大白菜LACS 家族基因鑒定與表達(dá)分析
2022-07-09 04:00:52王榮花王樹彬劉栓桃張志剛李巧云王立華趙智中
山東農(nóng)業(yè)科學(xué) 2022年6期
關(guān)鍵詞:分析
王榮花,王樹彬,劉栓桃,張志剛,李巧云,王立華,趙智中
(山東省農(nóng)業(yè)科學(xué)院蔬菜研究所/國家蔬菜改良中心山東分中心/山東省設(shè)施蔬菜生物學(xué)重點(diǎn)實(shí)驗(yàn)室/農(nóng)業(yè)部黃淮地區(qū)蔬菜科學(xué)觀測實(shí)驗(yàn)站(山東),山東 濟(jì)南 250100)
脂酰輔酶A 合成酶(fatty acyl-CoA synthetase,ACS)存在于內(nèi)質(zhì)網(wǎng)線粒體外膜上,根據(jù)碳鏈的長短被分為4 類:短鏈脂酰輔酶A 合成酶(C2~C4)、中鏈脂酰輔酶A 合成酶(C4~C12)、長鏈脂酰輔酶A 合成酶(C12~C20)及超長鏈脂酰輔酶A 合成酶(>C20)。 其中,長鏈脂酰輔酶A合成酶(LACS)廣泛存在于各種生物中,在脂肪酸合成和分解代謝中發(fā)揮著重要作用[1]。 研究人員在擬南芥基因組中鑒定到9 個(gè)LACS 成員,分別為AtLACS1 ~ AtLACS9[2],其中,AtLACS1、At-LACS2和AtLACS4基因在蠟質(zhì)合成和角質(zhì)合成途徑中起著重要的作用[3-5],且AtLACS1和AtLACS4在花粉壁形成中也具有一定功能[5];AtLACS6和AtLACS7基因產(chǎn)物定位在過氧化體中,對(duì)脂肪酸β-氧化有加性效應(yīng),參與擬南芥中脂肪酸的降解[6];轉(zhuǎn)化AtLACS9可促進(jìn)大豆的油脂合成[7]。在油菜中的研究表明,BnLACS2的轉(zhuǎn)錄增加了種子中參與糖酵解、脂肪酸和脂質(zhì)合成基因的表達(dá),是影響甘藍(lán)型油菜種子產(chǎn)油的一個(gè)重要因素[8];BnLACS9參與了葉綠素的生物合成[9]。
