前列腺素E2 促進損傷腸黏膜修復的研究進展
2022-07-25 06:44:06樂夢真范漢程朱清仙曾慧紅邵立健
新醫(yī)學 2022年7期
樂夢真 范漢程 朱清仙 曾慧紅 邵立健
腸道黏膜上皮處于不斷的再生-凋亡循環(huán),更新周期通常為3~5 d。腸隱窩底部的腸道干細胞(ISC)具有自我更新與多向分化為各種腸道上皮細胞(IEC)的潛能,當腸黏膜上皮受到損傷時,這種潛能對于維持腸黏膜屏障完整性至關重要。另外,當受炎癥和輻射等刺激時,ISC 及IEC 受到損傷,若存活的ISC 不足以補充缺失的IEC,腸黏膜屏障將會受損,從而導致各種消化道疾病。腸黏膜損傷情況下,體內(nèi)多種物質(zhì)參與促進腸黏膜損傷的修復,如前列腺素E(PGE)、IL-6、胰島素樣生長因子-1(IGF-1)、血管心外膜活性物質(zhì)(BVES)等。這些物質(zhì)促進黏膜修復機制及激活的信號通路不盡相同,但作用的共同點都是通過促進腸隱窩底部的ISC 增殖分化,從而促進損傷腸黏膜的修復。但在此修復過程中,如果相關通路的某些關鍵基因發(fā)生突變,將導致ISC 惡性增殖乃至腸道腫瘤的發(fā)生。因此,ISC 的生長調(diào)控是近年來腸道疾病防治的研究熱點。隨著PGE對成體干細胞(包括ISC 等)作用的深入研究, PGE促進損傷腸黏膜修復方面的研究也有了明顯的進展,由于篇幅限制,本文主要就PGE在病理條件下對腸道黏膜損傷的修復作用及其對ISC 功能的影響做一綜述。
一、PGE2 的合成及其受體在腸道中的分布
PGE可由多種類型細胞經(jīng)一系列酶促反應產(chǎn)生,細胞膜上的膜磷脂在磷脂酶A2(PLA2)的作用下釋放出花生四烯酸(AA)。在環(huán)氧合酶(COX)催化下,AA 被氧化生成前列腺素G(PGG)和前列腺素H(PGH), PGH作為前列腺素合成的中間產(chǎn)物,在特定前列腺素E 合成酶(PGES) 作用下生成PGE(圖1)。
COX 作為PGE形成過程中的關鍵酶,有兩種同工酶:COX-1 和COX-2,但這兩種同工酶在腸道的表達及其作用有較大的差異。Xiao 等(2017年)在小鼠實驗中發(fā)現(xiàn),正常情況COX-1 在包括ISC 在內(nèi)的所有隱窩細胞中持續(xù)性表達,COX-1 介導的PGE在腸腔內(nèi)發(fā)揮保護腸黏膜屏障、維持腸上皮完整性的作用,使腸黏膜在日常生活中免受胃酸、藥物等刺激,保證腸黏膜上皮持續(xù)更新。傳統(tǒng)NSAID 如阿司匹林由于抑制COX-2 的同時也抑制了COX-1,使COX-1 對腸黏膜的保護作用受到破壞,從而增加了潰瘍等各種胃腸道不良反應的發(fā)生風險。COX-2 在正常腸道細胞內(nèi)表達很少,主要在炎癥和腫瘤等病理刺激下呈誘導性表達。COX-2 在炎癥和輻射中能促進損傷腸黏膜的修復,而在腫瘤中則促進其發(fā)生發(fā)展。以上研究結果表明在生理和病理條件下,表達COX-1 或COX-2 在腸黏膜損傷修復中起重要作用。
