梓醇調控miR-143-3p干預Th17糖酵解和細胞分化的機制
2022-07-11 05:07:52申美玉榮秋妮狄昱希田鋒祥張明菲蔣寶平周玲玲
中國藥理學通報 2022年7期
申美玉,榮秋妮,狄昱希,田鋒祥,張明菲,王 祥,蔣寶平,周玲玲
(南京中醫藥大學藥學院,江蘇 南京 210023)
類風濕關節炎(rheumatoid arthritis,RA)是一種以關節滑膜炎癥為特征的慢性自身免疫性疾病,涉及心血管等多系統病變,其病理機制和生物標志物目前尚未完全闡明。CD4+輔助性T細胞(CD4+helper cells,Th)是適應性免疫細胞的主要群體之一,在不同的免疫環境下,na?ve CD4+T細胞能夠激活并分化成為不同的亞群(Th1/Th2/Th17/Tfh/Treg等)。有研究顯示[1],Th17細胞的過度分化可促進RA的發展。因此,尋找造成RA中Th17細胞異常分化的關鍵因素對RA的防治至關重要。
免疫代謝研究表明,T細胞各亞群具有不同的代謝程序,其中Th17細胞特別依賴于糖酵解[2]。而RA疾病狀態下為保持持續的滑膜組織浸潤,其CD4+T細胞始終處于高合成代謝的細胞增殖狀態,主要表現為葡萄糖攝取異常增加、糖酵解增強、線粒體有氧氧化減弱,為Th17細胞分化提供了合適的條件[3]。因此,靶向CD4+T細胞中關鍵糖酵解酶葡萄糖轉運蛋白(Glut1)、己糖激酶(HK2)、丙酮酸激酶(PKM2)、乳酸脫氫酶(LDHA)等的表達可為臨床干預RA提供更多的治療方案。
MicroRNA(miRNA)是一種內源性的保守小分子,可以通過與靶分子mRNA的3′末端非翻譯區(3′UTR)結合而發揮轉錄后基因調節功能。多種miRNA被發現在RA CD4+T細胞中異常表達[4],并且可以通過多種機制調節細胞的糖脂代謝[5],這提示我們RA中miRNA表達異常可能通過調控糖酵解來介導Th17細胞的過度分化。最新研究顯示[6],miR-143-3p可能是T細胞分化和功能的關鍵調節靶點,miR-143-3p過表達可通過靶向Glut1抑制葡萄糖攝取,促進記憶CD8+T細胞的分化并維持其抗腫瘤功能。同時,miR-143-3p可增加線粒體呼吸鏈復合物的表達水平,提示miR-143-3p的下調可能介導了細胞從線粒體有氧氧化向糖酵解的代謝轉換[7]。
生地具有清熱生津、涼血止血等功效,臨床可治療以“陰虛絡熱”為主要病機的RA,其主要活性成分梓醇(catalpol,CAT)可明顯改善Th17細胞異常分化,減少炎性因子分泌,緩解RA病情[8],但其作用機制的研究還不完善。項目組前期研究已發現,CAT對miR-143-3p的表達具有調控作用,且能夠顯著影響CD4+T細胞中糖酵解產物水平。因此,本研究擬選取RA患者外周血CD4+T細胞,檢測miR-143-3p及相關糖酵解關鍵酶的異常表達情況,初步分析二者的相關性,并建立體外誘導Th17細胞分化模型,結合miR-143-3p慢病毒轉染,探討CAT調控miR-143-3p對Th17糖酵解和細胞分化的干預作用和作用機制,為RA等慢性復雜疾病的臨床用藥提供科學依據。
1 材料與方法
1.1 材料與儀器
1.1.1RA病例 經南京中醫藥大學附屬醫院倫理委員會審核(2016NL-KS14),依據美國風濕病學會(ACR)1987年發布的診斷標準,選擇10例RA活動期患者。所有RA患者未進行免疫治療。健康對照組另收集5例健康志愿者(HCs)。
1.1.2動物 10只6~8周齡的SPF級♂ C57/BL6小鼠,購自浙江維通利華實驗動物技術有限公司,實驗動物合格號SCXK(浙)2019-0001。動物在南京中醫藥大學實驗動物中心飼養,自由飲食、飲水,遵循自然晝夜節律,室溫為(23±2)℃,濕度為0.4~0.6。南京中醫藥大學動物倫理委員會審核、批準所有動物操作(202103A010)。
1.1.3藥物 梓醇,購自上海源葉生物科技有限公司(貨號:B21678,純度≥0.98)。