大白菜(Brassica rapaL. ssp.pekinensis)屬于十字花科蕓薹屬,是我國重要的十字花科蔬菜,隨著生活水平的提高,人們對(duì)其品質(zhì)的要求不斷提高。LACS基因家族在植物脂質(zhì)的合成和分解代謝中起著重要作用,研究其家族成員特性對(duì)提高大白菜品質(zhì)具有一定意義。LACS家族成員已在擬南芥[10]、小麥[11]、棉花[12]、大豆[13]、油菜[14]等物種中被鑒定出,但基因序列在不同物種間存在較大差異,而蛋白序列相對(duì)保守[15]。 目前大白菜LACS家族基因的研究尚未見報(bào)道。 因此,本研究以大白菜Chiifu 基因組數(shù)據(jù)作參考[16],通過生物信息學(xué)方法鑒定大白菜LACS 家族成員,并對(duì)其進(jìn)行系統(tǒng)發(fā)育樹構(gòu)建、染色體定位、基因結(jié)構(gòu)及表達(dá)模式分析,為深入研究大白菜LACS家族基因奠定基礎(chǔ)。
1 材料與方法
1.1 大白菜LACS 家族基因的鑒定
從TAIR 數(shù)據(jù)庫(http:/ /www. arabidopsis.org/)下載了擬南芥LACS 家族蛋白序列共計(jì)9 條(AtLACS1~AtLACS9)[15]。 在BRAD 數(shù)據(jù)庫(http:/ /brassicadb. org/brad/)[16,17]下 載 大 白 菜(V3.0版本)和甘藍(lán)(Braol JZS_V2.0 版本)全基因組蛋白序列文件和gff3 文件構(gòu)建本地BLASTP 數(shù)據(jù)庫,將AtLACS 家族的蛋白序列通過本地BLASTP 比對(duì)出大白菜(Br)LACS 和甘藍(lán)(Bo)LACS 的家族成員。BrLACS和BoLACS基因的命名根據(jù)其與AtLACS1~AtLACS9序列的同源性及共線性關(guān)系進(jìn)行,部分同源基因通過添加小寫英文字母后綴(a、b、c 等)進(jìn)行區(qū)分。
1.2 大白菜LACS 蛋白特征預(yù)測、系統(tǒng)發(fā)育、進(jìn)化樹分析及保守基序分析
利用在線ProtParam 工具(https:/ /web.expasy.org/protparam/)對(duì)獲得的BrLACS 蛋白質(zhì)序列分子量、理論等電點(diǎn)和親水性平均系數(shù)進(jìn)行分析。 利用Gene Structure Display Server(GSDS 2.0,http:/ /gsds.gao-lab.org/)根據(jù)每個(gè)BrLACS基因組序列和相應(yīng)的CDS 序列繪制基因結(jié)構(gòu)圖[18]。