蠟梅科植物次生木質部導管分子形態特征
2022-07-08 05:43:42李紅芳張小玲
廣西林業科學 2022年3期
李紅芳,薛 成,張小玲
(1.咸陽職業技術學院,陜西西安 712000;2.渭南職業技術學院,陜西渭南 714000;3.中國科學院植物研究所 系統與進化植物學國家重點實驗室,北京 100093;4.清華大學附屬中學秦漢學校,陜西咸陽712000)
蠟梅科(Calycanthaceae)是第三紀孑遺植物,該科植物是著名的觀花樹種,且具有藥用價值,為落葉或常綠灌木或小喬木,主要分布于溫帶,僅兩屬,7 ~9 種[1-2]。其中,蠟梅屬(Chimonanthus)植物和夏蠟梅屬(Calycanthus)的夏蠟梅(Calycanthus chinensis)為中國特有植物,也是我國二級重點保護植物。
蠟梅科的系統位置一直存在爭議。恩格勒(Engler)系統將蠟梅科置于毛茛目(Ranales),《中國植物志》[1]采用這種分類方式;克郎奎斯特(Cronquist)系統中,蠟梅科隸屬于樟目(Laurales)[3];哈欽松(Hutchison)系統將蠟梅科置于薔薇目(Rosales)[4];在根據分子數據建立的APG 系統中,蠟梅科歸屬于樟目(Laurales),屬基部被子植物的木蘭分支[5-6],在基部被子植物中演化程度較高,目前這種分類方式得到普遍認可。
蠟梅科的系統學研究多依據早期的形態學、解剖學和孢粉學等證據[7-10]。雖然近年來有一些基因研究[2,5],但其解剖學數據依然發揮著不可忽視的作用。導管作為被子植物的特征之一,其形態是系統學研究的重要部分。對于蠟梅科木質部解剖結構的研究不多,曾有一些對Idiospermum植物的研究[11-12];對蠟梅屬植物和夏蠟梅的研究較少,僅Metcalfe[12]對其進行了報道。近年來,掃描電子顯微鏡(SEM)開始應用于導管研究,越來越多導管分子特征被發現,如紋孔膜、原本無導管類群的植物中發現導管[13-15]等。因此,有必要對蠟梅科植物的導管分子進行進一步研究。
本研究選擇兩個中國特有種,即蠟梅(Chimonanthus praecox)(蠟梅屬)和夏蠟梅(夏蠟梅屬),利用掃描電鏡對其次生木質部導管分子進行觀察,并將結果進行展示,以期為蠟梅科植物的解剖學和系統學研究提供更多的資料和依據。
1 材料與方法
1.1 材料
選取蠟梅和夏蠟梅3年生以上枝。蠟梅取自陜西師范大學校園(108°95′E,34°20′N),夏蠟梅取自西安市植物園(108°96′E,34°20′N),均為人工栽培植株。憑證標本存放于陜西師范大學生命科學學院植物標本室(SANU)。
1.2 方法