1.1.4主要試劑 人CD4+T細胞分選磁珠(貨號557767),購自美國BD公司;小鼠na?ve CD4+T細胞分選試劑盒(貨號130-104-453)、MACS分選緩沖液(貨號130-091-221)、LS柱(130-042-401),購自德國美天旎公司;Mouse anti-CD3e(貨號553057)、Mouse anti-CD28(貨號553294),購自美國BD公司;Mouse IL-6(貨號CG39)、Mouse IL-23(貨號CI18)、Mouse TGF-β(貨號CA59),購自上海近岸科技有限公司;RPMI1640培養基(貨號C11875500B)、TRIzol裂解液(貨號15596026),購自美國賽默飛世爾科技公司;紅細胞裂解液(貨號R1010),購自北京索萊寶科技有限公司;miR-143-3p mimics(貨號133F0E6)、miR-143-3p inhibitor(貨號1341413)、轉染陰性對照(貨號133F0EC)、Trans G病毒感染液(貨號133F027),購自上海吉凱基因公司;miRNA 1st Strand cDNA Synthesis Kit(貨號MR101-02)、miRNA Universal SYBR qPCR Master Mix(貨號MQ101-02)、HiScript Ⅱ Q RT SuperMix for qPCR(貨號R222-01)、ChamQ Universal SYBR qPCR Master Mix(貨號Q711-02),購自南京諾唯贊生物科技有限公司;流式破膜固定劑(貨號554722)、Mouse CD4-FITC抗體(貨號553057)、Mouse IL-17A-PE抗體(貨號561020),購自美國BD公司;小鼠IL-17A高敏ELISA試劑盒(貨號EK217HS),購自聯科生物技術有限公司;丙酮酸測定試劑盒(貨號A081)、乳酸測定試劑盒(貨號A019-2-1),購自南京建成生物工程研究所。
1.1.5主要儀器 5920R臺式低溫離心機,購自德國Eppendorf公司;LV100N光學顯微鏡,購自日本NIKON公司;Micro 21R高速冷凍離心機、HFU 486超低溫冰箱,購自美國賽默飛世爾科技公司;KQ-500E型超聲波清洗器,購自昆山市超聲儀器有限公司;CO2培養箱,購自日本SANYO電氣有限公司;autoMACS Pro全自動磁珠分選儀,購自德國美天旎公司;M200Pro酶標儀,購自瑞士TECAN公司;7500型熒光定量PCR儀,購自美國ABI公司;Gallios分析型流式細胞儀,購自美國Beckman公司。
1.2 實驗方法
1.2.1RA患者臨床指標觀察及標本采集 根據RA診斷標準,對所有RA患者疾病活動評分(DAS28)進行評估;并于清晨空腹狀態下采集患者靜脈血5 mL,其中1 mL用于血沉(ESR)、類風濕因子(RF)和C反應蛋白(CRP)水平檢測,4 mL用于分離外周血CD4+T細胞。RA患者及健康志愿者的具體情況如Tab 1所示。

Tab 1 Characteristics of patients with RA and HCs
1.2.2人外周血CD4+T細胞分離 無菌收集HCs和RA患者外周抗凝血,采用Ficoll密度梯度離心法提取外周血單個核細胞(peripheral blood mononuclear cells,PBMCs);每107個細胞加入人CD4+T細胞分選磁珠50 μL,室溫避光孵育30 min,BD緩沖液補足液面至1 mL并立即置于BD Imagnet磁力架上,10 min后,吸棄上清,即獲得人CD4+T細胞,留作后續qPCR檢測。
1.2.3小鼠na?ve CD4+T細胞分離與體外Th17誘導分化 取C57BL/6小鼠脾臟制備單個核細胞懸液,照小鼠na?ve CD4+T細胞分選試劑盒說明依次加入MACS buffer(40 μL/ 107個細胞)、Biotin-Antibody Cocktail(10 μL/107個細胞)、Anti-Biotin磁珠(20 μL/107個細胞)、CD44磁珠(10 μL/107個細胞),4 ℃避光孵育10 min。打開MACS全自動磁珠分選儀,設置陰性選擇程序,將上述含免疫磁珠的細胞懸液加到LS柱中,收集柱下流出的液體,重復3次后離心(1 000 r·min-1,5 min),棄上清,用含10%胎牛血清的RPMI1640培養基重懸、計數,使得細胞濃度為(0.5-1)×109·L-1。
將小鼠na?ve CD4+T細胞分別以2×105細胞/每孔接種于24孔板,24孔板應提前包被濃度為2 mg·L-1的anti-CD3、anti-CD28,每孔中分別加入細胞因子IL-6、IL-23、TGF-β至終濃度為25、20、5 μg·L-1[9],同時進行相應的細胞實驗給藥處理,于37 ℃、5% CO2環境下繼續培養72 h。