利用MEGA 7.0 對(duì)BrLACS、AtLACS 和BoLACS 蛋白質(zhì)序列繪制系統(tǒng)進(jìn)化樹,采用鄰接法(Neighborjoining),設(shè)置Bootstrap 值為1 000,其他參數(shù)保持默認(rèn)值[19]。 使用模體分析工具M(jìn)EME(http:/ /meme-suite.org/)對(duì)BrLACS 蛋白進(jìn)行保守基序分析,其中最適基序?qū)挾仍O(shè)置為6~50,最大基序設(shè)置為10,其他為默認(rèn)參數(shù)。
1.3 大白菜LACS 基因的染色體定位及共線性分析
根據(jù)BrLACS基因在染色體上的物理位置,使用MapChart 2.30[20]對(duì)BrLACS基因進(jìn)行染色體定位。 利用BRAD(http:/ /brassicadb.org/brad/searchSyntenytPCK.php)數(shù)據(jù)庫分析大白菜、甘藍(lán)和擬南芥之間的親緣關(guān)系,利用TBtools[21]繪制共線性圖。
1.4 大白菜LACS 基因在不同器官/組織以及不同材料中的表達(dá)模式分析
2 結(jié)果與分析
為了分析大白菜不同器官/組織中BrLACS基因的表達(dá)水平,在NCBI 的GEO 數(shù)據(jù)庫下載大白菜不同器官/組織(愈傷組織、根、莖、葉、花和角果) 轉(zhuǎn) 錄 組 數(shù) 據(jù)(登 錄 號(hào): GSE43245), 以BrLACS基因的log2(FPKM+1)(fragments per kilobase per million mapped reads)值來表示BrLACSs在不同器官/組織中的表達(dá)水平,并繪制熱圖。
前期構(gòu)建了大白菜花莖有蠟粉組(RHL065_1_T1、RHL065_1_T2、RHL065_1_T3)與無蠟粉組(RHL065_2_T1、RHL065_2_T2、RHL065_2_T3)兩組cDNA 文庫,并對(duì)文庫質(zhì)量檢測后應(yīng)用Illumina HiSeq 平臺(tái)進(jìn)行測序[22]。 根據(jù)RNA-Seq 數(shù)據(jù)中獲取的每個(gè)BrLACS基因的FPKM 值來繪制柱形圖。
