4種哺乳動物乳中低聚糖的定性和定量分析研究進展
2022-07-29 02:40:58藍航蓮張麗娜
食品科學 2022年13期
關鍵詞:質量
藍航蓮,施 悅,張麗娜,*,周 鵬
(1.內蒙古乳業技術研究院,內蒙古 呼和浩特 010110;2.江南大學食品學院,江蘇 無錫 214122)
母乳作為嬰兒的“黃金食品”,不僅為嬰兒的生長發育提供完整的營養物質,且含有多種生物活性成分,例如活性蛋白、活性脂類、低聚糖及生長因子等,在嬰兒腸道微生物菌群的定植及免疫系統的建立及完善中發揮著重要的作用。
低聚糖是乳中的一類生物活性成分,具有多種功能活性。首先,低聚糖可作為益生元刺激益生菌生長。其在胃中難以被消化,通過腸道下部內源性微生物群發酵產生短鏈脂肪酸。短鏈脂肪酸可為腸黏膜細胞提供能量;此外,還能夠增加腸道滲透壓,加速腸道蠕動,軟化腸道內容物,促進排便;同時還可降低腸道內pH值以提供酸性環境,抑制有害菌的定植和生長,維持腸道內菌群正常化。其次,低聚糖具有顯著的抗病毒活性,其結構與腸黏膜細胞上的糖脂/糖蛋白相似,可以干擾病原體與腸黏膜的結合,以保護嬰兒免受腸道有害菌及其他病原體的侵害。再次,低聚糖在抗炎、免疫調節、促進大腦發育等方面也具有一定的作用。
低聚糖不僅存在于人乳中,山羊、牛、豬、駱駝等哺乳動物的初乳和成熟乳中也存在多種低聚糖。低聚糖一般由3~10個單糖通過糖苷鍵連接而成,組成低聚糖的單糖有-葡萄糖(glucose,Glc)、-半乳糖(galactose,Gal)、-乙酰氨基葡萄糖(-acetylglucosamine,GlcNAc)、-巖藻糖(fucose,Fuc)、-乙酰神經氨酸(-acetylneuraminic acid,Neu5Ac)和-羥乙酰神經氨酸(-glycolylneuraminic acid,Neu5Gc)。乳低聚糖的還原端通常由一個核心結構組成,在人乳低聚糖中核心結構一般為Gal-(1→4)-Glc,在動物乳中發現了Gal-(1→4)-GlcNAc這一不同的核心結構。特定的糖基轉移酶將上述單糖殘基從核苷酸糖轉移到低聚糖核心結構的非還原端形成低聚糖,由于人乳中單磷酸胞苷-Neu5Ac羥化酶的缺失,攜帶Neu5Gc的低聚糖只在動物乳中被發現。
目前,已有相關研究表明動物乳中的低聚糖具有與人乳低聚糖相似的生物活性功能。將動物乳中的低聚糖大規模提取,作為一種新型食品組分添加至嬰幼兒配方乳粉或功能性食品中,具有一定的應用前景。哺乳動物乳作為極具吸引力的生物活性低聚糖來源,確定其中低聚糖的種類及含量具有重要意義,可為合理選擇優質乳源提供依據。目前國內外就哺乳動物乳中低聚糖的種類及含量已經展開了大量的研究,本文對文獻中報道過的4種哺乳動物(山羊、牛、豬、駱駝)的初乳和成熟乳中的低聚糖種類及含量進行對比和分析,旨在為后續動物乳源低聚糖的開發與利用提供參考與指導。
1 哺乳動物乳中低聚糖的結構
低聚糖的結構具有復雜性和多樣性,其結構包括單糖的組成、糖殘基的連接順序、連接位置。單糖組成相同的低聚糖可能存在不同的結構,例如6’-唾液基-乳糖(6’-sialyllactose,6’-SL)和3’-唾液基-乳糖(3’-sialyllactose,3’-SL)的單糖組成均為Neu5Ac-Gal-Glc,但由于二者的唾液酸殘基的連接位置的差異,6’-SL對應的結構為Neu5Ac-(2→6)-Gal-(1→4)-Glc,3’-SL對應的結構則為Neu5Ac-(2→3)-Gal-(1→4)-Glc。