PGE在體內(nèi)發(fā)揮作用是通過與前列腺素E 受體(EP)結合實現(xiàn)。EP 受體屬G 蛋白偶聯(lián)受體(GPCR),有EP1、EP2、EP3 和EP4 四種。EP1 和EP2 可以分別激活磷脂酶C(PLC)和環(huán)磷酸腺苷(cAMP),導致正信號轉導,激活相關信號通路,促進細胞增殖;EP3 則負向調(diào)節(jié)cAMP 水平,起到相反的作用;而EP4 可以激活cAMP 和磷脂酰肌醇3 激酶(PI3K)兩條途徑,激活相關信號通路,促進細胞增殖 (圖1)。各類型EP 受體在腸道中的表達水平有所不同,正常情況下,小鼠結腸固有層細胞表達所有EP 受體,隱窩細胞只表達EP2和EP4,而小鼠小腸表達低水平EP 受體。近期筆者團隊也證明了小鼠腸隱窩細胞表達EP2 和EP4,并證實了氟尿嘧啶作用下腸隱窩細胞內(nèi)的EP2 表達升高,但EP4 的表達未受影響。在炎癥等刺激下,小腸隱窩細胞可高表達EP2 和EP4,PGE與其結合可促進隱窩細胞增殖并修復損傷的腸黏膜。但病理條件下,小腸隱窩細胞EP 受體表達的作用未知,ISC 是否表達EP 受體及其意義值得探討。
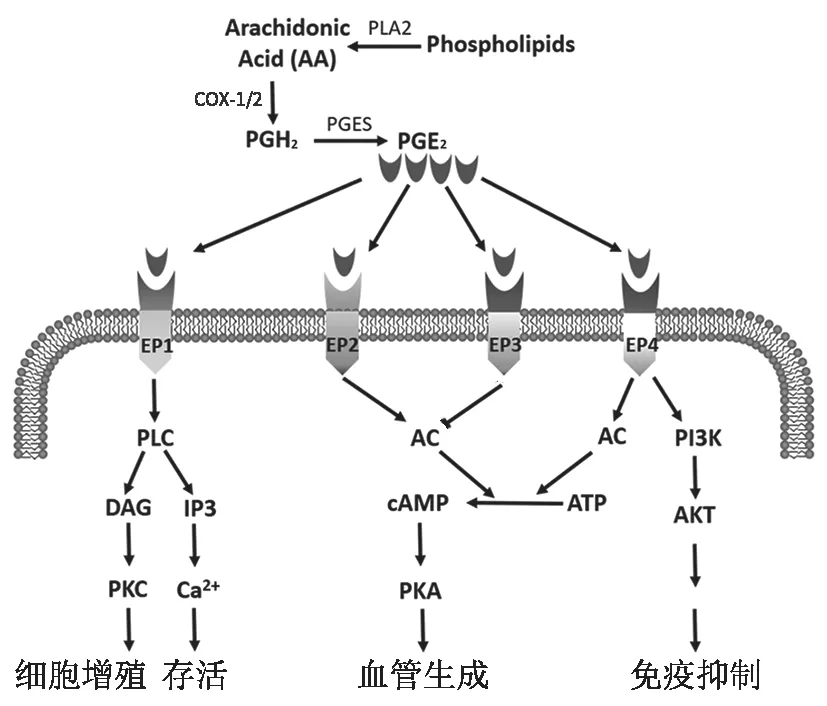
圖1 PGE2 的合成及胞內(nèi)相關信號通路
二、腸黏膜結構及其形成的調(diào)節(jié)因素
正常腸黏膜由腸絨毛與腸隱窩構成,腸隱窩底部的ISC 能夠自我更新與分化為多種成熟腸道細胞,構成絨毛與隱窩結構的細胞支架。在ISC 增殖分化過程中,ISC 先增殖產(chǎn)生各種早期祖細胞-短暫擴增細胞(TA),緊接著TA 細胞快速分裂,并沿腸隱窩-絨毛軸縱向遷移與定向分化為各種成熟腸道細胞,吸收細胞、杯狀細胞和內(nèi)分泌細胞向上遷移,構成絨毛結構,而潘氏細胞則向下遷移至隱窩底部,與ISC、TA 細胞共同構成腸隱窩結構(圖2)。哺乳動物腸隱窩內(nèi)有兩種不同類型的ISC,一類位于隱窩底部,其分布于潘氏細胞之間,更新迅速,能被富含亮氨酸重復單位的G蛋白偶聯(lián)受體5(Lgr5)、嗅素4(Olfm4)和無剛毛-盾片樣蛋白2(Ascl2)等標記,主要受Wnt信號通路調(diào)節(jié),對輻射刺激敏感,稱為活性干細胞(A-ISC)或隱窩底部柱狀細胞或Lgr5ISC(另有說法為Lgr5ISC 位于結腸隱窩底部,小腸隱窩+4 區(qū));另一類位于隱窩底部潘氏細胞上方的+4區(qū),其更新緩慢,能夠被B 淋巴細胞特異的莫洛尼鼠白血病病毒插入位點-1(Bmi1)、多亮氨酸重復區(qū)免疫球蛋白樣蛋白1(Lrig1)、唯同源異型結構域蛋白X(Hopx)和小鼠端粒酶逆轉錄酶(mTert)等標記,主要由骨形態(tài)發(fā)生蛋白(BMP)信號通路調(diào)節(jié),Wnt 信號通路處于抑制狀態(tài),有輻射抵抗作用,稱為靜止干細胞(Q-ISC) 或標記滯留細胞。除以上標記物外,近年來發(fā)現(xiàn)有A-ISC 與Q-ISC 共表達標記物,如干細胞RNA 結合蛋白Musashi1 和Mex-3 同源蛋白A(Mex-3A)等。在生理情況或腸黏膜損傷較輕時,腸黏膜自我更新與修復主要由Lgr5ISC 介導,在腸黏膜受到嚴重損傷,存活的Lgr5ISC 不足以維持腸黏膜完整性時,一部分Q-ISC 可以轉化為Lgr5ISC,以促進腸道細胞再生與損傷黏膜的修復。