2.1 大白菜LACS 家族基因的鑒定、系統(tǒng)發(fā)育和蛋白特征分析
通過對(duì)全基因組蛋白質(zhì)數(shù)據(jù)庫的鑒定和結(jié)構(gòu)域驗(yàn)證,在大白菜和甘藍(lán)基因組中分別鑒定到13個(gè)BrLACS 和13 個(gè)BoLACS 蛋白(圖1,表1)。 從大白菜、甘藍(lán)和擬南芥LACS 蛋白序列的無根系統(tǒng)進(jìn)化樹可見,這些LACS 蛋白成員根據(jù)親緣關(guān)系的遠(yuǎn)近被分為4 組,Group A、B、C、D 分別包含7、14、8、6 個(gè)LACS 成員(圖1)。 其中,Group A 包括BrLACS1a、BrLACS1b 和BrLACS1c,Group B 包括BrLACS2、 BrLACS3、 BrLACS4、 BrLACS5a 和BrLACS5b,Group C 包括BrLACS6a、BrLACS6b 和BrLACS7,Group D 包括BrLACS8 和BrLACS9。
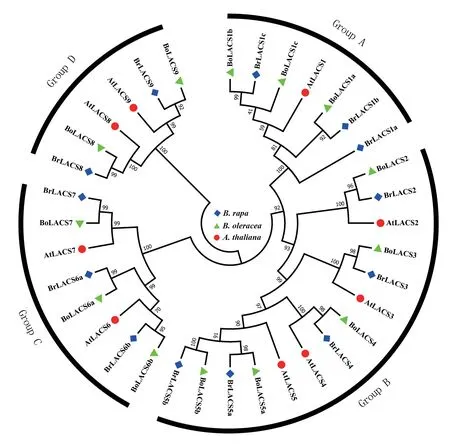
圖1 大白菜(Br)、甘藍(lán)(Bo)、擬南芥(At)LACS 蛋白系統(tǒng)進(jìn)化樹分析
通過在線ProtParam 工具預(yù)測了BrLACS 蛋白的物理和化學(xué)特征,結(jié)果(表1)顯示,BrLACS蛋白長度為199 ~ 1 072 aa,分子量為23.10~121.10 kDa,理論等電點(diǎn)為5.66~9.21,總平均親水性均為負(fù)值(-0.398~-0.049),為親水性蛋白。
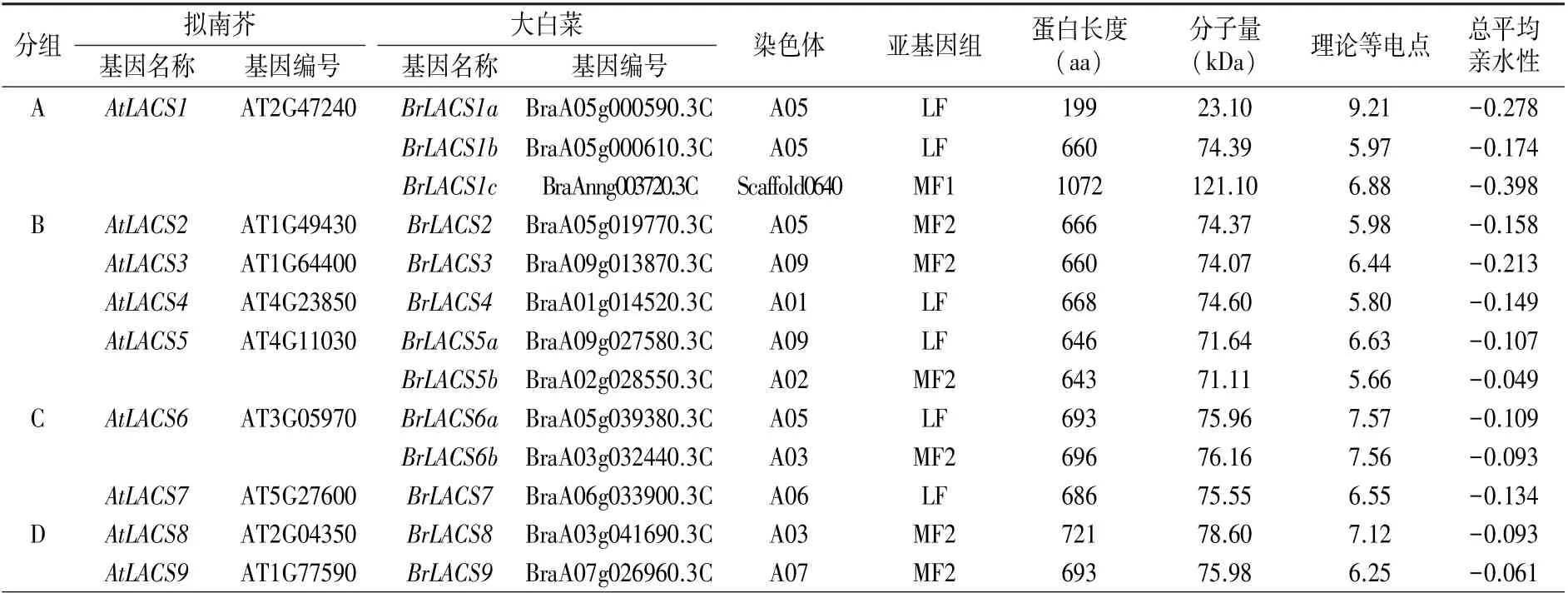
表1 大白菜和擬南芥相對(duì)應(yīng)LACS 家族成員信息
2.2 大白菜LACS 基因的染色體分布和共線性分析
