bta-miR-34b/c和bta-miR-449a/b/c靶基因預(yù)測(cè)及生物信息學(xué)分析
2022-07-27 06:47:24杜鑫澤馬鑫浩張殿琦杜嘉偉謝琨成昝林森
中國(guó)畜牧獸醫(yī) 2022年7期
關(guān)鍵詞:小鼠
杜鑫澤,馬鑫浩,張殿琦,杜嘉偉,馬 婧,謝琨成,何 杰,昝林森,2
(1.西北農(nóng)林科技大學(xué)動(dòng)物科技學(xué)院,楊凌 712100;2.國(guó)家肉牛改良中心,楊凌 712100;3.西安市奶牛育種中心,西安 710000)
在雄性哺乳動(dòng)物中,精子發(fā)生是一個(gè)復(fù)雜且受多種途徑精密調(diào)控的生理過(guò)程,主要經(jīng)歷4個(gè)階段:第一階段,A0型精原細(xì)胞有絲分裂經(jīng)歷A1-A4精原細(xì)胞、中間型精原細(xì)胞、B型精原細(xì)胞并最終形成初級(jí)精母細(xì)胞;第二階段,初級(jí)精母細(xì)胞開(kāi)始第一次減數(shù)分裂形成次級(jí)精母細(xì)胞;第三階段,初級(jí)精母細(xì)胞進(jìn)行第二次減數(shù)分裂形成圓形精子細(xì)胞;第四階段,圓形精子細(xì)胞經(jīng)過(guò)一系列變形分化為成熟精子[1]。在精子發(fā)生的整個(gè)過(guò)程中,雄性生殖細(xì)胞分化過(guò)程具有復(fù)雜的轉(zhuǎn)錄調(diào)控機(jī)制,除了編碼蛋白質(zhì)的mRNA外,許多非編碼RNA如微小RNA(microRNA,miRNA)、piwi相互作用RNA(PIWI interacting RNA,piRNA)、環(huán)狀RNA(circularRNA,circRNA)、長(zhǎng)鏈非編碼RNA(long noncoding RNA,lncRNA)等都發(fā)揮著重要的轉(zhuǎn)錄后調(diào)控作用。
miRNA是一類(lèi)進(jìn)化上高度保守的長(zhǎng)度在22 nt左右的小分子非編碼RNA,miRNA可以參與調(diào)控包括性成熟、睪丸發(fā)育、精子發(fā)生在內(nèi)的諸多生物學(xué)過(guò)程,其在精子發(fā)生過(guò)程中以細(xì)胞特異性或階段特異性方式表達(dá)[2]。靶向mRNA的3′-UTR以實(shí)現(xiàn)基因沉默是miRNA的經(jīng)典作用機(jī)制[3],miRNA常通過(guò)與靶向mRNA的3′-UTR中的互補(bǔ)序列結(jié)合,并通過(guò)與多蛋白R(shí)NA誘導(dǎo)的沉默復(fù)合物(RNA-induced silencing complex,RISC)相互作用來(lái)加速轉(zhuǎn)錄物衰減或翻譯抑制來(lái)抑制基因表達(dá)[4]。miRNA在眾多生物學(xué)途徑中的作用突出了轉(zhuǎn)錄后基因調(diào)控的復(fù)雜性。研究表明,miRNA的表達(dá)具有較強(qiáng)的組織特異性,表明miRNA在不同組織、不同生物學(xué)過(guò)程中具有獨(dú)特的功能。miRNA因其在基因表達(dá)的轉(zhuǎn)錄后調(diào)控中占據(jù)主導(dǎo)地位而被深入研究并廣泛應(yīng)用于諸多領(lǐng)域[5]。就畜牧生產(chǎn)而言,種公畜的繁殖性能被育種學(xué)家高度關(guān)注,因此,探究miRNA對(duì)家畜精子發(fā)生的影響和作用機(jī)制成為了農(nóng)業(yè)科學(xué)研究的熱點(diǎn)之一[6]。
