沙化和人工植被重建對高寒草地土壤細菌群落特征的影響
2022-07-08 06:24:38王亞妮胡宜剛王增如李以康張振華周華坤
草業學報 2022年5期
關鍵詞:結構
王亞妮 ,胡宜剛 ,王增如 ,李以康 ,張振華 ,周華坤
(1. 中國科學院西北生態環境資源研究院沙坡頭沙漠研究試驗站,甘肅 蘭州 730000;2. 中國科學院西北高原生物研究所,青海 西寧 810008;3. 中國科學院大學,北京100049)
高寒草地是青藏高原的典型生態系統類型之一,在畜牧業生產、水源涵養、生物多樣性保護、水土保持、生態系統“碳匯”等諸多方面發揮著不可替代的生態和生產功能[1]。由于青藏高原海拔高且氣候寒冷的自然地理環境,高寒草地的生態系統脆弱性和敏感性特點非常突出[2]。近幾十年來,由于全球氣候變暖、過度放牧、開墾等人類活動的多重作用,青藏高原高寒草地出現了以植物物種減少、植被覆蓋度下降、初級生產力降低、優質牧草比例下降、土壤肥力衰減等為主要特征的不同程度的草地退化,局部地區甚至出現草地完全沙化、“黑土型”次生裸地等極端退化現象[3-4]。這種草地退化也引發了草地生產力降低、水土流失加劇、生物多樣性喪失等一系列生態環境問題,嚴重影響了當地的社會、經濟和生態環境的可持續發展乃至青藏高原生態安全屏障作用的發揮[1]。
近年來,如何對青藏高原退化的高寒草地進行有效治理和恢復備受人們的關注。采用圍欄封育、劃區輪牧、補播、施肥等措施可對輕度退化的草地進行植被恢復,而針對完全沙化的草地和“黑土型”次生裸地,合理又科學地進行人工植被重建是其必經之路[5-6]。在多種措施綜合應用的情況下,評價不同措施的恢復效果是指導人們科學地開展退化草地的恢復與治理工作的關鍵。目前,一些學者從土壤肥力特征、植被功能屬性等方面研究了退牧還草、草地圍封、禁牧、補播、施肥[7]、適生樹種和優良牧草間種[8]、多年生禾本科牧草混播人工種草[9-10]等不同恢復措施對高寒草地的影響及其恢復效果。研究發現,這些措施不同程度地促進了退化草地的植被群落結構、植物生產力和土壤養分的恢復,但不同措施的恢復效果存在較大差異[11]。然而,由于大多數的相關研究主要關注土壤肥力、草地生產力等方面,往往忽視了土壤微生物群落結構的恢復,其評價指標體系并不完整,因此,也影響了評價結果的準確性。
土壤微生物通過參與生物固氮、凋落物分解、養分循環和周轉等多個生態過程直接或間接地影響著草地生態系統的結構和功能[12-13]。生物量、多樣性和群落結構是度量土壤微生物群落特征的主要指標。由于土壤微生物對環境因子的變化非常敏感,其生物量、多樣性和群落結構如何響應環境因子的變化及其機理成為土壤微生物生態學研究的重要方向。細菌是土壤微生物的主要群落之一,在土壤微生物中的占比較高,部分地區甚至在82%以上[14]。許多研究發現,土壤細菌群落生物量、多樣性及其結構與土壤pH、含水量、有機碳、全氮、植被類型等因子密切相關[15-17]。草地沙化顯著降低了土壤含水量、養分水平和植物多樣性[18-19],相反,人工植被重建則有效地促進了植物的定居與繁殖、土壤物理特性和養分水平的提高和改善[20]。因此,這些因素的改變勢必會促使土壤細菌群落朝著不同方向演替。然而,目前對高寒草地土壤細菌群落演變的研究主要集中在單方面的退化或恢復過程,缺少對沙化和人工植被重建這兩個相反生態過程的對比研究,對土壤細菌群落如何響應沙化和植被重建及其機理尚不清楚。況且,有關利用草本物種和灌木物種進行人工植被重建如何影響土壤細菌群落結構的研究幾乎未見報道,從而導致人工植被重建對于植物物種選擇的科學依據不夠充分。