從表1 可以看出,BrLACS家族基因的13 個(gè)成員分布在3 個(gè)亞基因組上,其中亞基因組LF 有6 個(gè)BrLACS基因,MF1 有1 個(gè)BrLACS基因,MF2有6 個(gè)BrLACS基因。 與AtLACS基因相比,BrLACS基因在蕓薹族特異的全基因組三倍化事件后均被保留。 總共有6 個(gè)BrLACS基因(BrLACS2、BrLACS3、BrLACS4、BrLACS7、BrLACS8和BrLACS9)保留了單拷貝,2 個(gè)BrLACS基因(BrLACS5和BrLACS6) 保 留 了 雙 拷 貝, 1 個(gè)BrLACS基因(BrLACS1)保留了三拷貝。 13 個(gè)BrLACS基因中,BrLACS1c只錨定到Scaffold0640上,未能錨定到大白菜的任何染色體上,其余12個(gè)則被定位在7 條染色體上,其中,BrLACS4位于染色體A01 上,BrLACS5b位于染色體A02 上,BrLACS6b和BrLACS8位 于 染 色 體 A03 上,BrLACS1a、BrLACS1b、BrLACS2和BrLACS6a位于染色體A05 上,BrLACS7位于染色體A06 上,BrLACS9位 于 染 色 體 A07 上,BrLACS3和BrLACS5a位于染色體A09 上(圖2)。
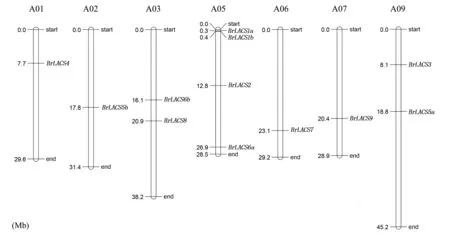
圖2 大白菜LACS 基因在7 條染色體上的定位分布
將鑒定到的直系和旁系同源LACS基因用于分析BrLACS、BoLACS和AtLACS基因間的共線性,結(jié)果鑒定到大白菜之間3 對(duì)旁系同源基因、大白菜與擬南芥之間12 對(duì)直系同源基因以及大白菜與甘藍(lán)之間20 對(duì)直系同源基因(圖3)。
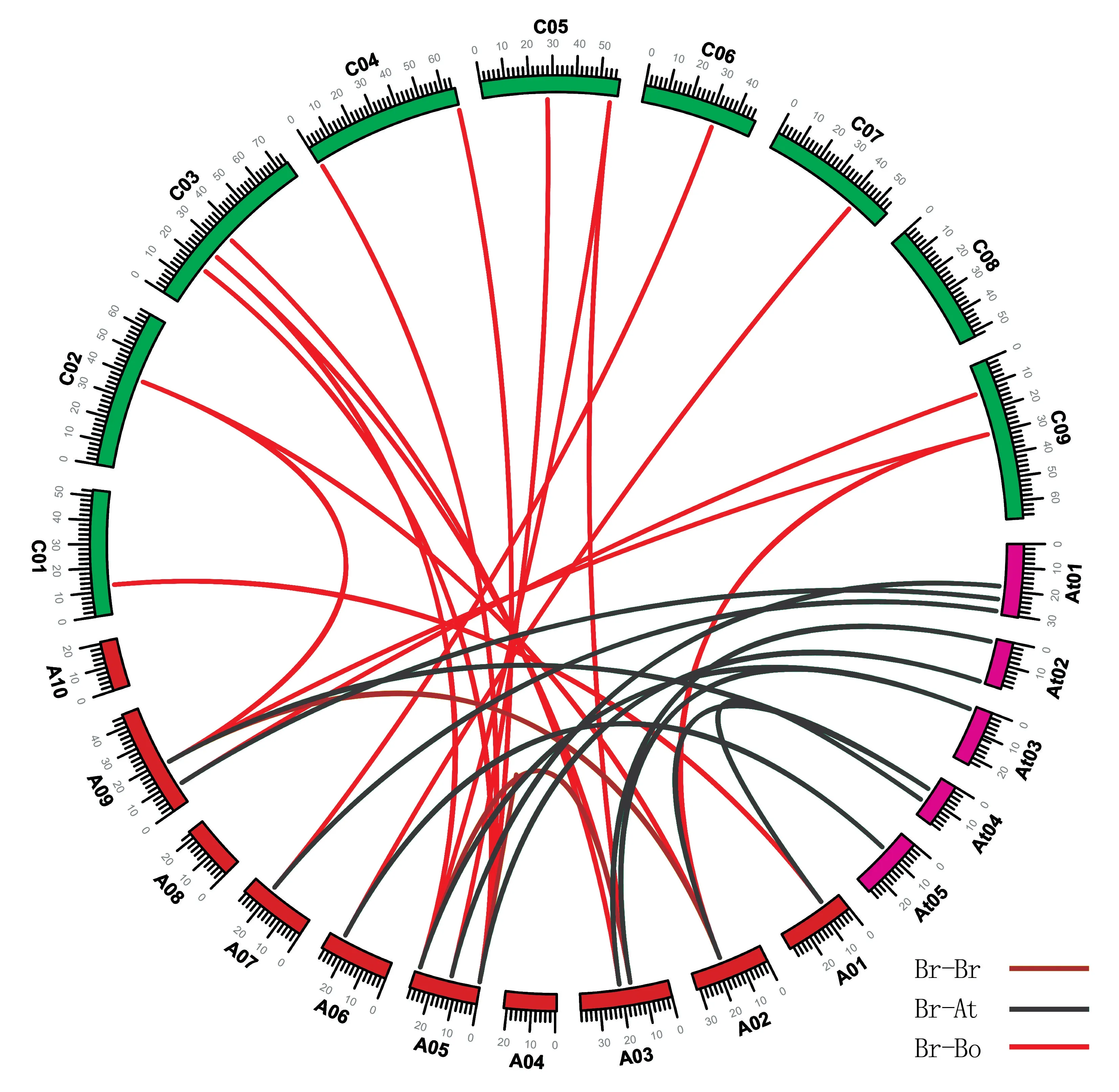
圖3 大白菜(Br)、擬南芥(At)和甘藍(lán)(Bo)LACS 基因的共線性關(guān)系
2.3 大白菜LACS 基因結(jié)構(gòu)分析
根據(jù)BrLACS家族成員的CDS 和DNA 序列,繪制BrLACS基因結(jié)構(gòu)圖,結(jié)果(圖4) 顯示,BrLACS家族成員之間的外顯子數(shù)量差異較大(4~32 個(gè))。 其中BrLACS1a含有4 個(gè)外顯子,BrLACS9含有10 個(gè)外顯子,BrLACS8含有11 個(gè)外顯子,BrLACS1b含有17 個(gè)外顯子,BrLACS3、BrLACS4和BrLACS5a含有18 個(gè)外顯子,BrLACS2含有19 個(gè)外顯子,BrLACS5b含有20 個(gè)外顯子,BrLACS6a、BrLACS6b和BrLACS7含有23 個(gè)外顯子,BrLACS1c基因均含有32 個(gè)外顯子。