miR-34家族由3個(gè)成員組成,分別是miR-34a、miR-34b和miR-34c。miR-34b/c在精子發(fā)生過(guò)程中具有多種功能,既可以調(diào)節(jié)減數(shù)分裂過(guò)程,也可以調(diào)節(jié)精子發(fā)生后期的變形和獲能等生物學(xué)過(guò)程。研究證明,多種動(dòng)物的精子中含有miR-34b/c,這些成員在精子發(fā)生[7]和早期胚胎發(fā)育[8]中發(fā)揮著重要作用。miR-449家族也由3個(gè)成員組成,分別是miR-449a、miR-449b和miR-449c,它們被定位到Cdc20b基因的第2個(gè)內(nèi)含子上,形成一個(gè)miRNA簇,在不同物種間高度保守。miR-449成員在控制細(xì)胞周期和表皮分化中起主要作用[9]。報(bào)道顯示,miR-34/449家族在纖毛發(fā)生[10]、細(xì)胞周期調(diào)節(jié)[11]、細(xì)胞分化[12]、癌癥[13]等多種生物學(xué)過(guò)程中發(fā)揮作用。雖然miR-34和miR-449最初被指定為單獨(dú)的miR家族,但miR-34與miR-449家族成員具有相同的種子序列,且miR-34和miR-449被證明可以調(diào)節(jié)一組相同的靶基因,因此可以被視為一個(gè)miRNA簇,命名為miR-34/449[14]。鑒于這2個(gè)家族成員在基因表達(dá)調(diào)控中可能一起發(fā)揮作用,且2個(gè)基因家族之間具有相互代償機(jī)制,本研究利用生物信息學(xué)方法探究miR-34b/c、miR-449a/b/c在牛精子發(fā)生中的作用機(jī)制及調(diào)控網(wǎng)絡(luò),通過(guò)預(yù)測(cè)共同調(diào)控miR-34b/c、miR-449a/b/c的轉(zhuǎn)錄因子(transcription factor,TF)以及受miR-34b/c、miR-449a/b/c共同調(diào)控的靶基因以構(gòu)建miR-34b/c、miR-449a/b/c的分子調(diào)控網(wǎng)絡(luò),以期為后續(xù)miR-34b/c和miR-449a/b/c調(diào)控牛精子發(fā)生的相關(guān)研究提供理論基礎(chǔ)。
1 材料與方法
1.1 材料
牛(Bostaurus,bta)、人(Homosapiens,hsa)、小鼠(Musmusculus,mmu)、大鼠(Rattusnorvegicus,rno)、雞(Gallusgallus,gga)和豬(Susscrofa,ssc)等物種的miR-34b/c、miR-449a/b/c及其前體的序列信息來(lái)源于 miRBase(http:∥mirbase.org/index.shtml)在線(xiàn)數(shù)據(jù)庫(kù)。
1.2 方法
1.2.1 miR-34b/c和miR-449a/b/c的相似性分析 利用MegaX[15]軟件對(duì)miR-34b/c和miR-449a/b/c前體序列分別進(jìn)行多序列比對(duì),并計(jì)算各模型組合的BIC分?jǐn)?shù)(Bayesian information criterion),選取BIC分?jǐn)?shù)最低的模型組合,即最大似然估計(jì)法(maximum likelihood)、Jukes-Cantor模型和Gamma分布分別構(gòu)建miR-34b/c和miR-449a/b/c的系統(tǒng)進(jìn)化樹(shù)。