1.2.4細胞實驗分組及處理 將CD4+T細胞分為Control組、miR-143-3p下調組、miR-143-3p上調組,每培養孔分別加入預先配制的陰性對照、miR-143-3p inhibitor、miR-143-3p mimics慢病毒感染復合物各100 μL,以誘導miR-143-3p的高表達/低表達;將CD4+T細胞分為Control組以及不同濃度的給藥組:CAT(20、40、80 mg·L-1),Control組給予相同體積1640培養基;將CD4+T細胞分為Control組、miR-143-3p下調組、miR-143-3p下調 + CAT處理組,于miR-143-3p inhibitor慢病毒感染復合物處理12 h后,miR-143-3p下調 + CAT處理組加入CAT至終濃度為40 mg·L-1,miR-143-3p下調組給予相同體積RPMI1640培養基。
1.2.5qPCR檢測miR-143-3p、Glut1、HK2、PKM2、LDHA、RORγt表達 使用TRIzol法提取細胞總RNA,將其反轉錄成cDNA,以此cDNA為模板擴增DNA,檢測miR-143-3p、Glut1、HK2、PKM2、LDHA、RORγt mRNA的相對表達量,以U6作為miR-143-3p內參,以GAPDH作為Glut1、HK2、PKM2、LDHA、RORγt內參,結果用2-ΔΔCt表示。引物根據Primer 5.0引物設計軟件設計,再由生工生物工程(上海)有限公司合成,引物序列如Tab 2所示。

Tab 2 Primers used for quantitative real-time PCR
1.2.6比色法檢測細胞培養上清丙酮酸、乳酸含量 收集細胞上清液及人外周血血清樣本,根據說明書進行操作后混勻,在分光光度計505 nm處檢測OD值,計算丙酮酸含量;在分光光度計530 nm處檢測OD值,計算乳酸含量。
1.2.7ELISA法檢測細胞培養上清IL-17A水平 收集細胞培養上清,嚴格按照小鼠IL-17A高敏ELISA試劑盒說明書操作,并分別于450和570 nm處檢測OD值,計算出對應的IL-17A濃度。
1.2.8流式細胞術檢測Th17細胞分化比例 收集體外誘導的Th17并用PBS洗滌1次,加入1 μLCD4-FITC流式抗體室溫避光孵育30 min,PBS清洗,加入1 mL破膜劑室溫避光孵育20 min,1 mL破膜洗液清洗,加入2 μL IL-17A-PE流式抗體室溫避光孵育40 min,破膜洗液清洗,加入500 μL PBS重懸細胞,置于流式細胞儀上檢測Th17分化比例。采用Flow Jo V10軟件分析并處理流式數據。
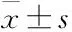
2 結果
2.1 RA患者miR-143-3p表達與糖酵解及Th17細胞分化的關聯性如Fig 1所示,與HC組比較,RA患者外周血CD4+T細胞中miR-143-3p表達明顯下降(P<0.05),Th17特異性轉錄因子RORγt mRNA表達明顯升高(P<0.05),Pearson′s分析結果表明,miR-143-3p表達與RORγt mRNA水平(P<0.05)、疾病嚴重度DAS28(P<0.01)具有相關性且均呈負相關;如Fig 2所示,與HC組比較,RA患者外周血CD4+T細胞中糖酵解關鍵酶Glut1、HK2、PKM2、LDHA mRNA表達均顯著升高(P<0.05);Pearson′s分析結果得出,miR-143-3p表達與Glut1、HK2、LDHA mRNA表達水平具有相關性且均呈負相關(P<0.05)。
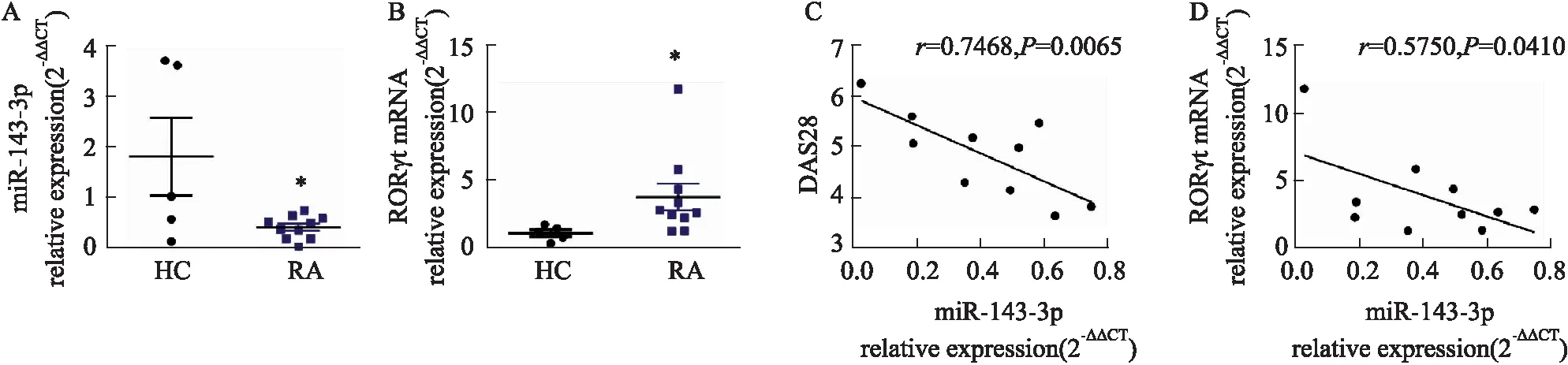
Fig 1 Correlation analysis of miR-143-3p and RORγt, DAS28 from RA patients HC: n=5; RA: n=10)(A-B) The relative expression of miR-143-3p and RORγt mRNA in peripheral blood CD4+T cells of RA patients and HCs was detected by RT-qPCR. *P<0.05 vs HC group by student′s t-test. (C) The correlation between miR-143-3p expression and DAS28 and RORγt mRNA expression. The Pearson correlation coefficient was determined.