圖4 大白菜LACS 家族基因外顯子-內(nèi)含子結(jié)構(gòu)分析
2.4 大白菜LACS 蛋白保守結(jié)構(gòu)域分析
利用MEME 在線分析工具對(duì)BrLACS 家族蛋白保守結(jié)構(gòu)域進(jìn)行分析,共獲得10 個(gè)保守基序,分別命名為Motif1~Motif10。 13 個(gè)BrLACS 家族成員中,有9 個(gè)成員均含有Motif1~Motif10 保守基序,而BrLACS1a 中只有motif 1 和motif 3,BrLACS7 中沒有motif 9,BrLACS8 和BrLACS9 中沒有motif 8(圖5)。

圖5 大白菜LACS 家族成員蛋白保守結(jié)構(gòu)域分析
2.5 大白菜LACS 家族基因表達(dá)分析
2.5.1 組織特異性表達(dá)分析 通過分析BrLACS家族基因在大白菜不同器官中的表達(dá),結(jié)果(圖6)顯示,大多數(shù)BrLACS在大白菜花中高表達(dá),其次是在角果、葉、根和莖中;BrLACS4和BrLACS8在各器官/組織中的整體表達(dá)量最高,BrLACS5a和BrLACS5b在各器官/組織中的整體表達(dá)量最低。 花中表達(dá)量較高的是BrLACS8、BrLACS6a和BrLACS4,角果中表達(dá)較高的是BrLACS4、BrLACS9和BrLACS8,葉中表達(dá)量較高的是BrLACS4、BrLACS9和BrLACS1b;各器官/組織中均未找到BrLACS1a基因的表達(dá)。
2.5.2 在有蠟粉和無蠟粉大白菜中的表達(dá)分析如圖7 所示,總體而言,BrLACS1b、BrLACS2、BrLACS4、BrLACS5a、BrLACS5b、BrLACS6a、BrLACS8和BrLACS9基因在有蠟粉大白菜中的表達(dá)量顯著高 于 無 蠟 粉 大 白 菜;BrLACS3、BrLACS6b和BrLACS7基因在有蠟粉大白菜中的表達(dá)量顯著低于無蠟粉大白菜;BrLACS1c在兩組大白菜中的表達(dá) 量 都 很 高, 但 差 異 不 顯 著。BrLACS3、BrLACS5a、BrLACS5b和BrLACS7整體表達(dá)水平都很低。