離析法:將材料用FAA 固定,切成1 cm 長、火柴棍粗細的細條,用Jeffrey 離析液(10%鉻酸∶10%硝酸= 1∶1),40 ℃下離析約48 h。離析后噴金鍍膜,在Hitachi S-570掃描電子顯微鏡下觀察。
石蠟切片法:材料進行石蠟包埋后,石蠟切片機切片,脫蠟,CO2臨界點干燥,噴金鍍膜后在Hitachi-700掃描電子顯微鏡下觀察。
1.3 數據處理
在顯微鏡下測量導管分子的長度與直徑。采用SPSS Version 22.0 軟件進行以下分析:計算均值和變異系數;采用Student's t-test對數量性狀進行差異顯著性分析;采用Pearson相關性檢驗對數量性狀進行相關性分析。
2 結果與分析
導管作為被子植物的特征之一,在解剖學和系統學上均有重要意義。普遍認為單穿孔板、直徑變大和長度變短是導管演化的方向,網狀或梯狀穿孔板、細而長是導管相對原始的性狀[16-18]。導管和穿孔板的形態特征一定程度上可顯示一個類群的演化水平,也是系統學分類的重要形態學證據之一。
2.1 導管分子形態特征
2.1.1 蠟梅導管分子形態觀察
蠟梅次生木質部中,導管分子端壁均為單穿孔板(圖1)。穿孔板傾斜角度不同,多數穿孔板較傾斜(圖1a ~g),極少數穿孔板較平截(圖1h)。
穿孔板大多具尾,尾普遍較長,尾長度與形狀有差異。尾長度變化較大,形狀不一(圖1a ~h)。有的穿孔板尾很短(圖1a);有的穿孔板尾很長(圖1b),單側尾長度通常接近導管分子的主體長度,即尾長度占整個導管分子的1/2 ~2/3。除少數很短的尾(圖1a)外,穿孔板尾的形狀大致分為兩種,一種前端較尖(圖1d ~f),數量較多;另一種前端較圓(圖1b,g)。也有穿孔板不具尾(圖1h,j),通常較平截(圖1h),數量很少。
導管分子兩端的穿孔板形態可能相同(圖1a),也可能不同(圖1 c,g ~h)。穿孔板形態不同時,表現為一端穿孔板具尾,另一端不具尾(圖1h),或一端穿孔板具長尾,另一端具短尾(圖1 c,g)。
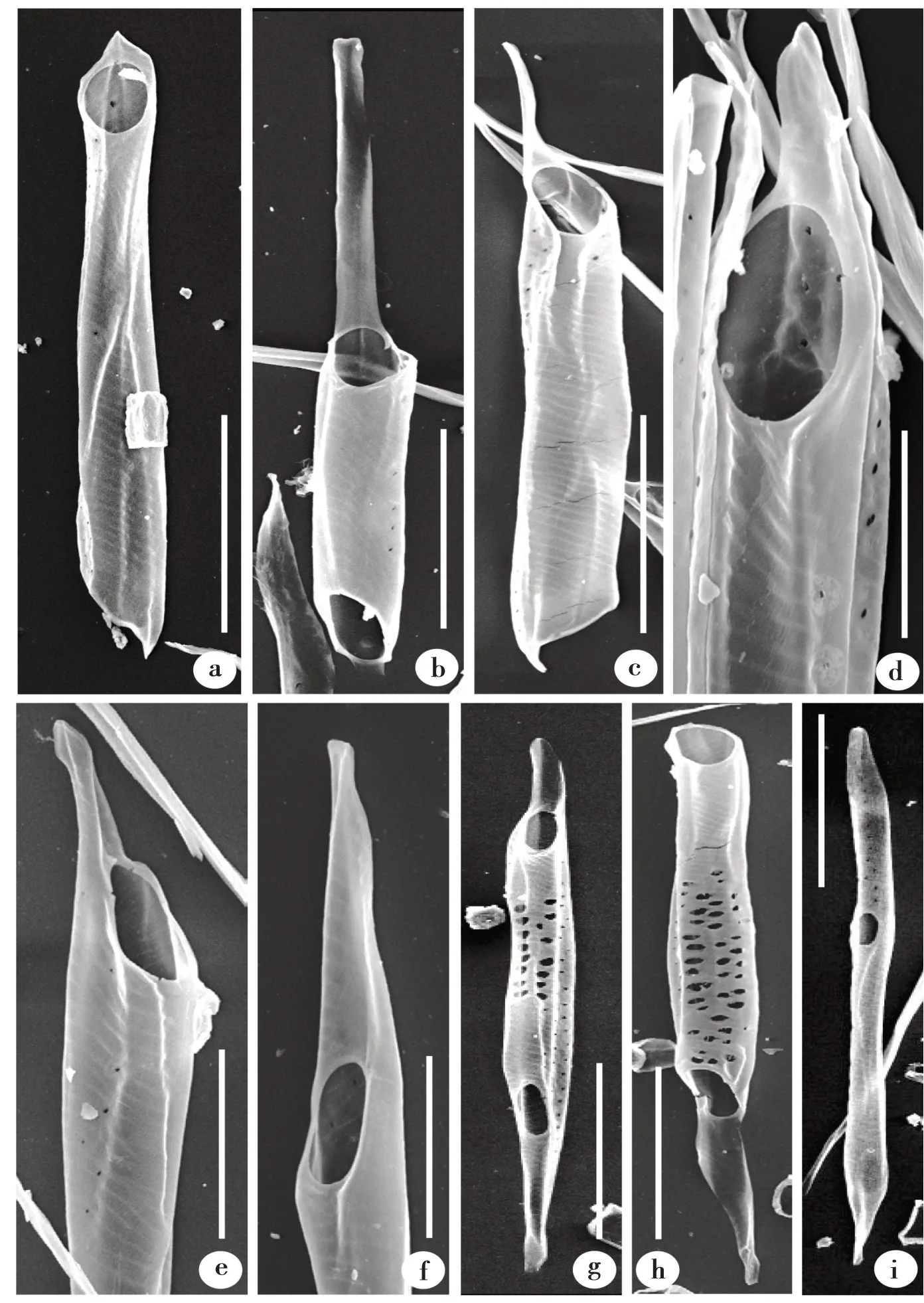

圖1 蠟梅導管分子電鏡照片Fig.1 SEM photos of vessel elements in C.praecox
觀察中還發現有穿孔板位于導管側壁(圖1i),即側壁穿孔板。導管內壁具螺紋加厚(圖1k ~l),從導管內部切片上可更清楚地看到其形態(圖1k)。
2.1.2 夏蠟梅導管分子形態觀察
夏蠟梅導管分子端壁均為單穿孔板(圖2),多數側壁不具有明顯螺紋加厚。穿孔板不具尾(圖2a ~d)或具尾(圖2e ~k)。不具尾的穿孔板通常較平截(圖2a),有些有不同程度傾斜(圖2b ~d)。穿孔板口部邊緣內斂(圖2a),石蠟切片法也觀察到這種情況(圖2c)。不具尾的穿孔板數量較多。