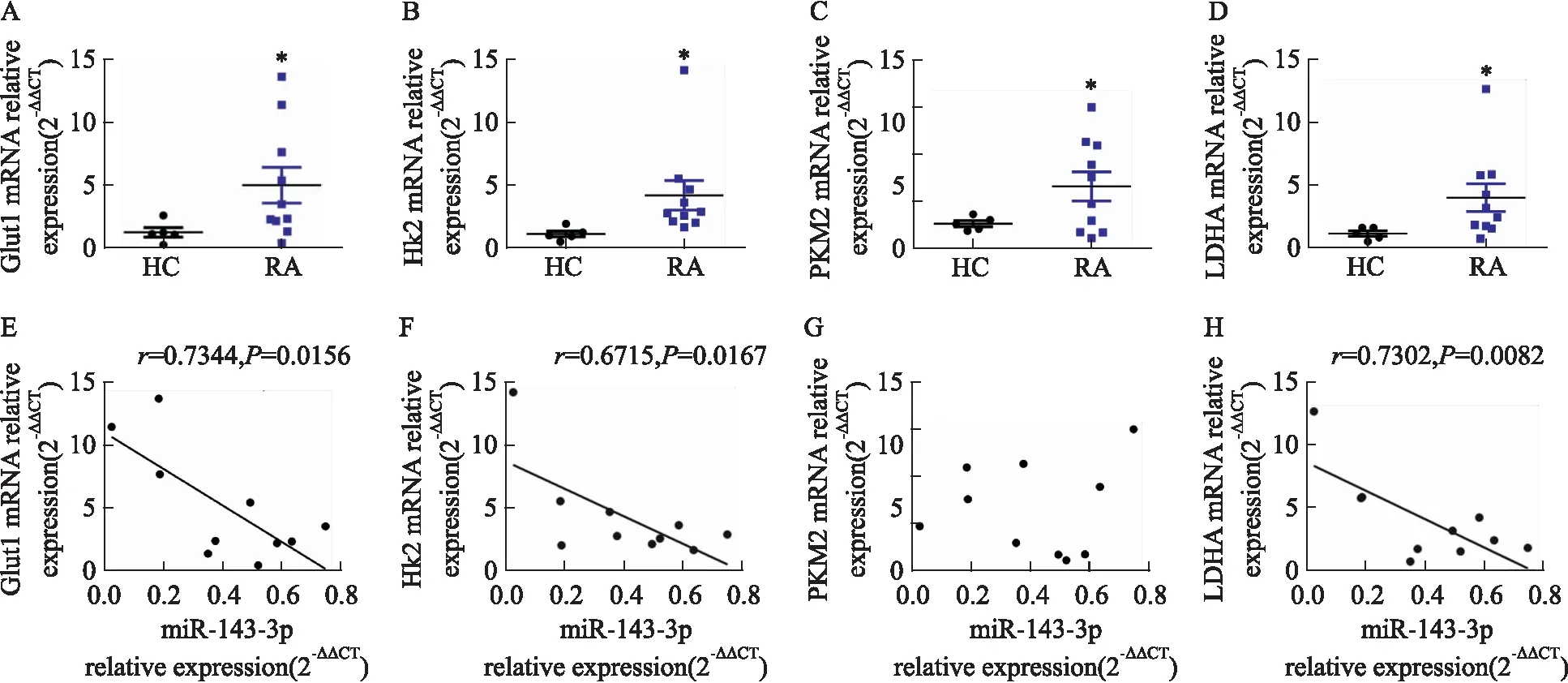
Fig 2 Correlation analysis of Glut1, HK2, PKM2, LDHA mRNA relative expression and miR-143-3p from RA patients HC: n=5; RA: n=10)(A-D) The mRNA relative expression of Glut1, HK2, PKM2 and LDHA in peripheral blood CD4+T cells of RA patients and HCs was detected by RT-qPCR. *P<0.05 vs HC group by student’s t-test. (E-H) The correlation between miR-143-3p expression and Glut1, HK2, PKM2, LDHA mRNA expression. The Pearson correlation coefficient was determined.