本研究采用qPCR 和Illumina MiSeq 高通量測序技術研究高寒草地沙化和人工植被重建后土壤細菌群落生物量、多樣性及其結構的變化特征。結合植被群落和土壤屬性的變化,剖析驅動高寒草地退化和恢復過程中土壤細菌群落的關鍵因素及其貢獻量,旨在從土壤微生物的角度為高寒退化草地的人工植被重建與生態恢復提供科學依據。
1 材料與方法
1.1 研究區概況
研究區位于青海省貴南縣過馬營鎮(101°6′2″E,35°48′49″N),平均海拔3107 m。該區域屬于高原大陸性氣候,全年干旱少雨,氣候干燥而寒冷。氣溫年較差小而日較差大,年平均氣溫2.3 ℃,最高氣溫29.3 ℃,最低氣溫-29.2 ℃,年均降水量391 mm,多集中在6-9月,年蒸發量在1300 mm 以上,年均日照時數為2703 h。自然植被類型為高寒草原,主要以早熟禾(Poa annua)、矮嵩草(Kobresia humilis)、叢生黃耆(Astragalus confertus)、多色苦荬(Ixeris chinensissubsp.versicolor)為優勢種,局部地區地表部分沙化或完全沙化。近30年來,在沙化草地通過播種草本植物早熟禾和叢生黃耆或栽植灌木植物檸條(Caragana korshinskii)進行了人工植被重建。
1.2 試驗樣地與植被群落調查
選擇青海省貴南縣過馬營鎮沙溝小流域,位于同一坡面相同海拔的天然草地(natural grassland,NG)、沙化草地(desertified grassland,DG)、草本人工恢復草地(grass-based artificial grassland,AG)和灌木人工恢復草地(shrub-based artificial grassland,AS)4個草地類型為試驗樣地(圖1)。在每個試驗樣地,設置1 條100 m 的樣線,沿每條樣線每隔10 m 設置1個10 m×10 m 的大樣方。每個大樣方內,沿對角線均勻設置3個1 m×1 m 的小樣方。2020年7月,現場調查樣方內的植被蓋度(plant coverage,PC)和植物物種組成。調查結束后,刈割植物地上部分,帶回實驗室在80 ℃下烘干后稱重,作為植物地上生物量(aboveground biomass,Abs)。采用環刀(100 cm3)采集每個1 m×1 m 的小樣方內的0~10 cm 土壤,帶回實驗室后80 ℃下烘干至恒重后稱重,測定其土壤容重(soil bulk density,BD)。

圖1 不同草地類型景觀和稀釋曲線Fig.1 The view of different alpine grasslands and rarefaction curves
1.3 土壤樣品的采集與處理
2020年7月,在每個樣地的1 m×1 m 的樣方內,采用5 點混合采樣法鉆取0~10 cm 的土壤樣品,迅速轉移至冰盒中,帶回實驗室內過2 mm 篩后充分混合,人工揀出植物根系等雜物。從中取適量土壤樣品在105 ℃下烘干至恒重后測定土壤水分(soil moisture,SM)。采用2 mol·L-1KCl 浸提(土∶水=1∶5)土壤中的無機氮。剩余土壤樣品分為兩份,一份在-20 ℃下存貯用于土壤DNA 的提取;另一份在室內自然風干,用于土壤理化性質的測定。
1.4 土壤理化性質的測定
采用電位法(土∶水=1∶2.5)測定土壤pH;采用電導法測定電導率(electrical conductivity,EC);采用Costech元素分析儀(ECS 4010,美國)測定土壤總有機碳(total organic carbon,TOC)和全氮(total nitrogen,TN);采用酸溶-鉬銻抗比色法[21]測定全磷(total phosphorus,TP);采用NaHCO3浸提-鉬銻抗比色法進行測定土壤有效磷(available phosphorus,AP);采用NH4OAc 浸提-火焰光度計法[22]測定速效鉀(available potassium,AK);利用分段式連續流分析儀(Skarlar Analytical,荷蘭)測定硝態氮(NO3--N)和銨態氮(NH4+-N)[23]。