穿孔板尾的長度多數中等,形態較一致(圖2g ~j)。少數穿孔板具短尾(圖2e ~f)或長尾(圖2k),這些穿孔板傾斜,但角度不大。導管分子兩端的穿孔板形態可能相同(圖2l),也可能不同(圖2m)。石蠟切片法可觀察到完整的穿孔板(圖2n),其內部紋飾可見輕微加厚的較細螺紋(圖2p)。
2.2 兩種植物導管分子形態特征比較
蠟梅和夏蠟梅的導管分子均為單穿孔板,但兩者有明顯差異(圖1 ~2,表1)。

夏蠟梅導管分子中,不具尾穿孔板占比較大,約為37%;穿孔板的尾普遍比蠟梅短,僅少數穿孔板具長尾;蠟梅導管分子中,穿孔板的尾普遍較長,形態各異,僅少數穿孔板不具尾。
蠟梅導管分子中的穿孔板多數傾斜,極少數平截;夏蠟梅導管分子中的穿孔板傾斜角度小,且較一致,平截的穿孔板較多。
導管內壁上,蠟梅具有明顯的螺紋加厚,與木蘭科(Magnoliaceae)的螺紋加厚較相似[19-20]。夏蠟梅的螺紋加厚不明顯;在石蠟切片上,可觀察到其內部紋飾不呈螺旋形。
2.3 統計分析
蠟梅導管分子長度為257 ~471 μm,平均為367.0μm,變異系數為15.9%(小于20%),變異幅度小,離散程度低;直徑為21 ~60 μm,平均為40.4μm,變異系數為27.0%(大于20%),變異幅度較大,離散程度高(表1)。

表1 導管分子特征及統計分析Tab.1 Characters of vessel elements and statistical analysis
夏蠟梅導管分子長度為273 ~394 μm,平均為340.6μm,變異系數為13.3%(小于20%),變異幅度小,離散程度低;直徑為34 ~61 μm,平均為51.1μm,變異系數為32.6%(大于20%),變異幅度較大,離散程度高。
單因素方差分析t檢驗結果顯示,兩種植物導管分子的平均長度差異不顯著,平均直徑差異顯著(P<0.05)。
長度與直徑的相關性檢驗結果顯示,兩種植物導管分子的長度與直徑均呈極顯著相關(P<0.01)。李紅芳等[21]關于番荔枝科(Annonaceae)植物導管分子長度與直徑的研究結果顯示,導管分子的直徑與長度不相關。導管分子的直徑與長度是平行演化,各自獨立進行,還是有相關性的共同演化,還需進行更多的研究來證實。
夏蠟梅導管分子的平均長度小于蠟梅,但差異不顯著;蠟梅最短的導管分子比夏蠟梅短,最長的比夏蠟梅長,導管分子長度的變異系數大于夏蠟梅,離散程度比夏蠟梅高。夏蠟梅導管分子的最大直徑與最小直徑均大于蠟梅,導管分子直徑的平均值顯著大于蠟梅(P<0.05)。Metcalfe[12]研究中,蠟梅的直徑和長度與本試驗結果接近,直徑稍大;美國夏蠟梅(Calycanthus floridus)的長度更長,直徑比蠟梅小。