2.2 MiR-143-3p異常表達對Th17分化及細胞糖酵解的影響如Fig 3所示,Th17細胞轉染miR-143-3p inhibitor后,miR-143-3p表達較Control組明顯降低(P<0.05),轉染miR-143-3p mimics后,miR-143-3p表達較Control組明顯升高(P<0.01),表明轉染成功;如Fig 4所示,轉染miR-143-3p inhibitor后,RORγt mRNA水平及上清IL-17A水平較Control組明顯升高(P<0.05),轉染miR-143-3p mimics后,RORγt mRNA水平及上清IL-17A水平較Control組明顯降低(P<0.05),提示miR-143-3p能夠負調控Th17細胞分化及分泌活性;同時,如Fig 5所示,Th17細胞轉染miR-143-3p inhibitor后,Glut1、HK2、LDHA mRNA水平較Control組明顯升高(P<0.01),其細胞培養上清中丙酮酸水平也明顯升高(P<0.05);而在轉染miR-143-3p mimics后,HK2、LDHA mRNA水平較Control組明顯下降(P<0.01),其細胞培養上清中乳酸水平也明顯下降(P<0.05),提示miR-143-3p能夠負調控Th17的細胞糖酵解。

Fig 3 The mRNA relative expression of miR-143-3p in miR-143-3p mimics/inhibitor-transfected Th17 cells n=3)*P<0.05,**P<0.01 vs control group.
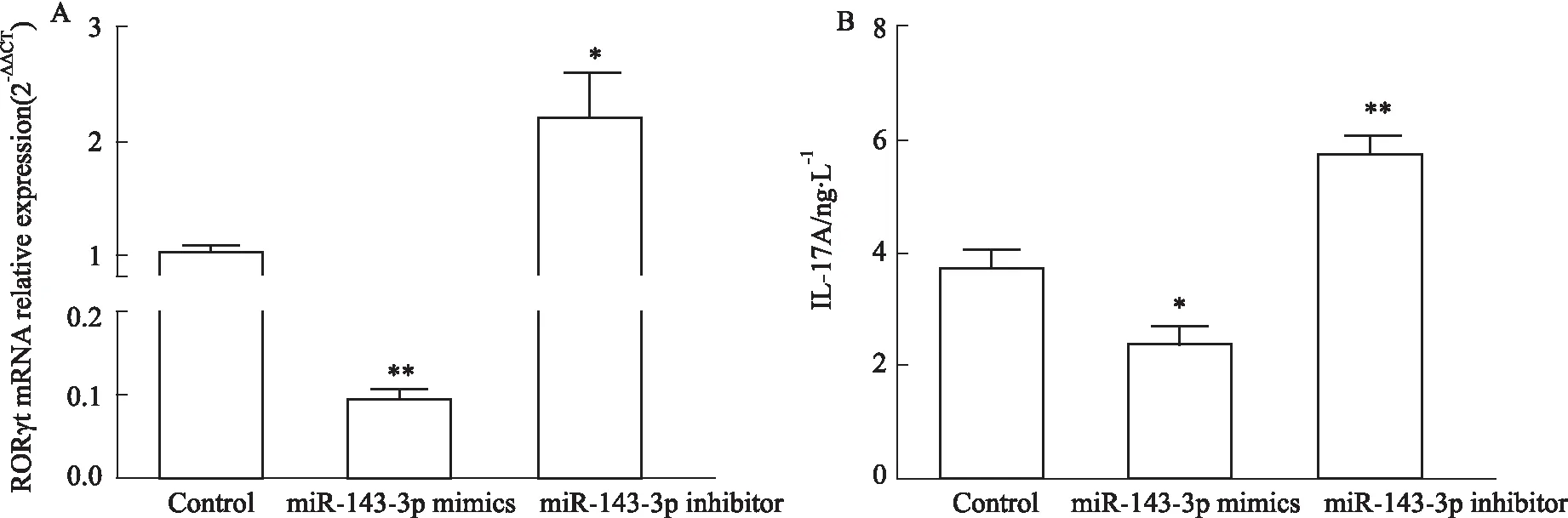
Fig 4 The mRNA relative expression of RORγt and level of IL-17A in miR-143-3p mimics/inhibitor-transfected Th17 cells n=3)CD4+T cells were transfected with miR-143-3p mimics or inhibitor for 12 h, then cultured with the condition of Th17-polarization for 72 h. (A) The mRNA relative expression of RORγt in CD4+T cells was detected by RT-qPCR. (B) The IL-17A levels in cell supernatant was determined by ELISA. *P<0.05,**P<0.01 vs control group by one-way ANOVA with Dunnett′s post-hoc test.

Fig 5 The mRNA relative expression of Glut1, HK2, LDHA and levels of pyruvate, lactate in miR-143-3p mimics/inhibitor-transfected Th17 cells n=3)CD4+T cells were transfected with miR-143-3p mimics or inhibitor for 12 h, then cultured with the condition of Th17-polarization for 72 h. (A) The mRNA relative expression of Glut1, HK2 and LDHA in CD4+T cells was detected by RT-qPCR. (B-C) The pyruvate and lactate levels in cell supernatant. *P<0.05,**P<0.01 vs control group by one-way ANOVA with Dunnett′s post-hoc test.