1.2.2 bta-miR34/449靶基因預(yù)測(cè) 通過(guò)在線(xiàn)軟件TargetScan 7.2(http:∥www.targetscan.org/vert_72/)[16]、miRDB(http:∥www.mirdb.org/)[17]和miRWalk(http:∥mirwalk.umm.uni-heidelberg.de/)[18]預(yù)測(cè)bta-miR-34b/c、bta-miR-449a/b/c的靶基因,取三者預(yù)測(cè)結(jié)果的交集作為bta-miR34/449的靶基因。
1.2.3 靶基因功能預(yù)測(cè) 根據(jù)靶基因預(yù)測(cè)結(jié)果,選取bta-miR-34b/c和bta-miR-449a/b/c共同作用的靶基因。分別使用在線(xiàn)軟件DAVID 5.8(https:∥david.ncifcrf.gov/)[19]和KOBAS(http:∥kobas.cbi.pku.edu.cn/)[20]對(duì)其進(jìn)行GO功能和KEGG通路富集分析(物種為牛),收集調(diào)整后P<0.05的條目并使用R語(yǔ)言程序包ggplot2繪制柱狀圖和氣泡圖。
1.2.4 轉(zhuǎn)錄因子預(yù)測(cè) 在NCBI數(shù)據(jù)庫(kù)中檢索出bta-miR-34b/c和bta-miR-449a/b/c的前體序列,使用AnimalTFDB(http:∥bioinfo.life.hust.edu.cn/AnimalTFDB/#!/)[21]對(duì)其上游2 kb至上游0.1 kb的序列進(jìn)行轉(zhuǎn)錄因子結(jié)合位點(diǎn)預(yù)測(cè),獲取可能調(diào)控bta-miR-34/449的轉(zhuǎn)錄因子。
1.2.5 構(gòu)建TF-miRNA-靶基因作用網(wǎng)絡(luò) 選取同時(shí)調(diào)控bta-miR-34b/c和bta-miR-449a/b/c的轉(zhuǎn)錄因子,結(jié)合靶基因預(yù)測(cè)和String在線(xiàn)數(shù)據(jù)庫(kù)(http:∥string-db.org/)[22]構(gòu)建蛋白互作網(wǎng)絡(luò),使用Cytoscape 3.8.0在線(xiàn)軟件(http:∥www.cytoscape.org/)[23]構(gòu)建TF-miR-34/449-mRNA作用網(wǎng)絡(luò)圖。
2 結(jié) 果
2.1 miR-34/499的相似性分析
根據(jù)miRBase數(shù)據(jù)庫(kù)的查詢(xún)結(jié)果,牛miR-34/499分別位于第15和20號(hào)染色體上,對(duì)比牛、人、小鼠、大鼠、雞和豬等物種的miR-34/449序列并構(gòu)建系統(tǒng)進(jìn)化樹(shù),結(jié)合序列比對(duì)發(fā)現(xiàn),miR-34/449在進(jìn)化中相對(duì)保守,成熟體序列基本一致(表1、2,圖1)。
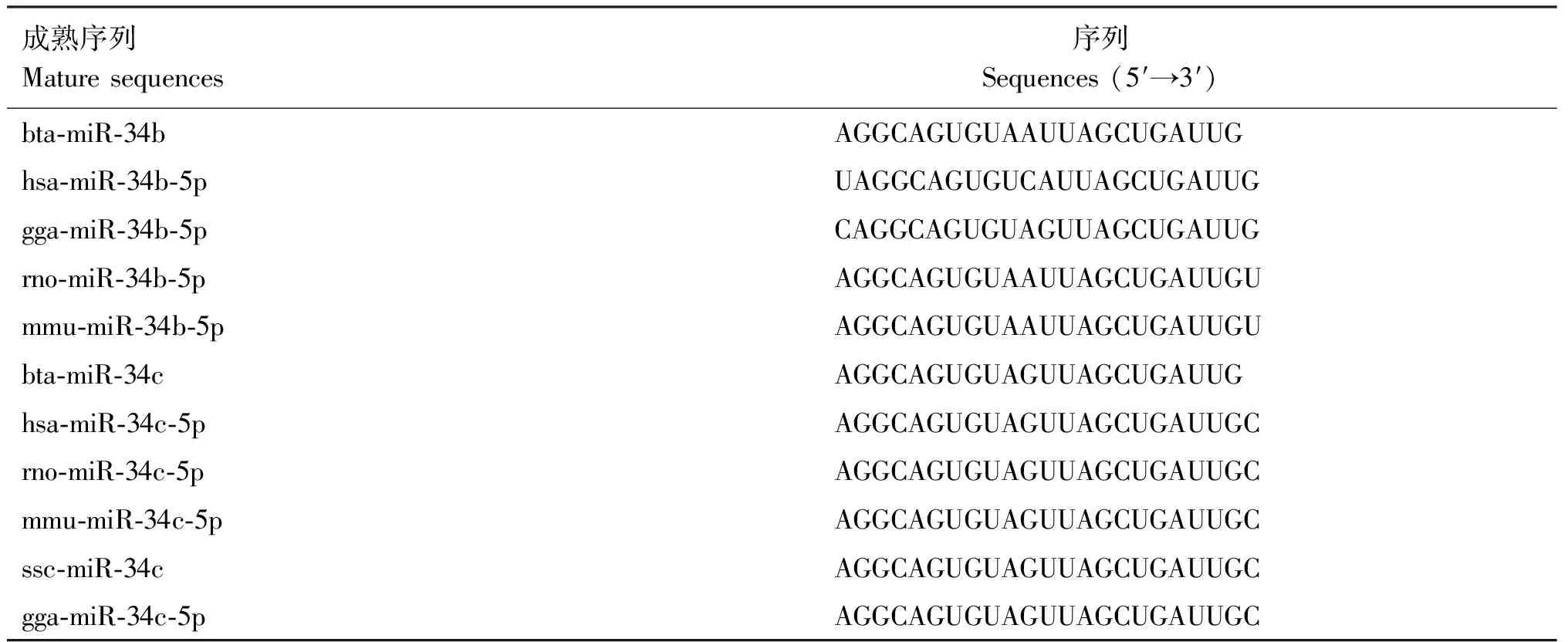
表1 各物種miR-34b/c的成熟序列
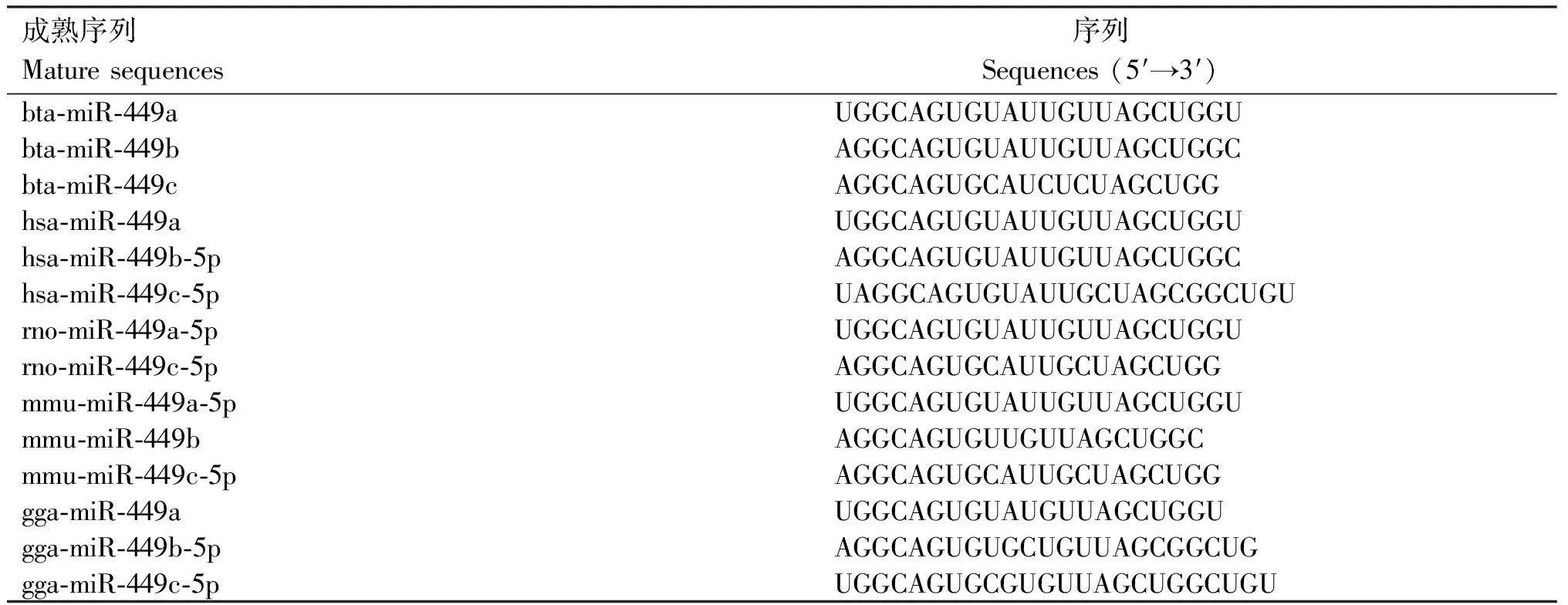