1.5 土壤DNA 提取與qPCR
采用E.Z.N.A.?soil DNA 試劑盒(Omega Bio-tek,美國)提取土壤樣品中的總DNA。用1%的瓊脂糖凝膠電泳和 Nanodrop?ND-2000 紫外分光光度計(NanoDrop Technologies,美國)測定260/280 nm 和 260/230 nm 的吸光值來檢測DNA 的純度和濃度。提取的DNA 稀釋后保存于TE 緩沖液中(10 mmol·L-1Tris-HCl,1 mmol·L-1EDTA,pH 8.0),并保存在-20 ℃下備用。
采用StepOne 實時熒光PCR 儀(ABI 7500,美國)測定細菌16S rRNA 基因的絕對豐度。細菌的一對擴增引物分別為 Eub338(5′-ACTCCTACGGGAGGCAGCAG-3′)和 Eub806(5′-GGACTACHVGGGTWTCTAAT-3′)。反應體系由10 μL 高靈敏性染料法定量PCR 檢測試劑(Vazyme Biotech Co.,Ltd,中國)、正向和反向引物各 0.8 μL(5 μmol·L-1)、1 μL DNA 模板和 7.4 μL 等離子水組成。擴增條件為 95 ℃下預變性 5 min,95 ℃下變性30 s,55 °C 下退火 30 s,72 ℃下延伸 1 min,共 40個循環,所有 qPCR 均重復 3 次。標準曲線由含有擴增子引物和一組含有質粒10 倍系列的稀釋液構建。細菌16S rRNA 基因拷貝數由回歸方程中的轉換拐點(cycle threshold,Ct 值)和已知的標準曲線中的拷貝數計算得到。
1.6 PCR 擴 增和 Illumina MiSeq 測 序
用與qPCR 相同的引物構建細菌群落基因文庫。具體來說,以10 ng DNA 樣品為模板在GeneAmp?9700 PCR 擴增儀(Applied Biosystems,美國)上擴增細菌 16S rRNA 基因。20 μL 的反應體系包括 4 μL FastPfu Buffer溶液、2 μL 2.5 mmol·L-1脫氧核糖核苷三磷酸(deoxyribonucleoside triphosphate,dNTPs)、正向和反向引物各0.8 μL(5 μmol·L-1)、0.4 μL FastPfu 聚合酶、10 ng DNA 模板,加等離子水至 20 μL。擴增條件如下:95 ℃預變性3 min,95 ℃變性 30 s,循環 29 次,55 ℃下退火 30 s,72 ℃下退火 45 s,72 ℃下延伸 10 min。PCR 產物用 2% 的瓊脂糖凝膠和AxyPrep DNA 凝膠試劑盒回收(Axygen Biosciences,美國),用QuantiFluor?-ST 微型熒光劑進行定量(Promega Corporation,美國)。純化后的PCR 產物混合后在Illumina MiSeq 高通量測序平臺上(Illumina,美國)從兩端進行測序,測序工作由上海美吉生物科技有限公司完成。原始測序數據上傳到美國國家生物技術信息中心(National Center for Biotechnology Information,NCBI)(https://www.ncbi.nlm.nih.gov/),SRA(Sequence ReadArchive)數據庫,登錄號為SRP311564。
1.7 數據處理與統計分析