2.3 CAT對miR-143-3p表達和Th17細胞分化的影響如Fig 6所示,與Control組比較,CAT處理后Th17細胞分化比例、IL-17A分泌水平、RORγt mRNA表達水平均呈劑量依賴性下降(P<0.01),提示CAT可明顯抑制Th17細胞分化和分泌活性;而與Control組相比,CAT處理后miR-143-3p表達水平呈劑量依賴性上升(P<0.01),提示CAT可促進Th17細胞中miR-143-3p表達,見Fig 7。

Fig 6 Frequency of Th17 cells and level of IL-17A and RORγt mRNA relative expression after CAT treatment n=3)CD4+T cells (with the condition of Th17-polarization) were treated with CAT (20, 40, 80 mg·L-1) for 72h. (A-B) Representative dot plots and the percentage of Th17 (CD4+IL-17A+) cells was determined by flow cytometry. (C) The mRNA relative expression of RORγt in CD4+T cells was detected by RT-qPCR. (D) The IL-17A levels in cell supernatant was determined by ELISA. *P<0.05,**P<0.01 vs control group by one-way ANOVA with Dunnett's post-hoc test.
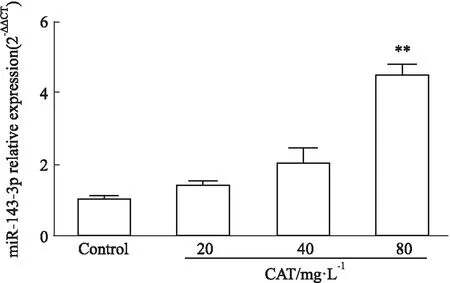
Fig 7 The mRNA relative expression of miR-143-3p after CAT treatment n=3)**P<0.01 vs control group.
2.4 CAT對Th17細胞糖酵解的影響如Fig 8所示,與Control組比較,CAT處理后糖酵解關鍵酶HK2、LDHA mRNA表達水平呈劑量依賴性降低(P<0.01),糖酵解代謝產物丙酮酸、乳酸也同樣隨劑量依賴性下降(P<0.05),提示CAT可顯著抑制Th17的細胞糖酵解。

Fig 8 The mRNA relative expression of Glut1, HK2, LDHA and levels of pyruvate, lactate after CAT treatment n=3)CD4+T cells (with the condition of Th17-polarization) were treated with CAT (20, 40, 80 mg·L-1) for 72h. (A) The mRNA relative expression of Glut1, HK2 and LDHA in CD4+T cells was detected by RT-qPCR. (B-C) The pyruvate and lactate levels in cell supernatant. *P<0.05,**P<0.01 vs control group by one-way ANOVA with Dunnett′s post-hoc test.
2.5 CAT對miR-143-3p低表達狀態下Th17細胞分化的干預作用如Fig 9所示,與Control組比較,miR-143-3p inhibitor組miR-143-3p表達明顯降低(P<0.05),提示轉染成功,而miR-143-3p inhibitor結合CAT給藥組較miR-143-3p inhibitor組miR-143-3p表達明顯升高(P<0.01),提示CAT可恢復Th17細胞中miR-143-3p的被抑制狀態;同時,與miR-143-3p inhibitor組比較,miR-143-3p inhibitor結合CAT給藥組Th17細胞分化比例、IL-17A分泌水平、RORγt mRNA水平均明顯降低(P<0.01),并恢復至與Control組同等的水平(Fig 10),提示CAT可明顯抑制miR-143-3p低表達狀態下Th17細胞的異常分化。
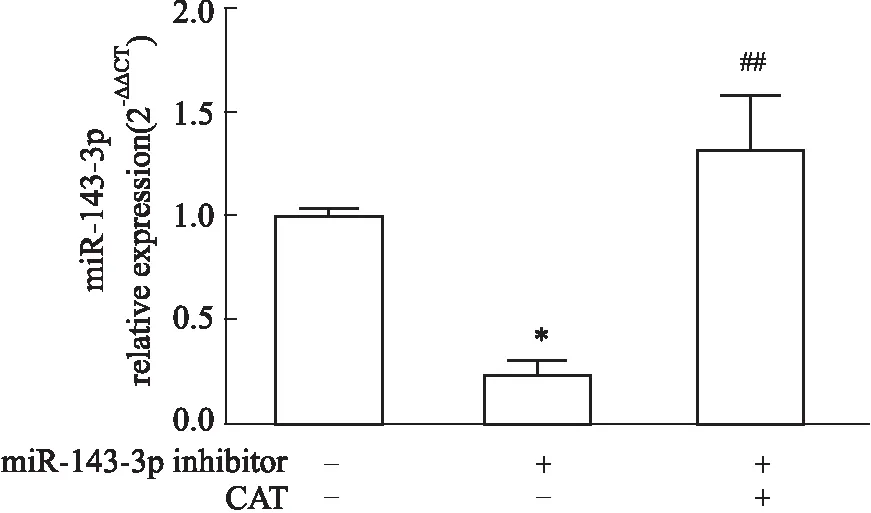
Fig 9 The mRNA relative expression of miR-143-3p in miR-143-3p low expression-Th17 cells after CAT treatment n=3)*P<0.05 vs control group; ##P<0. 01 vs miR-143-3p inhibitor group.