表2 各物種miR-449a/b/c的成熟序列
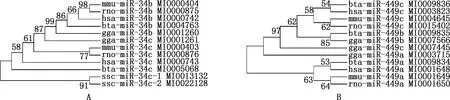
圖1 ML法構(gòu)建miR-34b/c(A)、miR-449a/b/c(B)系統(tǒng)進(jìn)化樹(shù)
2.2 靶基因預(yù)測(cè)
通過(guò)在線(xiàn)軟件TargetScan、miRDB和miRWalk對(duì)bta-miR-34b/c、bta-miR-449a/b/c的靶基因進(jìn)行預(yù)測(cè),取三者的交集分別得到了115、151、161、149和152個(gè)靶基因(圖2)。

A,bta-miR-34b;B,bta-miR-34c;C,bta-miR-449a;D,bta-miR-449b;E,bta-miR-449c
2.3 靶基因功能預(yù)測(cè)
bta-miR-34b/c和bta-miR-449a/b/c共同作用的靶基因分別為73和80個(gè),用于功能分析的靶基因?yàn)?9個(gè)(圖3、表3),如SYT1、SNX15、LGI1、ELMOD1、RCAN1等。對(duì)靶基因集進(jìn)行GO功能和KEGG通路富集分析,結(jié)果顯示,bta-miR-34b/c和bta-miR-449a/b/c的靶基因主要富集在精子發(fā)生、鈣離子依賴(lài)性胞吐的調(diào)節(jié)、胰島素分泌的調(diào)節(jié)等GO條目,以及HIF-1信號(hào)通路、甲狀腺激素信號(hào)通路、Wnt信號(hào)通路等KEGG通路(圖4)。

A,bta-miR-34b∩bta-miR-34c;B,bta-miR-449a∩bta-miR-449b∩bta-miR-449c;C,bta-miR-34b/c∩bta-miR-449a/b/c。圖5同

表3 bta-miR-34b/c和bta-miR-449a/b/c的共同靶基因

圖4 靶基因的GO功能(A)和KEGG通路(B)富集分析
2.4 轉(zhuǎn)錄因子預(yù)測(cè)
預(yù)測(cè)到bta-miR-34b和bta-miR-34c共同的轉(zhuǎn)錄因子有77個(gè),bta-miR-449a、bta-miR-449b和bta-miR-449c共同的轉(zhuǎn)錄因子有17個(gè),以上5個(gè)miRNAs共同的轉(zhuǎn)錄因子有2個(gè)(圖5、表4),分別為核受體亞家族2C組成員2(nuclear receptor subfamily 2C group member 2,NR2C2)和HICZBTB轉(zhuǎn)錄抑制因子1(HIC ZBTB transcriptional repressor 1,HIC1)。
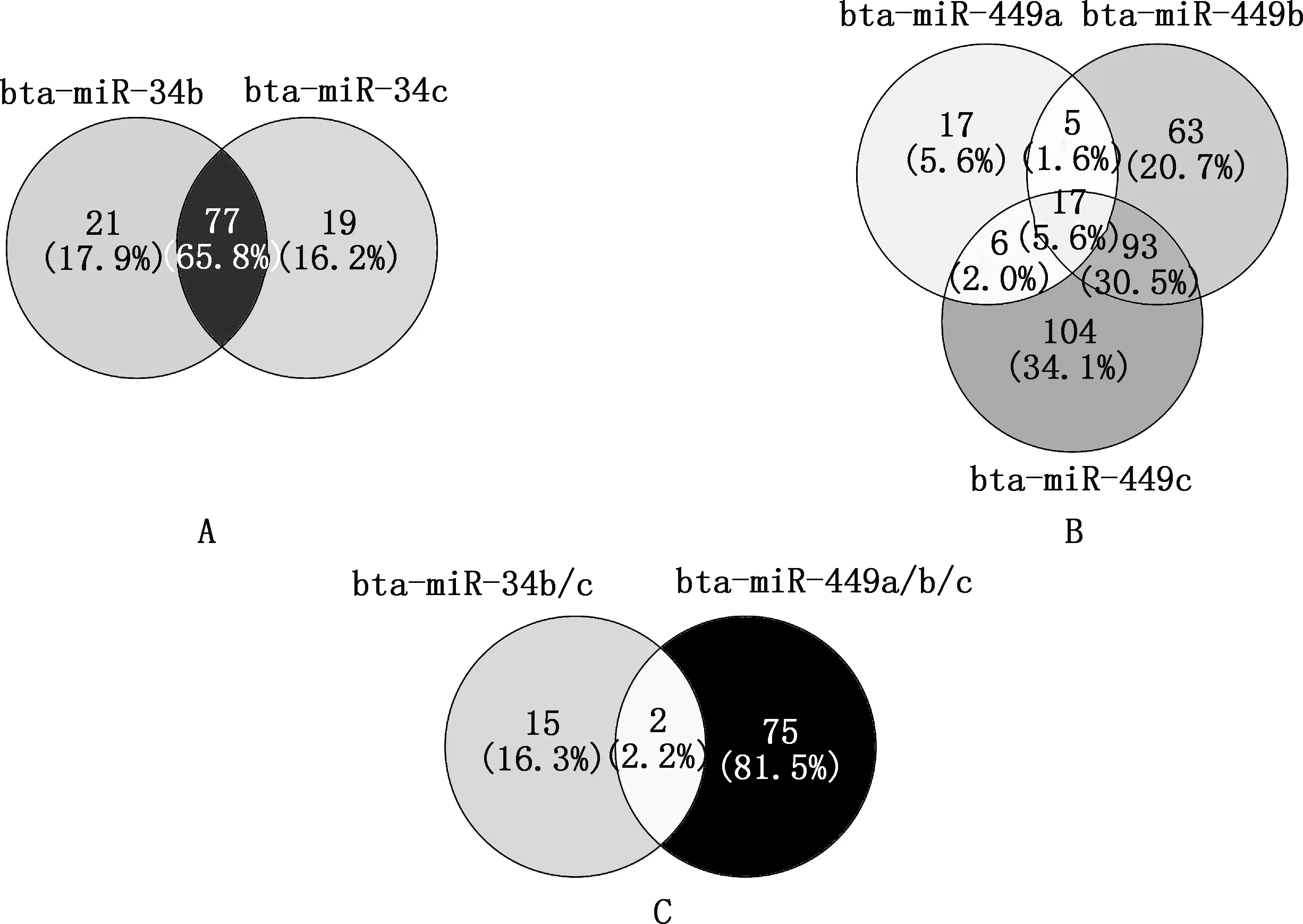
圖5 bta-miR-34b/c和bta-miR-449a/b/c的轉(zhuǎn)錄因子預(yù)測(cè)

表4 bta-miR-34b/c和bta-miR-449a/b/c的轉(zhuǎn)錄因子
2.5 TF-miRNA-mRNA作用網(wǎng)絡(luò)的構(gòu)建