采用豐富度實際觀測值(the observed richness,Sobs),香農指數(Shannon-Wiener index),譜系多樣性指數(phylogenetic diversity index,PD)[24],Ace 指數和 Chao1 指數[25]來衡量細菌 α 多樣性。其中,Shannon-Wiener 指數計算公式如下:
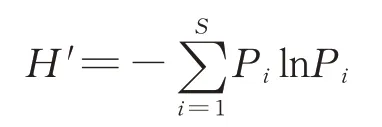
式中:Pi表示第i個物種重要值與群落物種總重要值之比,S為植物種數。
高通量測序數據的原始序列fastq 文件使用FLASH、Trimmomatic 軟件進行數據去雜和質控過濾以獲得優化序列。使用UPARSE 軟件(7.1 版,http://drive5.com/uparse/)對有效序列以97%的相似度進行聚類分析生成操作分類單位(OTUs)。使用BLAST 軟件與NCBI 中的GenBank 數據庫比較進行分類學分析。使用R 3.4.3分析Illumina MiSeq 測序數據。采用主坐標分析(principal coordinate analysis,PCoA)可視化樣本之間的Bray-Curtis 距離,以判定不同草地類型之間細菌群落結構的相似性。采用LEfSe 分析在門和綱分類水平上的差異顯著性。采用 Mantel tests、冗余分析(redundancy analysis,RDA)和方差分解分析(variance partitioning analysis,VPA)分析細菌群落結構與植被和土壤屬性之間的相關性。
2 結果與分析
2.1 植被和土壤特征
不同草地類型的植物群落結構具有明顯差異。NG 以矮嵩草、早熟禾、多色苦荬、叢生黃耆、狗娃花(Heteropappus hispidus)為優勢種;AG 以早熟禾和叢生黃耆為優勢種;AS 以檸條、紫大麥草(Hordeum violaceum)和霧冰藜(Bassia dasyphylla)為優勢種。4個樣地中,NG 的PC、植物物種豐富度(plant richness,PR)和植物香農指數(plant Shannon Wiener index,PS)最高,與 DG 有顯著差異。NG、AG 和 AS 3個樣地的PC、PS 指數和Abs 差異不顯著。然而,AS 的物種豐富度顯著高于AG(表1),表明利用灌木進行沙化高寒草地的恢復與治理,將更有助于物種的定居。
草地沙化顯著(P<0.05)降低了土壤SM(35.2%),而增加了BD(20.0%)。相反,人工植被重建則具有相反效果(表 1),AG 和 AS 的 SM 分別是 DG 的 1.5 和 1.4 倍,而其 BD 分別降低了 6.2% 和 5.0%。同時,草地沙化促使土壤pH 和NH4+-N 含量顯著增加,分別是 NG 的 1.1 和 2.2 倍,也導致土壤 EC、TOC、TN、TP、AK 和 NO3--N含量顯著下降。人工植被重建顯著提升了沙化草地的土壤EC、TOC 和NO3--N 水平。AG 土壤 TOC、TN、AP、AK 和NH4+-N 含量顯著低于AS 土壤,而其TP 和 NO3--N 含量顯著高于AS 土壤。
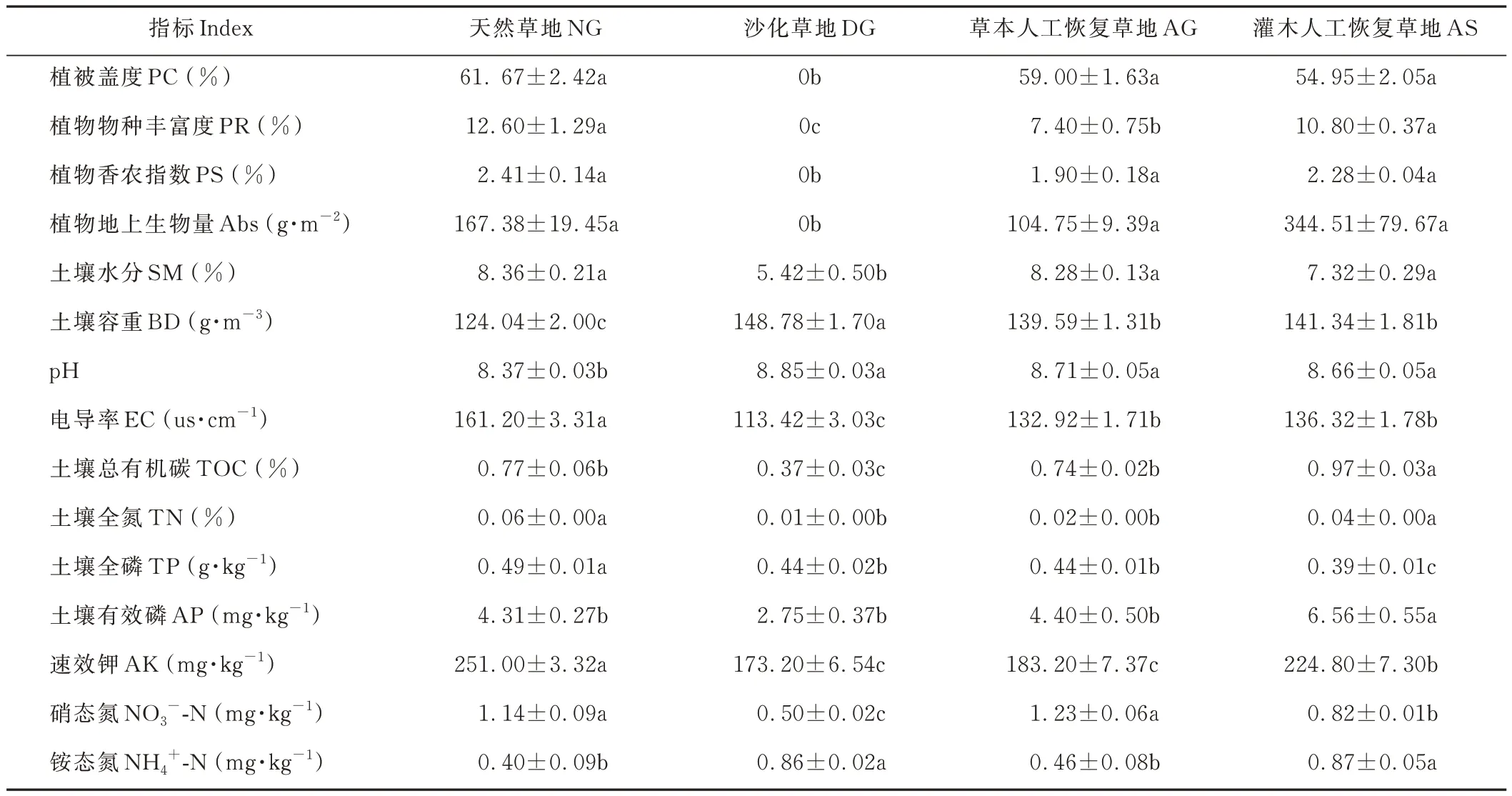
表1 不同草地類型植被和土壤特性Table 1 Vegetation and soil physiochemical properties in different grasslands
2.2 細菌群落生物量和多樣性