Fig 10 Frequency of Th17 cells and level of IL-17A and RORγt mRNA relative expression in miR-143-3p low expression-Th17 cells after CAT treatment n=3)CD4+T cells were transfected with miR-143-3p inhibitor for 12 h, then treated with CAT (40 mg·L-1) under the condition of Th17-polarization for 72 h. (A-B) Representative dot plots and the percentage of Th17 (CD4+IL-17A+) cells was determined by flow cytometry. (C) The mRNA relative expression of RORγt in CD4+T cells was detected by RT-qPCR. (D) The IL-17A levels in cell supernatant was determined by ELISA. **P<0.01 vs control group; ##P<0.01 vs miR-143-3p inhibitor group by one-way ANOVA with Dunnett′s post-hoc test.
2.6 CAT對miR-143-3p低表達狀態下Th17細胞糖酵解的干預作用進一步驗證CAT調控miR-143-3p對Th17分化的干預作用是否與細胞糖酵解有關。如Fig 11所示,與miR-143-3p inhibitor組比較,miR-143-3p inhibitor結合CAT給藥組HK2、LDHA mRNA表達水平(P<0.01),代謝產物丙酮酸、乳酸水平均明顯降低(P<0.05),其抑制程度與control組類似,提示CAT可恢復miR-143-3p低表達狀態下Th17細胞的糖酵解異常上調。
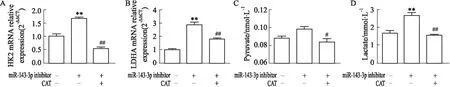
Fig 11 The mRNA relative expression of HK2, LDHA and levels of pyruvate, lactate in miR-143-3p low expression-Th17 cells after CAT treatment n=3)CD4+T cells were transfected with miR-143-3p inhibitor for 12 h, then treated with CAT (40 mg·L-1) under the condition of Th17-polarization for 72h. (A-B) The mRNA relative expression of HK2 and LDHA in CD4+T cells was detected by RT-qPCR. (C-D) The pyruvate and lactate levels in cell supernatant. **P<0.01 vs control group; ##P<0. 01 vs miR-143-3p inhibitor group by one-way ANOVA with Dunnett′s post-hoc test.
3 討論
Th17細胞作為CD4+T細胞中特異性分泌IL-17的細胞亞群,不僅可以清除細胞外細菌及霉菌,也可介導機體炎癥反應、自身免疫及移植排斥反應。RA是目前常見的自身免疫性疾病,大量研究表明Th17細胞的異常分化可影響RA的疾病進程,因此,調控Th17細胞的過度分化是RA免疫治療的研究熱點。前期項目組研究已發現,CAT可明顯下調膠原誘導關節炎(CIA)小鼠中Th17細胞的分化及分泌活性,有效緩解小鼠關節炎癥的進展[10]。本研究結果同樣顯示,生地中主要活性成分CAT可在體外顯著抑制Th17細胞分化比例,下調Th17細胞特
征性轉錄因子RORγt mRNA水平,減少Th17細胞特征性細胞因子IL-17A分泌,進一步證明CAT可顯著抑制Th17細胞的分化和功能。但CAT調控Th17細胞分化的作用機制目前尚不明確。
miRNA是一種保守的內源性非編碼RNA,其調控作用幾乎涉及包括細胞增殖分化、細胞間信號通路傳導、自噬、凋亡在內的所有生物學過程,是CD4+T細胞分化與效應功能發揮的重要調節劑[11]。項目組前期針對RA患者、CIA小鼠外周血miRNAs表達進行檢測,共篩選出包括miR-143-3p在內的6種miRNAs存在表達失調[12],其中miR-143-3p作為一種典型的多功能microRNA,已被證實在多種自身免疫性疾病及炎癥模型(RA、慢性結腸炎、膠原誘導小鼠關節炎模型等)中表達出現異常下調[13-15]。同時,多項研究顯示,miR-143-3p可抑制Th17細胞分化所需的IL-6、IL-23分泌[16]。結合項目組前期發現CAT可顯著促進CD4+T細胞中miR-143-3p表達,本實驗進一步嘗試研究CAT通過miR-143-3p調控Th17細胞分化的作用機制。