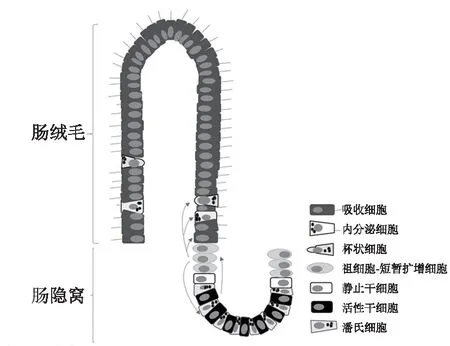
圖2 ISC 在隱窩底部的分布及分化
生理情況下,腸黏膜上皮完整性主要由ISC維持,而ISC 的增殖分化受多種信號通路的精確調(diào)控,主要有Wnt/β-連環(huán)素(β-catenin)、BMP和Notch 等信號通路。其中,Wnt 信號通路是調(diào)控A-ISC 生長增殖的主要通路,而BMP、Notch通路主要調(diào)控TA 分化。Wnt 通路參與了多種腸道疾病的發(fā)生發(fā)展,且在腸黏膜損傷的修復中發(fā)揮主要作用,因此了解Wnt 通路調(diào)節(jié)機制對于研究損傷腸黏膜修復具有重要意義。隱窩周圍肌成纖維細胞分泌R-脊椎蛋白1(R-spondin1),隱窩底部潘氏細胞分泌Wnt3,這些蛋白與相應受體結合后激活Wnt 信號通路,穩(wěn)定細胞內(nèi)β-catenin,β-catenin 通過激活T 淋巴細胞因子4(TCF4)促進細胞增殖。腸隱窩內(nèi)一種軸蛋白Axin,可以結合β-catenin、糖原合成酶激酶-3β(GSK-3β)與結腸腺瘤息肉蛋白(APC)形成大分子復合物,通過影響β-catenin 水平對Wnt 通路進行負性調(diào)節(jié)。GSK-3β 和APC 與Axin 結合后促進β-catenin磷酸化,從復合物脫離并降解,GSK-3β 也可以磷酸化APC,暴露APC 上的β-catenin 結合位點,使APC 與β-catenin 親和力增加,更加有利于β-catenin 降解,以此負性調(diào)節(jié)Wnt 信號通路,抑制Wnt/β-catenin 促細胞增殖作用。正常情況下,Wnt/β-catenin 信號通路的正性與負性調(diào)控處于動態(tài)平衡,保證ISC 的生理性增殖,從而維持腸黏膜的正常結構。如負性調(diào)控通路中某些基因突變或缺失,β-catenin 堆積,將導致Wnt 信號通路過度激活,使ISC 增長失控,形成腸道腫瘤。因此,調(diào)控Wnt 信號通路為研究腸黏膜損傷修復與腸道腫瘤靶向治療的關鍵點。
三、PGE2 促進損傷腸黏膜修復及其機制
完整的腸黏膜屏障保證機體消化道功能處于正常狀態(tài)。在慢性炎癥、藥物、輻射以及微生物入侵等各種刺激下,腸黏膜遭到破壞,導致腸黏膜表面潰瘍形成等病理變化,腸黏膜屏障的保護功能受到削弱,使機體產(chǎn)生腹瀉和便血等消化道癥狀。因此,緩解此類臨床癥狀的關鍵在于促進損傷腸黏膜的修復。
Hanson 等(1983 年)就證實了PGE可以促進照射損傷后的腸上皮增殖,但由于當時實驗條件限制,PGE促進腸上皮增殖的具體機制并不清楚。近年來,隨著科技的進步及實驗技術的提高,對PGE促進損傷腸黏膜修復的研究有了很大進展。由于炎癥性腸病(IBD)與放射、化學治療導致的胃腸道綜合征(RIGS)在臨床中極為常見,因此在相關實驗中,以藥物誘導的腸道炎癥及放射治療誘導的腸黏膜損傷模型最為常見。