基于qPCR 的細菌群落生物量估計發現,NG、AG 和AS 這3個樣地土壤細菌群落的基因拷貝數均顯著高于DG,而這3個樣地之間差異不顯著(圖2)。類似地,DG 土壤的Sobs 和PD 指數顯著低于NG、AS 和AG。Shannon、Ace 和 Chao1 多樣性指數在 4個草地類型之間的差異不顯著,P值分別為 0.482、0.520 和 0.522,其在DG 土壤中均為最低,AG 和 AS 的 Ace 和 Chao1 多樣性指數與 NG 十分接近。AG 和 AS 的 Shannon、Ace 和 Chao1多樣性指數比DG 分別增加了5.4%、66.4%、64.7%和4.3%、63.8%、61.9%,表明人工植被重建有效地促進了沙化高寒草地土壤細菌群落生物量和α 多樣性的恢復。

圖2 不同草地類型土壤細菌群落生物量和多樣性指數Fig.2 Soil bacterial community biomass and diversity indices in different alpine grasslands
基于Bray-Curtis 距離的PCoA 分析結果顯示,DG、NG 與AG 和AS 的細菌群落結構在空間上明顯分離,而AG 和AS 相互分離不明顯(圖3),表明DG、NG 與AG 和AS 的細菌群落結構相似性不高,而AG和AS 則高度相似。這也反映了草地沙化和人工植被重建顯著(P=0.001)改變了高寒草地的土壤細菌群落結構,利用草本或灌木進行人工植被重建對土壤細菌群落結構的影響不大。
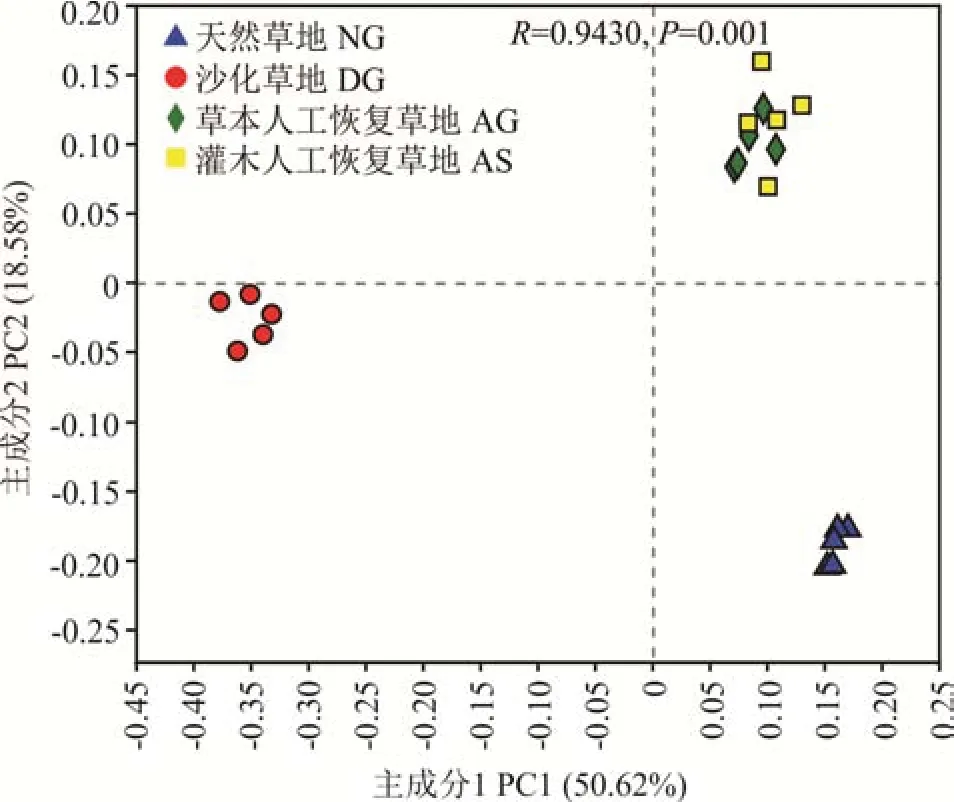
圖3 不同草地類型土壤細菌群落PCoA 分析Fig. 3 PCoA of soil bacterial communities in different grasslands
2.3 細菌群落結構與組成
Illumina Miseq 高通量測序總共得到616560 條有效 序 列 ,共 計 5505個 OTU,4個 樣 地 共 有 1831個OTU,160、182、262 和 553個 OTU 分別在 AS、AG、NG 和 DG 中是特有的(圖 4)。所有 OTU 分屬于 35個細菌門、108個細菌綱、270個細菌目、432個細菌科和763個細菌屬。如圖4所示,放線菌門(Actinobacteriota)、變形菌門(Proteobacteria)、綠彎菌門(Chloroflexi)、酸桿菌門(Acidobacteriota)為優勢細菌,在NG、DG、AG 和AS中的平均相對豐度分別為36.3%、25.1%、11.7% 和11.7%。芽單胞菌門(Gemmatimonadota)、擬桿菌門(Bacteroidota)、厚壁菌門(Firmicutes)、粘球菌門(Myxococcota)、浮霉菌門(Planctomycetota)、藍細菌門(Cyanobacteria)和未分類門(Unclassified-k—norank)細菌的平均相對豐度較低(0.5%~4.3%)。

圖4 不同草地類型土壤細菌OTUs Venn 圖和門水平群落組成Fig.4 Venn and histogram of soil bacterial community composition at OTUs and phylum level in different grasslands
如圖5 所示,4 種草地類型有9個門和27個綱的細菌相對豐度存在顯著差異(LDA≥3.0)。相對于AG 和AS,有更多的細菌門和綱在NG 和DG 土壤中明顯富集。其中,浮霉菌門、Entotheonellaeota 和Desulfobacterota 在NG 土壤中顯著(P<0.05)富集,其 LDA 值分別為 3.3、3.0 和 3.0;芽單胞菌門、放線菌門和 Methylomirabilota 在DG 土壤中明顯富集,其 LDA 值為 3.2~4.2;AG 土壤中則富集了變形菌門細菌(LDA 值=4.44);AS 土壤中富集了擬桿菌門和藍細菌門,其LDA 值分別為3.8 和3.4。