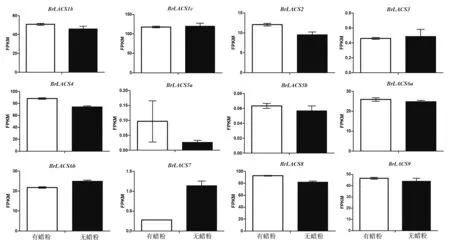
圖7 BrLACS 基因在有蠟粉和無蠟粉大白菜材料中的表達(dá)分析
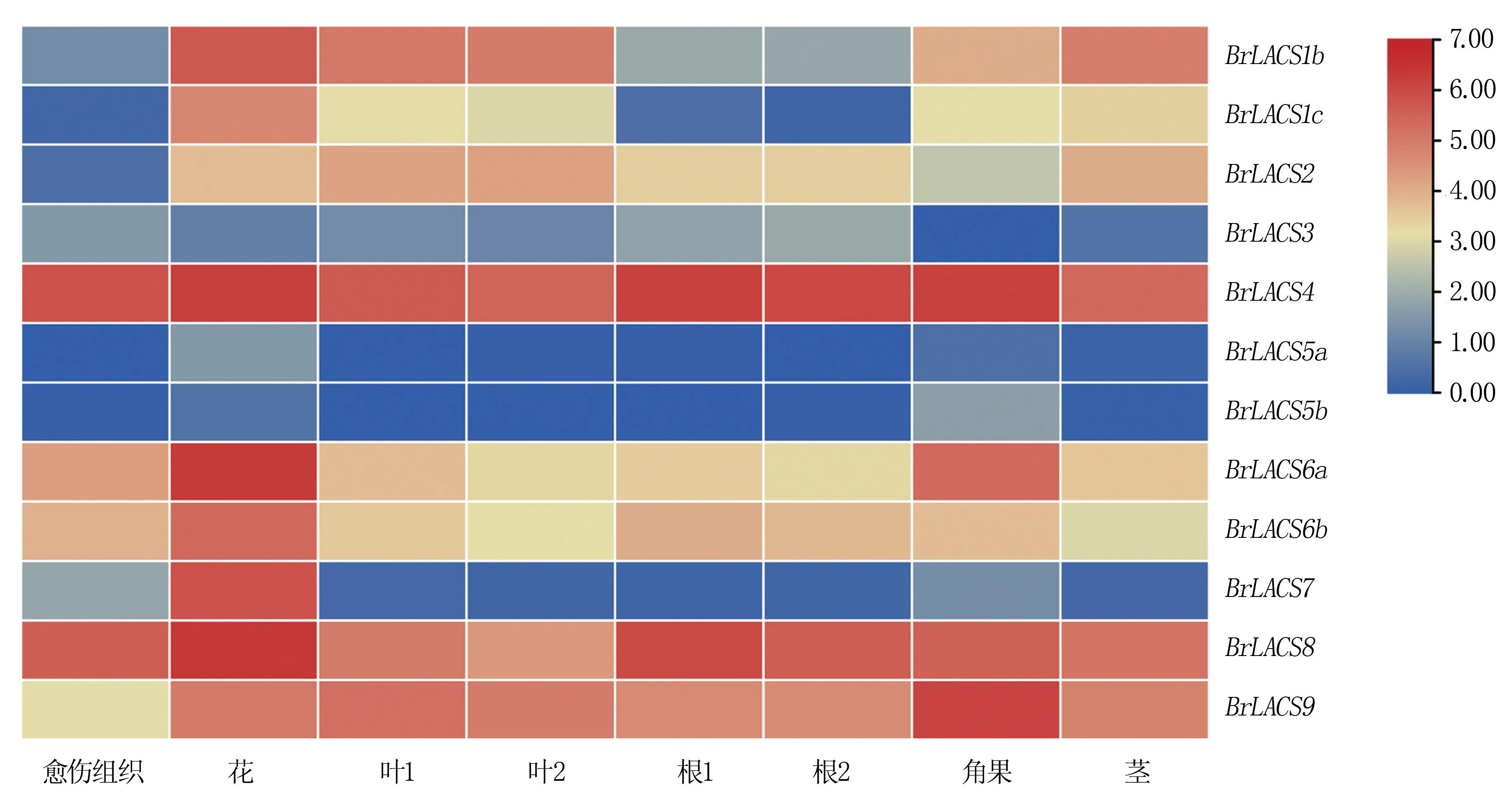
圖6 大白菜LACS 基因在不同組織/器官中的表達(dá)分析
3 討論
植物表皮蠟粉主要由超長鏈脂肪酸及其衍生物組成,在植物抗干旱、抵御病害等生物與非生物脅迫方面起著重要作用[22,23]。 同時(shí)對(duì)于大白菜等葉用蔬菜作物特別是白菜薹品種,蠟粉還是重要的商品性狀。 LACS 是脂肪酸代謝途徑中的關(guān)鍵酶,催化長鏈酰基輔酶A 的合成,能夠在多種生物過程中發(fā)揮作用[15,24-26]。
隨著基因組學(xué)的快速發(fā)展,越來越多的LACS家族基因成員被分離鑒定[1,15],擬南芥中發(fā)現(xiàn)了9 個(gè)LACS基因[10],小麥中發(fā)現(xiàn)了148 個(gè)LACS基因[11],陸地棉中發(fā)現(xiàn)了21 個(gè)LACS基因[12],油菜中發(fā)現(xiàn)了34 個(gè)LACS基因[14]。 本研究發(fā)現(xiàn)大白菜和甘藍(lán)中分別有13 個(gè)BrLACSs和13 個(gè)BoLACSs基因,均多于AtLACSs的數(shù)量,但不是理論上的三倍數(shù)量關(guān)系,這可能是蕓薹屬特有的全基因組三倍化事件以及偏性基因丟失事件造成的[16,27]。對(duì)擬南芥、大白菜和甘藍(lán)的共線性分析結(jié)果顯示,大白菜與擬南芥發(fā)生分化之后,BrLACS5和BrLACS6保留了雙拷貝,BrLACS1保留了三拷貝,其余BrLACSs均保留了單拷貝。
本研究結(jié)果顯示,大白菜LACS基因成員間在外顯子-內(nèi)含子結(jié)構(gòu)上具有較大差異,而外顯子-內(nèi)含子結(jié)構(gòu)差異在基因家族的進(jìn)化中起著重要作 用[28]。 此 外, 旁 系 同 源 基 因BrLACS1a、BrLACS1b和BrLACS1c之間發(fā)生了明顯可變剪切和內(nèi) 含 子 插 入, 導(dǎo) 致BrLACS1a、BrLACS1b與BrLACS1c基因之間出現(xiàn)了功能的多樣性,BrLACS1b與BrLACS1c在不同器官/組織以及有蠟粉和無蠟粉大白菜中表達(dá)水平差異顯著,而BrLACS1a基因在以上分析中均沒有表達(dá)量。