根據(jù)bta-miR-34/449預(yù)測(cè)的轉(zhuǎn)錄因子、靶基因以及蛋白間互作關(guān)系構(gòu)建TF-miRNA-mRNA作用網(wǎng)絡(luò),結(jié)果顯示,bta-miR-34b/c、bta-miR-449a/b/c對(duì)精子發(fā)生的調(diào)控作用通過(guò)其與上游轉(zhuǎn)錄因子、下游靶基因共同構(gòu)成的TF-miRNA-mRNA作用網(wǎng)絡(luò)發(fā)揮作用,bta-miR-34b/c、bta-miR-449a/b/c受到上游轉(zhuǎn)錄因子NR2C2和HIC1的調(diào)控,同時(shí)會(huì)抑制AXL、E2F3、DAAM1等下游靶基因的表達(dá),以實(shí)現(xiàn)對(duì)精子發(fā)生的生物學(xué)調(diào)控(圖6)。

圖6 TF-bta-miR-34b/c、bta-miR-449a/b/c-mRNA作用網(wǎng)絡(luò)
3 討 論
在基因表達(dá)過(guò)程中,miRNA能夠通過(guò)轉(zhuǎn)錄后調(diào)控影響基因的表達(dá),miRNA是基因表達(dá)的關(guān)鍵調(diào)節(jié)因子,有望成為生物標(biāo)志物開(kāi)發(fā)的候選分子。單個(gè)miRNA可以靶向數(shù)百個(gè)mRNA并影響許多基因的表達(dá)[24]。精子發(fā)生過(guò)程是雄性動(dòng)物得以產(chǎn)生優(yōu)質(zhì)精子的重要生物學(xué)過(guò)程,miRNA已被證明參與精子發(fā)生的各個(gè)時(shí)期[25]。miR-34家族可以靶向許多細(xì)胞周期調(diào)節(jié)因子,如Notch1、CDK4和MYC,且TGIF2和Notch2在精子發(fā)生過(guò)程中是miR-34c的2個(gè)重要靶基因,Notch信號(hào)能夠促進(jìn)生殖細(xì)胞的分化[26]。有報(bào)道稱(chēng),miR-34c在精母細(xì)胞和圓形精子細(xì)胞中高表達(dá),抑制原代精母細(xì)胞中的miR-34c可以防止生殖細(xì)胞免受睪酮?jiǎng)儕Z誘導(dǎo)的細(xì)胞凋亡,在體外培養(yǎng)的生殖細(xì)胞中miR-34c的過(guò)度表達(dá)會(huì)觸發(fā)細(xì)胞凋亡[27-28]。miR-449家族成員也在精子發(fā)生中扮演著重要角色,miR-449a、miR-449b和miR-449c在減數(shù)分裂開(kāi)始期間上調(diào),并優(yōu)先在精母細(xì)胞和精子細(xì)胞中表達(dá)[29]。大量研究已經(jīng)揭示了miRNA-34和miR-449家族對(duì)精子發(fā)生各個(gè)時(shí)期的調(diào)控作用及機(jī)制,但其對(duì)精子發(fā)生過(guò)程中上游轉(zhuǎn)錄因子和下游靶基因構(gòu)成的基因表達(dá)調(diào)控網(wǎng)絡(luò)還有待進(jìn)一步挖掘。
本研究通過(guò)查詢(xún)miRBase在線(xiàn)數(shù)據(jù)庫(kù)中牛、人、小鼠、大鼠等物種的miR-34b/c、miR-449a/b/c及其前體序列信息,運(yùn)用相似性分析以及miRNA轉(zhuǎn)錄因子、靶基因預(yù)測(cè)、富集分析等技術(shù)手段對(duì)bta-miR-34b/c、bta-miR-449a/b/c的序列保守性、上下游調(diào)控機(jī)制及功能進(jìn)行分析,得到了49個(gè)共同靶基因和2個(gè)轉(zhuǎn)錄因子,其中,靶基因和轉(zhuǎn)錄因子的篩選通過(guò)在線(xiàn)數(shù)據(jù)庫(kù)TargetScan、miRDB、miRWalk及