從綱分類水平看,嗜熱油菌綱(Thermoleophilia)、Vicinamibacteria、紅色桿菌綱(Rubrobacteria)、厭氧繩菌綱(Anaerolineae)、浮霉菌綱(Planctomycetes)、Entotheonellia等 10個細菌綱在 NG 土壤中富集;芽單胞菌綱(Gemmatimonadetes)、酸微菌綱(Acidimicrobiia)、全噬菌綱(Holophagae)、Methylomirabilia、迷蹤菌綱(Elusimicrobia)、纖線桿菌綱(Ktedonobacteria)、Longimicrobia、Omnitrophia 等11個細菌綱富集于 DG 土壤中;α-變形菌綱(Alphaproteobacteria)、γ-變形菌綱(Gammaproteobacteria)和 Polyangia 3個細菌綱顯著富集在AG 土壤中;放線菌綱(Actinobacteria)、擬桿菌綱(Bacteroidia)和藍細菌綱(Cyanobacteria)在AS 土壤中明顯富集。

圖5 不同草地類型土壤細菌群落門和綱水平LEfSe 分析Fig.5 LEfSe analysis of soil bacterial communities in different alpine grasslands at phylum and class levels(LDA≥3.0)
分別以NG 和DG 為對照,計算相對豐度>1%的細菌門的退化程度和恢復效果,結果如表2 所示。4個樣地放線菌門、變形菌門、芽單胞菌門、擬桿菌門和浮霉菌門的相對豐度差異達到顯著水平(P<0.05),粘球菌門的相對豐度達到近顯著水平(P=0.06)。沙化后芽單胞菌門的相對豐度比NG 顯著增加了234%,而變形菌門和浮霉菌門的相對豐度則比NG 降低了23%和73%。人工植被重建后,變形菌門、擬桿菌門、粘球菌門和浮霉菌門的相對豐度門恢復程度達37%~81%。AG 和AS 的絕大部分細菌門的相對豐度與NG 的差異不顯著,表明人工植被重建22年后土壤優勢細菌門的相對豐度基本恢復到未退化前的水平。
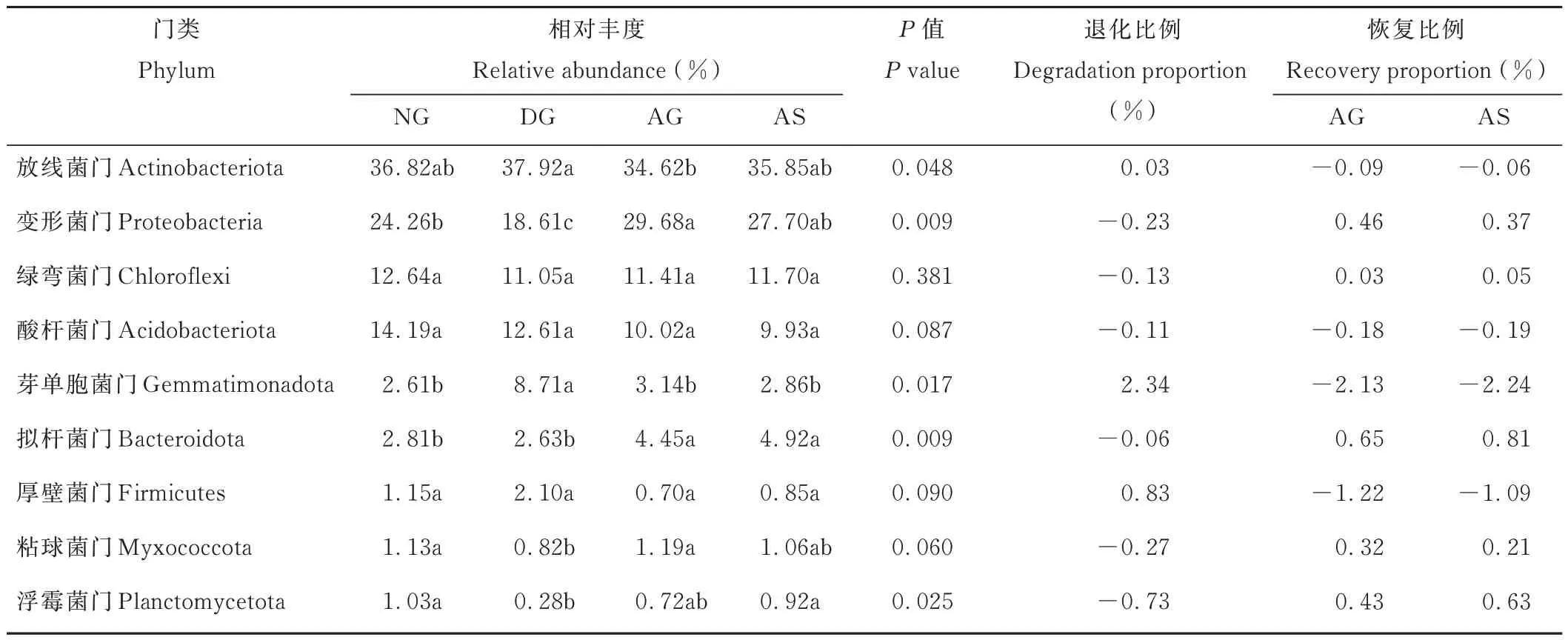
表2 不同草地類型優勢細菌門(>1%)相對豐度比較Table 2 Comparison of relative abundance of dominant bacterial phyla(>1%)in different grasslands
2.4 細菌群落結構與植被和土壤屬性的關系
Mental test 分析結果顯示,細菌群落結構與植被屬性(PC、PR、PS 指數和Abs)、土壤物理屬性(SM、BD、pH和EC)和化學屬性(TC、TN、AP、AK 和NO3--N)存在顯著相關性(P<0.05)。相比而言,細菌群落結構與植物屬性的相關性比土壤物理和化學屬性更高(表3)。植被和土壤屬性二者結合解釋了細菌群落結構70.8%的細菌群落結構變異(圖6A)。VPA 方差分解發現,植被屬性對細菌群落結構變化的解釋度最高(10.0%),土壤物理屬性次之(6.3%),土壤化學屬性最小(1.9%)。植被屬性與土壤物理屬性、植被屬性與土壤化學屬性、土壤物理屬性與化學屬性二者共同分別解釋33.7%,41.3%和51.9%的細菌群落結構的變化,上述所有指標共同解釋了72.0%的細菌群落結構變化。

圖6 土壤細菌群落結構與植物特性(V)、土壤物理(P)和化學(C)性質的RDA 和VPA 分析Fig. 6 RDA and VPA for relationships among soil bacterial community structure and vegetation(V),and soil physical(P)and chemical(C)properties