擬南芥LACS 家族的9 個(gè)成員中,已發(fā)現(xiàn)LACS1、LACS2和LACS4基因在表皮蠟質(zhì)合成和角質(zhì)合成途徑中起著重要的作用[3-5]。 研究發(fā)現(xiàn),擬南芥lacs1突變體表現(xiàn)出蠟質(zhì)含量減少,lacs2突變體也表現(xiàn)出角質(zhì)減少,而lacs1和lacs2的擬南芥雙突變體相對(duì)于各自單變體,蠟質(zhì)的總量有所降低且表皮滲透性增強(qiáng)[3,4];擬南芥lacs4突變體中蠟質(zhì)含量減少,占野生型的73%,lacs1突變體中蠟質(zhì)含量占野生型的35%,lacs1和lacs4雙突變體蠟質(zhì)含量占野生型的23%[5]。 擬南芥的外表皮蠟質(zhì)主要以柱狀晶體形式存在于莖、莢果和柱頭上,以非晶體形式存在于葉片上[29-31]。本研究發(fā)現(xiàn),大部分BrLACS家族成員在大白菜花中高表達(dá),而在愈傷組織中表達(dá)量很低;BrLACS4在各器官/組織中整體表達(dá)量最高,BrLACS8在各器官/組織中整體表達(dá)量高且在有蠟粉大白菜中的表達(dá)量顯著高于無蠟粉大白菜。本研究結(jié)果可為后續(xù)進(jìn)一步開展BrLACS8基因在控制大白菜蠟質(zhì)性狀中的具體功能研究奠定基礎(chǔ)。
4 結(jié)論
本研究在大白菜全基因組中共篩選出13 個(gè)BrLACS基因家族成員,可分為4 組,不均勻地分布在7 條染色體和1 個(gè)Scaffold 上。BrLACS4、BrLACS8在大白菜各器官/組織中表達(dá)量高且在有蠟粉大白菜中的表達(dá)量顯著高于無蠟粉大白菜,其可能在控制大白菜蠟質(zhì)性狀中發(fā)揮重要的作用。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會(huì)科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