AnimalTFDB實(shí)現(xiàn),以上數(shù)據(jù)庫(kù)均包含牛的相關(guān)信息或支持通過(guò)序列直接預(yù)測(cè),盡管miRNA在物種間具有較強(qiáng)的序列保守性,但是使用其他物種miRNA的相關(guān)信息預(yù)測(cè)牛miRNA的靶基因或轉(zhuǎn)錄因子仍然不是最優(yōu)選擇,因此本試驗(yàn)通過(guò)對(duì)預(yù)測(cè)工具的合理選擇在一定程度上提高了研究結(jié)果的準(zhǔn)確性。
本研究預(yù)測(cè)到的2個(gè)轉(zhuǎn)錄因子(NR2C2和HIC1)在精子發(fā)生中發(fā)揮著重要作用。NR2C2也被稱(chēng)為睪丸核受體4(testicular orphan receptor 4,TR4),是核受體家族的成員[30]。在小鼠睪丸中,NR2C2主要表達(dá)于初級(jí)精母細(xì)胞和圓形精子細(xì)胞中,并在精子發(fā)生的減數(shù)分裂前期和連續(xù)減數(shù)分裂前期起重要作用。NR2C2基因敲除小鼠的生殖潛能顯著降低,各個(gè)階段的精子數(shù)量明顯降低,初級(jí)精母細(xì)胞降解,生精小管壞死扭曲,精子發(fā)生減數(shù)分裂前期和連續(xù)減數(shù)分裂期延長(zhǎng)和中斷,導(dǎo)致細(xì)胞分裂中期延長(zhǎng)和生精細(xì)胞異常[31]。HIC1是一種鋅指轉(zhuǎn)錄抑制因子,可通過(guò)與沉默交配型信息調(diào)節(jié)2同源物1(sirtuin 1,SIRT1)啟動(dòng)子結(jié)合,控制細(xì)胞生長(zhǎng)和死亡,以響應(yīng)p53依賴(lài)性凋亡DNA損傷[32]。Jenal等[33]研究發(fā)現(xiàn),HIC1是細(xì)胞周期和凋亡調(diào)節(jié)因子E2F轉(zhuǎn)錄因子1(E2F transcription factor 1,E2F1)的新轉(zhuǎn)錄靶點(diǎn),E2F1通過(guò)包含HIC1 P0啟動(dòng)子上的TATA盒在內(nèi)的2個(gè)E2F DNA結(jié)合位點(diǎn)誘導(dǎo)HIC1的表達(dá)。HIC1與包含HIC1結(jié)合共有位點(diǎn)的E2F1啟動(dòng)子基因結(jié)合,通過(guò)調(diào)控E2F1轉(zhuǎn)錄抑制細(xì)胞生長(zhǎng)。Uchida等[34]研究證明,HIC1缺失促進(jìn)青春期前睪丸平滑肌細(xì)胞的增殖,導(dǎo)致成年睪丸中生精小管周?chē)腜MC數(shù)量增加,并最終影響精子發(fā)生。
本研究篩選到的部分靶基因已有報(bào)道顯示參與調(diào)控了精子發(fā)生過(guò)程。受體酪氨酸激酶AXL是TAM家族的成員,在細(xì)胞增殖、分化、凋亡和新陳代謝等生物學(xué)過(guò)程中發(fā)揮著重要作用[35]。AXL是牛睪丸支持細(xì)胞中miR-34c的直接靶基因,miRNA-34c通過(guò)靶向AXL基因調(diào)節(jié)牛支持細(xì)胞增殖和凋亡[36]。E2F3是E2F家族中的一員,是唯一被發(fā)現(xiàn)在精原細(xì)胞、細(xì)線(xiàn)期精母細(xì)胞和支持細(xì)胞中表達(dá)的E2F成員。