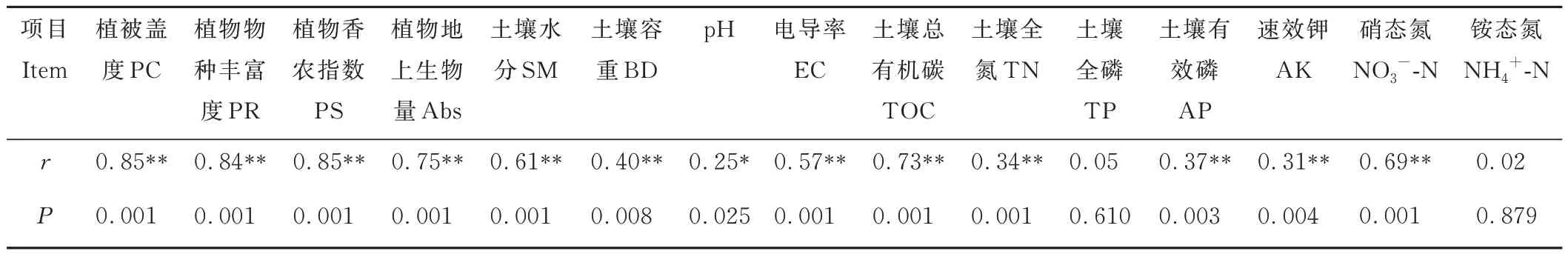
表3 土壤細菌群落結構與植被和土壤性質的相關性Table 3 Mantel tests of correlations between soil bacterial community structure and vegetation,and soil properties
3 討論
3.1 草地沙化與土壤細菌群落結構
草地退化是一個植被、土壤和微生物相互作用的復雜生態過程,伴隨著植被群落的逆向演替、物種的消亡、土壤養分狀況的惡化及其循環特征的改變等[14,26];相對于植被群落和土壤質量,土壤微生物能夠快速而敏感地感知和響應環境的變化。因此,常被作為指示環境變化的預測指標[27]。本研究發現,沙化后的高寒草地土壤細菌群落生物量和α 多樣性都顯著降低,表明高寒草地的極端退化導致了土壤細菌群落數量和多樣性的嚴重衰退。許多研究發現,土壤細菌群落結構和多樣性與植被群落結構和豐富度正相關[28-30]。植被根系分泌物和凋落物是供給土壤細菌生長和繁殖所必需的底物[25]。高寒草地沙化后,由于地上植被的衰退和消亡導致根系分泌物和凋落物的數量和多樣性銳減,致使供給土壤細菌群落的營養物質、能量和底物有效性顯著降低[31],一些土壤和根際細菌群落的數量和種類逐漸減少和消失。同時,土壤是土壤細菌群落維持其生命活動的主要場所,其理化性質顯著影響土壤細菌群落結構及其多樣性[32]。其中,土壤pH 值是決定土壤細菌群落結構和多樣性的重要指標,細菌群落的多樣性峰值往往出現在接近中性的土壤中[32-34]。另外,土壤養分有效性和水分狀況通過影響微生物細胞的代謝過程和生存策略而直接或間接地影響細菌群落結構及其多樣性。通常,細菌群落多樣性與土壤養分水平呈正比[35-36],在土壤水分適中時達到最高值[37-38]。草地沙化過程不僅降低了土壤養分水平和水分,也顯著提高了其鹽堿性,導致細菌群落生存和繁殖的環境惡劣,勢必導致細菌種群生物量及多樣性的明顯下降。再者,沙化后土壤容重的增加導致土壤通氣狀況變差,可能導致一些好氣型細菌的生長和繁殖受限。
4 種類型的草地土壤優勢細菌均為放線菌門、變形菌門、綠彎菌門和酸桿菌門,這與Zhou 等[39]對青藏高原不同退化高寒草地土壤細菌群落的研究報道基本一致。沙化后放線菌門和芽單胞菌門的相對豐度顯著增加。已有研究報道放線菌門最適生長在偏堿性的土壤環境中,且比其他細菌更耐干旱[17,40-41];芽單胞菌門更偏好土壤水分含量較低的環境[42]。本研究結果也印證了以上研究結論,同時也說明高寒草地沙化并未明顯改變土壤細菌群落的優勢分布模式,但通過顯著改變其相對比例影響其群落結構。
3.2 人工植被重建與土壤細菌群落結構
人工植被重建明顯提升了土壤細菌群落生物量和物種多樣性,22年后基本達到NG 的水平(圖2),說明在沙化草地進行人工植被重建對促進高寒草地土壤細菌數量和多樣性的恢復具有很好的效果,但仍需要較長的一段時間。與草地沙化相反,人工植被重建22年后,土壤養分水平、植被群落結構都得到了明顯提高(表1),這些土壤養分水平的提升為土壤細菌群落的生長和繁殖提供了更為多樣而豐富的底物[43-45],土壤水分狀況的改善也有益于細菌群落的生長和繁殖。同時,土壤通氣狀況也因土壤容重的下降而得到改善,為部分好氣型細菌的定居和繁殖創造了更為優越的環境條件。然而,歷經22年后,AG 和AS 的土壤細菌群落與NG 的相似性很差(圖3),表明沙化草地土壤細菌群落結構要想恢復到未退化前的狀態可能還需要更長的時間,這也充分彰顯了保護高寒草地、防止草地退化對保證青藏高原高寒草地土壤細菌群落結構的完整性和原真性具有重要生態意義。同時,也反映了僅僅用微生物生物量和物種多樣性并不能準確地評判草地質量和健康狀況,還需要考慮微生物的群落結構。