在既往研究中,Singer 等(1998 年)發(fā)現(xiàn)IBD患者腸組織內(nèi)COX-2 和PGE水平明顯升高,PGE作為一種炎癥介質(zhì),參與IBD 炎癥反應。因此臨床多采用NSAID 來抑制COX,減少PGE產(chǎn)生,緩解腸道炎癥。臨床中發(fā)現(xiàn),長期使用NSAID 胃腸道會出現(xiàn)潰瘍和出血等不良反應,導致IBD 遷延不愈甚至病情惡化。近些年,人們對NSAID 導致腸黏膜損傷的機制進行了研究,發(fā)現(xiàn)NSAID 對腸黏膜的破壞與其抑制PGE產(chǎn)生有關。Lai 等(2015 年)在實驗中發(fā)現(xiàn)用瑞巴匹特和阿司匹林共同處理小鼠后,阿司匹林導致的小腸損傷明顯減輕,瑞巴匹特保護腸黏膜的這種作用與其誘導腸細胞內(nèi)COX-2 表達和PGE產(chǎn)生有關。有研究顯示PGE通過腸黏膜上皮的EP4 抑制上皮壞死性凋亡并誘導結腸炎的消退。另外,在其他方式導致的小鼠實驗性腸炎實驗中,Hang 等(2015 年)、Chen 等(2015 年)發(fā)現(xiàn),抑制PGE降解酶減少PGE代謝或使用藥物促進PGE產(chǎn)生后,腸道炎癥及黏膜損傷得到緩解;在穩(wěn)態(tài)條件下,PGE信號通路對ISC 增殖至關重要(如Lgr5干細胞),并誘導干細胞向腸細胞分化。以上研究均表明,PGE除作為炎癥因子參與腸道炎癥反應外,還能夠促進損傷的腸道黏膜修復,其可能機制為PGE與其EP 受體結合激活Wnt/β-catenin 信號通路,促進腸隱窩內(nèi)TA 與ISC 增殖分化,從而減少上皮細胞凋亡,修復黏膜損傷。此機制的發(fā)現(xiàn),為IBD的治療提供了新的藥物研究方向。
放射治療對機體造成的損傷復雜多樣,其中輻射誘導的RIGS 是臨床放射治療最常見的不良反應,由于放射治療對機體的損傷機制還不清楚,因此目前仍缺乏有效緩解RIGS 的藥物。近些年,電離輻射(IR)導致腸黏膜損傷的研究有了明顯進展。目前的研究認為,IR 導致ISC(尤其是Lgr5ISC)的凋亡是RIGS 的始發(fā)因素。筆者之前的研究表明外源性PGE促進化學治療所致的腸損傷的恢復。這是通過16, 16-二甲基前列腺素E(dmPGE)治療促進ISC 的增殖和分化實現(xiàn)的。Shao 等(2008 年)、Elizabeth 等(2012 年)以及Parthasarathy 等(2016 年)發(fā)現(xiàn),照射會誘導小鼠體內(nèi)PGE含量升高,而升高的PGE增加了隱窩內(nèi)增殖細胞數(shù)量與促進了腸黏膜的修復;有研究報道外源性脂肪來源的間充質(zhì)干細胞(MSC)對腸道的放射損傷也有治療作用,其通過激活 COX-2-PGE信號軸進行修復。Chen 等(2015 年)發(fā)現(xiàn),用PGE類似物dmPGE處理照射后的小鼠,發(fā)現(xiàn)腸隱窩內(nèi)增殖期細胞增多,照射誘導的腸道細胞凋亡緩解;用縮宮素處理照射小鼠后,小鼠存活率升高,腸道細胞凋亡率下降,且縮宮素的這種作用與其促進COX-2 的表達與PGE生成有關。PGE可以通過激活Wnt/β-catenin 信號通路促進損傷腸黏膜修復。