在支持細(xì)胞特異性視網(wǎng)膜母細(xì)胞瘤蛋白(RB)敲除小鼠睪丸內(nèi)注射shRNA-E2F3,結(jié)果顯示,支持細(xì)胞中因RB缺失引起的生精功能障礙通過(guò)E2F3的體內(nèi)沉默得到部分恢復(fù)[37],暗示RB/E2F3途徑在精子發(fā)生過(guò)程中發(fā)揮重要調(diào)節(jié)作用。形態(tài)發(fā)生相關(guān)激活因子1(dishevelled associated activator of morphogenesis 1,DAAM1)是一種formin家族蛋白,通過(guò)Wnt信號(hào)通路參與非支鏈肌動(dòng)蛋白絲的成核、形態(tài)發(fā)生和器官發(fā)生等重要生物學(xué)過(guò)程[38]。Pariante等[39]評(píng)估了DAAM1與大鼠睪丸形態(tài)發(fā)生的可能關(guān)聯(lián),結(jié)果顯示,在有絲分裂階段,DAAM1與支持細(xì)胞、生殖細(xì)胞和精原細(xì)胞中的肌動(dòng)蛋白共享其定位,在減數(shù)分裂期間,2種蛋白都存在于精母細(xì)胞中,而在形成的血睪丸屏障中只能檢測(cè)到肌動(dòng)蛋白,DAAM1在整個(gè)精子發(fā)生過(guò)程中跟隨頂體系統(tǒng)的發(fā)展,最終保留在成熟精子的細(xì)胞質(zhì)內(nèi)。乳酸脫氫酶A(lactate dehydrogenase A,LDHA)在精子能量代謝過(guò)程中起重要作用,熱應(yīng)激能夠增加豬睪丸中葡萄糖轉(zhuǎn)運(yùn)蛋白3(glucose transporter type 3,GLUT3)和LDHA的表達(dá),繼而增加支持細(xì)胞中乳酸產(chǎn)生[40]。小鼠中乳酸脫氫酶C(lactate dehydrogenase C,LDHC)基因的缺失導(dǎo)致精子無(wú)法獲能并在精卵結(jié)合中不能穿過(guò)透明帶[41],而LDHA作為轉(zhuǎn)基因引入LDHC缺失小鼠中,轉(zhuǎn)基因小鼠獲得了正常的精子功能和生育能力[42]。Klinefelter綜合征(Klinefelter syndrome,KS)是男性性腺機(jī)能減退和性染色體異常的遺傳疾病,患有KS的男性睪丸中LDHA mRNA的表達(dá)量減少與睪丸內(nèi)乳酸含量的下降相關(guān)[40]。以上證據(jù)不同程度地印證了本研究預(yù)測(cè)的靶基因在精子發(fā)生中扮演著重要角色。此外,本研究預(yù)測(cè)得到的還沒(méi)有被廣泛報(bào)道的其他基因也具有進(jìn)一步研究的價(jià)值。
靶基因GO功能富集分析表明,bta-miR-34b/c和bta-miR-449a/b/c的靶基因主要富集在精子發(fā)生、鈣離子依賴(lài)性胞吐的調(diào)節(jié)、胰島素分泌的調(diào)節(jié)等GO條目,推測(cè)bta-miR-34b/c和bta-miR-449a/b/c的靶基因在精子發(fā)生和精子獲能中起作用。相關(guān)報(bào)道顯示,miR-34c參與小鼠睪丸精子發(fā)生和雄性生殖細(xì)胞(male germ-line stem cells,mGSCs)凋亡的調(diào)節(jié)[7]。miR-34b/c和miR-449a/b/c對(duì)于正常精子發(fā)生和男性生育不可或缺[43]。miR-34b和miR-34c存在于與透明帶結(jié)合的精子中,表明其可能與精子受精過(guò)程有關(guān)[8]。