肥產量、單位面積有效穗數和氮肥偏生產力最高,說明緩/控釋肥是提高水稻產量和氮肥偏生產力最優的氮肥類型。有機無機互混肥千粒重最高,這與有機無機互混肥增加了灌漿期光合作用有關 (表6)。
4 結論
與常規中稻相比,稻蝦共生模式延長了生育期,降低了LAI、DW、生長速率、SPAD值、光合速率和產量,綜合試驗結果,為了提高水稻產量,改善水稻品質,應降低稻蝦共生期田間水位,緩解長期深水灌溉引起的澇漬脅迫,促進水稻生長。有機無機互混肥的LAI、營養生長期光和能力和千粒重最高,復合肥的分蘗期光合能力和生殖生長期生長速率最高,緩/控釋肥的干物質積累量、分蘗期至抽穗期生長速率、產量和氮肥偏生產力最高。說明緩/控釋肥是適用于傳統中稻模式和稻蝦共生模式的最優氮肥類型。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年14期)2021-10-14 08:35:40
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