有學者認為植物的特性比植物豐富度對細菌群落結構的影響更大[30]。相比于AG,AS 草地的植物凋落物中富含來自灌木凋落物難分解的纖維素,需要擬桿菌門的大量積聚以分解纖維素[46];AS 中富集了藍細菌門(圖5)細菌,因其固氮作用[47]在一定程度上更有利于突破高寒草地生態N 限制作用,促進植被的正向演替。AG 土壤中富集了具有反硝化作用的變形菌門,能夠截留和轉化土壤中的氮素[48],有利于通過增加土壤N 的有效性促進植被正向演替。然而,AG 和AS 土壤細菌群落結構具有高度相似性特點(圖3),意味著在對沙化草地進行人工植被重建時,選擇草本物種還是灌木物種對高寒草地表層土壤細菌群落結構并無明顯影響。其可能的原因是兩種草地類型的大多數植被和土壤屬性并無顯著性差異(表1)。另外,檸條作為一種深根系灌木,其根系分泌物對表層土壤細菌群落的影響并不顯著,而對深層土壤細菌群落結構的影響程度還需要進一步深入研究。
3.3 植被和土壤屬性與土壤細菌群落結構
植被、土壤與微生物群落之間的相互關系一直是學者們探索的一個焦點。植物性狀的變化可以解釋部分土壤微生物群落組成的變化[49]。更多的植物促進了資源異質性,并以更豐富的根系分泌物和凋落物的形式為土壤細菌群落的生長和繁殖提供碳源和氮源。因此,植物多樣性越高,土壤細菌群落多樣性也越高[44-45,50]。同時,草地土壤細菌群落還會受到碳、氮等養分可利用性的顯著影響[51-52]。許多研究發現,土壤細菌的群落結構與土壤養分水平顯著正相關[35-36,52]。本研究發現,土壤細菌群落結構與絕大多數植被屬性、土壤物理屬性和化學屬性顯著正相關(表3),這一結果與其他研究報道相一致[30,35-36,53]。另外,土壤pH 值是影響土壤細菌群落結構的主要因子[32],而本研究中土壤pH 值與細菌群落結構的相關系數不高,其可能的原因是不同草地類型之間的pH 值變化不大所致。植被屬性對土壤細菌群落結構的解釋度高于土壤物理屬性和化學屬性各自的解釋度,說明植被群落對高寒草地土壤細菌群落結構具有主導作用,暗示著積極開展高寒草地植被保護和退化草地的人工植被恢復對土壤細菌群落結構的保護和恢復的重要性。此外,本研究中植被和土壤屬性(物理屬性和化學屬性)對土壤細菌群落結構變化的解釋度遠高于各自的解釋度(圖6B),這也更加證實了植被和土壤之間通過相互作用共同決定了高寒土壤細菌的群落結構,意味著加強土壤和植被保護對高寒草地細菌群落結構同樣重要。
4 結論
1)草地沙化后,土壤向貧瘠化和干旱化方向發展,土壤容重顯著增加。相反,人工植被重建顯著提升了植被的豐富度和物種多樣性、土壤養分水平,改善了水分狀況,土壤容重明顯下降。22年后絕大多數植被和土壤理化屬性基本恢復到未退化前的水平。
2)草地沙化顯著降低了土壤細菌群落的生物量和α 多樣性,人工植被重建則具有相反的效果。22年后其生物量和α 多樣性基本與未退化前的水平相持平。
3)草地沙化和人工植被重建均顯著改變了土壤細菌群落結構。其中,草地沙化后優勢細菌芽單胞菌門的相對豐度顯著增加,變形菌門和浮霉菌門則顯著降低;在人工植被重建后22年,絕大多數優勢細菌的相對豐度基本達到天然草地的水平。灌木人工恢復草地和草本人工恢復草地土壤細菌群落結構高度相似,但二者與天然草地的相似性不高。
4)相比而言,植被屬性比土壤物理屬性和化學屬性單獨對土壤細菌群落結構的解釋度更高,反映了其在驅動高寒草地土壤細菌群落結構演變中的主導作用。然而,植被和土壤二者通過相互作用共同決定青藏高原高寒草地土壤細菌群落結構。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