KEGG通路富集分析,bta-miR-34b/c和bta-miR-449a/b/c的靶基因顯著富集在HIF-1信號(hào)傳導(dǎo)途徑、甲狀腺激素信號(hào)通路和Wnt信號(hào)通路等。缺氧誘導(dǎo)因子(hypoxia-inducible factor,HIF)是細(xì)胞對(duì)缺氧反應(yīng)的關(guān)鍵轉(zhuǎn)錄調(diào)節(jié)因子,HIF由α亞單位(HIF-1α、HIF-2α或HIF-3α)和β亞單位(HIF-β)組成,HIF-1α和HIF-2α密切相關(guān),它們與HIF-β亞基的相互作用分別產(chǎn)生HIF-1和HIF-2,HIF-1和HIF-2 2種活性轉(zhuǎn)錄因子均與缺氧反應(yīng)元件(hypoxia-response element,HRE)結(jié)合,刺激與血管生成、能量代謝、細(xì)胞生長(zhǎng)和細(xì)胞周期進(jìn)展等相關(guān)的靶基因調(diào)節(jié)生物學(xué)過(guò)程[44]。HIF-1α和HIF-2α在睪丸支持細(xì)胞中的表達(dá)已經(jīng)得到證實(shí)[45],且HIF-2參與促卵泡素(follicle-stimulating hormone,FSH)對(duì)支持細(xì)胞增殖的調(diào)節(jié)[46]。Gruber等[47]對(duì)出生2 d的小鼠進(jìn)行HIF-2α消融手術(shù),導(dǎo)致小鼠支持細(xì)胞無(wú)法建立完整的血睪屏障,并伴有睪丸大小和重量減少,且經(jīng)消融手術(shù)處理的小鼠最終不育。本研究中靶基因富集到的另一個(gè)重要通路是Wnt信號(hào)通路,Wnt信號(hào)是一種高度保守的細(xì)胞間通信機(jī)制。典型的Wnt信號(hào)傳導(dǎo)也稱(chēng)為Wnt/β-連環(huán)蛋白通路,通常被認(rèn)為是一種干細(xì)胞自我更新機(jī)制[48]。原始生殖細(xì)胞的特異性分化和雄性胎兒生殖道的正常發(fā)育依賴(lài)于Wnt/β-catenin信號(hào)傳導(dǎo)[49]。Takase等[50]研究報(bào)道,睪丸中Wnt信號(hào)通路特別有助于精原干細(xì)胞(spermatogonial stem cells,SSC)的增殖,證實(shí)了本研究結(jié)果的可靠性。
4 結(jié) 論
bta-miR-34b/c、bta-miR-449a/b/c通過(guò)上游轉(zhuǎn)錄因子和下游靶基因組成的分子調(diào)控網(wǎng)絡(luò)共同調(diào)節(jié)精子發(fā)生過(guò)程。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫(huà)刊(2023年8期)2023-08-01 15:45:12
科學(xué)大眾(2021年6期)2021-07-20 07:42:44
科學(xué)(2020年3期)2020-11-26 08:18:30
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂(lè)園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
湖北師范大學(xué)學(xué)報(bào)(自然科學(xué)版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