從以上蠟梅與夏蠟梅導管分子的比較可以看出,夏蠟梅的導管分子演化水平高于臘梅;兩種植物導管分子端壁均為單穿孔板,具有種間或屬間差異,穿孔板形態和導管的直徑均有明顯區別。這些解剖學特征具有分類學意義,一定程度上有助于進行種類鑒定。
3 討論與結論
關于蠟梅科的系統位置,有很多爭議,現在普遍認可其歸屬于樟目。在APG 系統中,蠟梅科的系統位置位于基部被子植物中的木蘭分支[5],在基部類群中是較進化的類群。蠟梅與夏蠟梅導管分子端壁均為單穿孔板,為穿孔板演化的最高水平,該結果與蠟梅科在基部被子植物中的系統位置一致。
蠟梅科通常被認為包含2 ~4 屬[22]。Takhtajan、Thome 和Dahlgren 等學者認為,蠟梅科包含4個屬,分別為蠟梅屬(Chimonanthus)、美國夏蠟梅屬(Calycanthus)、中國夏蠟梅屬(Sinocalycanthus)和Idiospermum[10,23-26]。其中,Idiospermum被一些學 者認為應當獨立成科[3,27],但更多的人認為其應歸屬于蠟梅科[12,23,28-29],主張將其歸入美國夏蠟梅屬[12,23]。
另一個有爭議的地方為中國夏蠟梅屬(Sinocalycanthus)是否應當獨立成屬。Thome、Dahlgren、Mabberley 和吳征鎰等學者根據解剖學特征認為,中國夏蠟梅屬應獨立成屬[23-25,30-31];有些學者不支持獨立成屬,認為應將其并入美國夏蠟梅屬[12,28-29]。APG系統將蠟梅科分為蠟梅屬和夏蠟梅屬。本研究根據APG 系統,按照《中國植物志》的分類方式,將中國夏蠟梅屬歸入美國夏蠟梅屬,并按中國習慣稱之為夏蠟梅屬(Calyanthus)。
多數觀點認為夏蠟梅屬(包括美國夏蠟梅屬(Calycanthus)和中國夏蠟梅屬(Sinocalycanthus))較原始,蠟梅屬較進化;形態學和孢粉學等特征表明夏蠟梅在演化上處于較原始的位置[7,28,32]。李燁等[33]研究發現,蠟梅屬植物的葉脈類型較原始;李燁等[10]對蠟梅科的葉表皮細胞進行研究,認為蠟梅屬植物的氣孔器較原始,夏蠟梅屬(Sinocalycanthus)和美國夏蠟梅屬植物的氣孔器較進化。本研究中,夏蠟梅的導管分子比蠟梅短且粗,穿孔板傾斜角度比蠟梅小,且穿孔板中不具尾的比例較高,顯示夏蠟梅導管分子的演化水平比蠟梅更高,此結論與李燁等研究結果一致。
原始性狀與進化性狀可能在同一物種中并不統一,可能互相嵌合,不能單純依據導管或其他木質部特征去推論夏蠟梅和蠟梅的演化水平,但這些特征可為系統學研究提供重要的參考。蠟梅科作為被子植物的基部類群,其穿孔板的相關特征對于導管的演化研究也有重要意義。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