本研究結果提示,與健康對照組比較,RA患者CD4+T細胞中miR-143-3p表達明顯下降,Th17特異性轉錄因子RORγt mRNA表達明顯升高,且miR-143-3p表達與RORγt水平、疾病嚴重度DAS28均呈負相關,提示RA患者CD4+T細胞中存在miR-143-3p的表達異常,這可能是RA中Th17異常分化、疾病持續發展的關鍵因素之一;進一步建立體外誘導Th17細胞分化模型,并結合慢病毒轉染誘導miR-143-3p的高表達/低表達,發現過表達miR-143-3p后,RORγt mRNA水平及IL-17A分泌水平均顯著降低,反之則顯著升高,表明miR-143-3p可負調控Th17細胞分化及分泌活性,可成為Th17介導的RA疾病治療的有效靶點。因此,本實驗繼續在miR-143-3p低表達細胞模型的基礎上,結合CAT給藥,以進一步觀察CAT對miR-143-3p抑制狀態下Th17細胞分化的調控作用。實驗結果顯示,與miR-143-3p inhibitor組比較,miR-143-3p inhibitor結合CAT給藥組miR-143-3p的表達明顯上調,表明CAT可明顯逆轉Th17細胞分化過程中miR-143-3p的異常下調。同時,與miR-143-3p inhibitor組比較,miR-143-3p inhibitor結合CAT給藥組Th17細胞分化比例、IL-17A分泌水平、RORγt表達水平均明顯降低,表明CAT通過誘導miR-143-3p表達來抑制CD4+T細胞向Th17細胞亞群分化。然而,miR-143-3p潛在的調節范圍仍不確定,一種miRNA往往可以與多種基因相互作用,一種基因也往往含有多個miRNA識別位點。因此,接下來本實驗重點關注miR-143-3p是通過何種途徑調控Th17細胞的分化和功能。
多項研究表明,RA疾病進展與病灶處CD4+T細胞的代謝變化特點密切相關,由于RA關節滑膜腔是一個相對缺氧的局部微環境,導致了CD4+T細胞從氧化磷酸化向糖酵解的代謝轉換,而糖酵解的異常上調則可誘導CD4+T細胞向致炎性的Th17細胞亞群的持續分化,加重滑膜炎癥[17]。miRNAs可通過復雜的機制調控細胞代謝過程,包括直接靶向代謝過程的關鍵分子(轉運體或酶/激酶)和調節多種信號通路[5]。而miR-143-3p作為調控抑癌基因的非編碼小RNA,已被發現能夠直接靶向HK2的轉錄表達,抑制腫瘤細胞糖酵解,減緩其增殖能力[18]。結合CAT對糖脂代謝的調控作用,推測CAT調控miR-143-3p對Th17細胞分化的干預作用是否與糖酵解有關。
本研究結果提示,RA患者外周血CD4+T細胞中糖酵解關鍵酶Glut1、HK2、LDHA mRNA表達顯著上調,且與miR-143-3p呈負相關,提示RA患者CD4+T細胞中的糖酵解失調可能與miR-143-3p有關;慢病毒轉染誘導Th17細胞體外分化過程中miR-143-3p的高表達/低表達,發現抑制miR-143-3p后,Glut1、HK2、LDHA mRNA水平及丙酮酸、乳酸分泌較Control組顯著升高,反之則明顯降低,表明miR-143-3p可以負調控Th17細胞糖酵解。結合該調控機制的研究結果,研究進一步觀察了CAT對Th17細胞體外分化過程中糖酵解的影響,結果顯示,CAT處理后Th17細胞中糖酵解酶HK2、LDHA的mRNA水平以及代謝產物丙酮酸、乳酸的分泌水平均呈劑量依賴性下降,表明CAT可顯著下調Th17分化過程中的細胞糖酵解。結合慢病毒轉染誘導miR-143-3p的低表達,發現與miR-143-3p inhibitor組比較,miR-143-3p inhibitor結合CAT給藥組HK2、LDHA mRNA水平,丙酮酸、乳酸分泌水平均明顯降低,表明CAT可恢復miR-143-3p低表達狀態下的糖酵解異常上調和Th17細胞異常分化。
綜上所述,本研究證實miR-143-3p可負調控Th17細胞分化及分泌活性,可成為Th17介導的RA疾病治療的有效靶點,且miR-143-3p對Th17細胞的調控作用可能與其糖酵解有關。CAT可上調miR-143-3p,同時抑制Th17細胞的異常分化和糖酵解異常上調,表明CAT可通過調控miR-143-3p,抑制細胞糖酵解并干預Th17的異常分化,這可能是生地調節機體免疫平衡,發揮抗炎作用的重要機制之一,這為生地在RA等自身免疫病中的合理應用提供了實驗基礎。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
學苑創造·A版(2020年9期)2020-10-13 09:41:02
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
學苑創造·A版(2015年11期)2016-01-14 09:03:27
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
七彩語文·畫刊(2012年4期)2012-04-29 00:00:00