Miyoshi 等(2017 年)在對結腸鏡檢導致小鼠腸黏膜損傷的模型中也得以驗證。以上研究提示,PGE可以緩解輻射導致的腸黏膜損傷,其機制與其促進炎癥損傷腸黏膜修復機制相似,PGE通過與其EP 受體結合,激活Wnt/β-catenin 信號通路,使隱窩內(nèi)處于增殖期的TA 與ISC 增多,從而促進上皮細胞更新,修復損傷黏膜。由于導致IRGS 的因素主要為IR 導致Lgr5ISC損傷與凋亡,那么PGE修復IRGS 的機制是否與Lgr5ISC 直接相關?通過查閱以上文獻采用的研究方法,筆者發(fā)現(xiàn)上述實驗是用5-溴脫氧尿嘧啶核苷(BrdU)或增殖細胞核抗原(PCNA)標記增殖細胞的方式判定PGE對增殖細胞的作用,以及提取腸組織或腸隱窩中的相關蛋白或RNA 來探究其作用機制,然而這種方法并不能直接證明PGE是通過作用于Lgr5ISC 而發(fā)揮腸黏膜損傷的修復作用。因此,ISC 是否表達EP 受體?ISC 表達哪類EP 受體?PGE是否直接結合ISC 表面EP 受體來促進腸黏膜修復?這些問題值得深入探索。
四、PGE2/Wnt 通路和ISC 靶向治療現(xiàn)況與前景
PGE作為炎癥因子與促細胞增殖物質(zhì),在腸黏膜損傷的修復中起著雙刃劍的作用。一方面,抑制PGE產(chǎn)生,有助于減輕炎癥,而過度或長期減少PGE產(chǎn)生,將削弱PGE促進ISC 增殖的作用,影響腸黏膜更新,破壞腸黏膜完整性。另一方面,在克羅恩病患者來源的小腸中,TNF-α 導致 Lgr5干細胞功能障礙,外源性 PGE治療恢復了 Lgr5干細胞的功能,而PGE大量產(chǎn)生,將使Wnt 通路過度激活,導致ISC 增殖失控,如PGE/Wnt 通路中關鍵基因PlA2g2a(編碼PLA2)、APC(編碼APC)等發(fā)生突變,將促使ISC 轉化為腫瘤干細胞,導致腸道腫瘤的發(fā)生。且對于某些可以使體內(nèi)PGE升高的腫瘤,如宮頸癌的放射性治療中,過度使用PGE緩解RIGS,將降低該類腫瘤放射治療的敏感性,削弱放射治療療效。因此,探究PGE最佳給藥劑量、時間及Wnt 信號通路最佳激活程度,對于修復損傷腸黏膜及最大化減少其帶來的不良反應尤為重要。
干細胞治療一直是近年來研究的熱點,MSC具有多向分化能力和免疫調(diào)節(jié)功能,對于治療IBD方面具有獨特的優(yōu)勢。隨著干細胞研究的興起,有研究用MSC 移植的方式來治療實驗性腸炎,并發(fā)現(xiàn)PGE/Wnt 通路在其中發(fā)揮了重要作用,MSC以自分泌的方式分泌PGE,通過激活Wnt 信號通路促進自身增殖,以此促進腸上皮更新。ISC 移植能否成為新的干細胞治療研究方向?Yui 等用腔內(nèi)移植方式將ISC 移植到小鼠體內(nèi),發(fā)現(xiàn)被移植的ISC 能夠很好地在腸上皮組織中增殖分化,參與構建正常的隱窩結構并覆蓋葡聚糖硫酸鈉(DSS)誘導腸炎中的潰瘍損傷面,ISC 移植治療中PGE/Wnt 通路是否在其中發(fā)揮重要作用,這為將來ISC 移植治療的深入研究提供了重要的思路。
雖然ISC 移植治療對于腸黏膜損傷具有直接修復作用,但也有文獻指出,MSC 移植用于治療RIGS 時,MSC 分泌的PGE可能會促進原有腫瘤生長或降低腫瘤放射治療敏感性,因此,干細胞治療緩解RIGS 具有一定的局限性。對此,針對Wnt 信號通路的分子靶向治療已成為近年的研究熱點。Zhou 等發(fā)現(xiàn),用Wnt 激動劑R-spondin 1 和其在Lgr5ISC 上的受體Slit2 蛋白共同處理照射損傷的小鼠后,輻射誘導的ISC 凋亡明顯減少,而且小鼠DSS 誘導的多發(fā)性腸腺瘤放射治療敏感性并未下降。Bhanja 等發(fā)現(xiàn),一種分子制劑BCN057可以激活Wnt /β-catenin 信號通路,促進Lgr5ISC再生,從而緩解RIGS,且人結腸癌組織與小鼠腹部皮下腫瘤放射治療敏感性并未受到影響。以上研究提示,通過激活Wnt 信號通路的相關治療,可以在不影響腫瘤放射化學治療敏感性的前提下,促進ISC 增殖,緩解RIGS。但相關實驗仍停留在動物實驗或體外實驗階段,要使其投入臨床正式使用,還需要大量動物及臨床實驗對其可行性及可能帶來的不良反應進行探索。
干細胞治療作為新興研究方向,在腸道疾病的治療中有著非常重要的臨床意義,是未來腸道疾病防治研究的趨勢。但干細胞治療對相關實驗技術要求較高,ISC 體外培養(yǎng)方案還不十分完善,目前很多設想還停留在細胞或動物實驗階段,臨床可行性還不確定。而PGE是經(jīng)FDA 批準可用于臨床的藥物,經(jīng)過長期臨床應用,人們對PGE相關作用機制及不良反應已經(jīng)有了一定的認識,因此,探究PGE在修復腸黏膜損傷中的最佳給藥劑量與時間有著更為實際的研究意義。
五、結 語
大量研究證明,PGE可以激活Wnt/β-catenin信號通路促進TA/ISC 增殖,從而修復各種刺激造成的腸黏膜損傷,促進腸黏膜屏障重建。因此,通過調(diào)節(jié)PGE水平及Wnt 活性,調(diào)控ISC 增殖分化,也許能預防和緩解甚至治療腸黏膜損傷相關疾病。但還有許多未知的科學問題值得深入研究,如ISC是否表達EP 受體?表達哪些EP 受體?PGE是否直接作用ISC 表面EP 受體?PGE長時程作用于ISC 的利弊?PGE長時程應用和腸道腫瘤關系如何?PGE與ISC 表面EP 受體結合后除激活Wnt信號通路外,其他調(diào)節(jié)ISC 病理生理的信號通路(如Notch 和BMP 等)有何變化?這些問題的解決將為正確使用PGE提供依據(jù),同時為聯(lián)合其他藥物改善腸黏膜損傷提供理論基礎。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
學苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
中國生殖健康(2019年3期)2019-02-01 06:12:26
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
云南中醫(yī)學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