由于在不同的研究中采用的儀器及檢測方法不同,鑒定到的低聚糖結構完整程度也不同。本文就單糖組成的水平,對文獻中已經報道過的山羊、牛、豬和駱駝這4種哺乳動物乳中的低聚糖種類進行了整理。在本文中,類似6’-SL與3’-SL的同分異構體統一被描述為單糖組成的Hex2Neu5Ac1(或Neu5Ac-Gal-Glc)酸性低聚糖。如表1所示,在這4種哺乳動物的初乳/成熟乳中檢測到84種不同單糖組成的低聚糖。其中山羊乳含有43種不同單糖組成的低聚糖,牛乳中有64種不同單糖組成的低聚糖,豬乳中有46種不同單糖組成的低聚糖,駱駝乳中有26種不同單糖組成的低聚糖。

表1 山羊、牛、豬、駱駝乳中報道的低聚糖種類[18,22-40]Table 1 Types of oligosaccharides reported in goat, cow, pig, and camel milk[18,22-40]
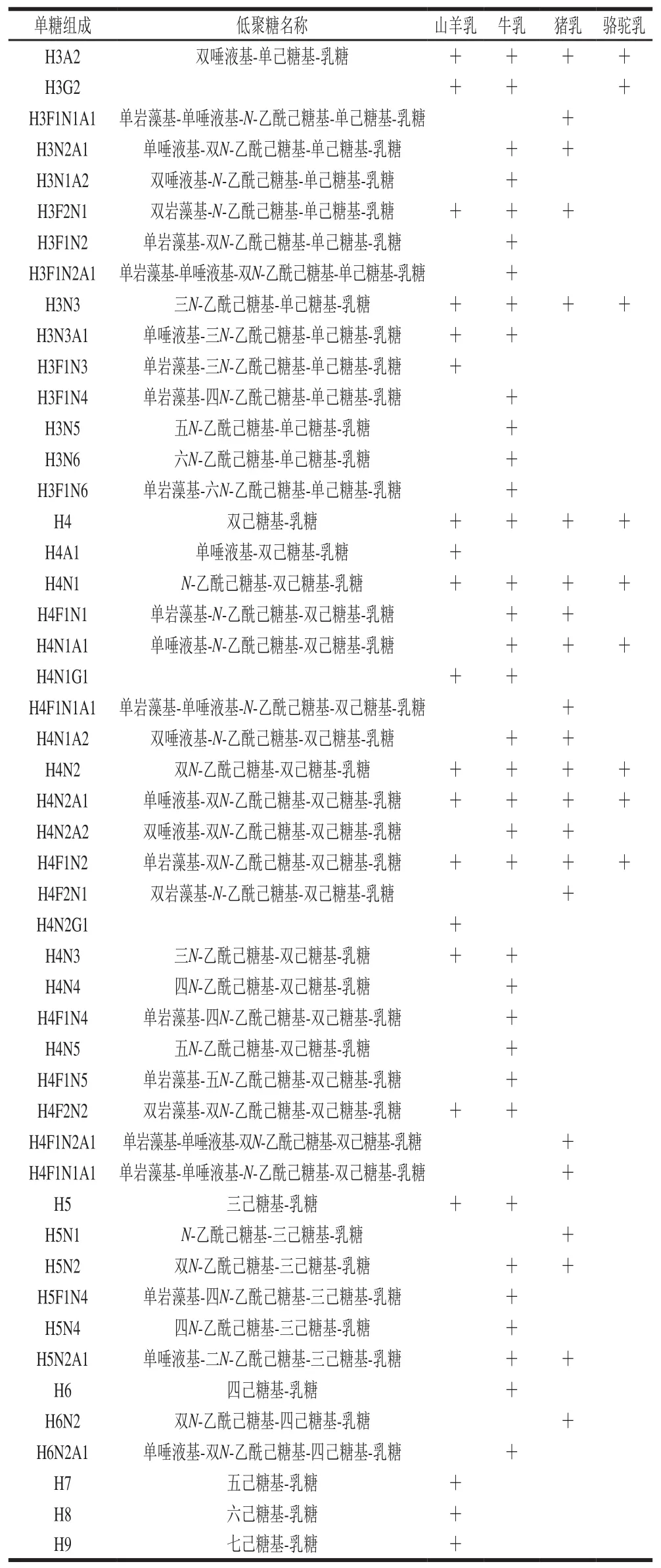
續表1
哺乳動物乳中檢測到的低聚糖類型可以分為中性低聚糖和唾液酸化的低聚糖兩大類。中性低聚糖包括非巖藻糖基化的中性糖和巖藻糖基化的中性糖,唾液酸化低聚糖包括帶有Neu5Ac或帶有Neu5Gc的低聚糖,或是既帶有Neu5Ac又帶有Neu5Gc的低聚糖,還有特殊的既帶有巖藻糖基又帶有Neu5Ac的低聚糖。
Martinez-Ferez等對山羊成熟乳中的低聚糖進行分析,共發現了7種單糖組成不同的非巖藻糖基化中性低聚糖和12種酸性低聚糖。Meyran等在Alpine山羊乳中鑒定到了31種單糖組成不同的低聚糖,其中16種為中性低聚糖(含5種巖藻糖基化中性糖),15種為酸性低聚糖。Martin-Ortiz等在Murciano-Granadina山羊初乳中鑒定出23種單糖組成不同的低聚糖,其中9種為中性非藻糖基化低聚糖,3種為巖藻糖基化中性低聚糖,11種為唾液化低聚糖;2017年,Martin-Ortiz等對Murciano-Granadina山羊成熟乳中的低聚糖展開了分析。結果表明,成熟乳中的低聚糖種類少于初乳中的低聚糖種類,在成熟乳中只鑒定到18種單糖組成不同的低聚糖,其中6種為中性非巖藻糖基化低聚糖,1種為巖藻糖基化中性低聚糖,11種為唾液酸化低聚糖。對山羊乳初乳和成熟乳中檢測到的低聚糖單糖組成進行對比,發現單糖組成為H1F1N1、H2F1、H2A2、H3A3、H3N3、H4N1和H5的低聚糖只在初乳中存在,而并未在成熟乳中發現;單糖組成為H3N3A1、H4N2A1這兩種低聚糖只在成熟乳中被發現。Albrecht等結合液相色譜串聯質譜技術,通過連接酶和特異性外切糖苷酶進行測序,對山羊乳中的低聚糖結構進行表征。在山羊成熟乳中共檢測到了25種單糖組成不同的低聚糖,其中10種為中性糖(含2種巖藻糖基化中性糖)、13種為酸性糖。Lu Jing等對中國關中和薩能山羊乳中的低聚糖種類進行了測定及比較,在這兩種山羊乳中共檢測到33種單糖組成不同的低聚糖,其中13種為非巖藻糖基化中性低聚糖,2種為巖藻糖基化聚糖,18種為酸性低聚糖。
Tao等在Jersey和Holstein牛乳中鑒定到了7種單糖組成不同的非巖藻糖基化中性聚糖、18種酸性聚糖。2010年,Barile等在Holstein牛的初乳和成熟乳中的低聚糖中發現了17種不同單糖組成的低聚糖,其中包括5種非巖藻糖基化中性低聚糖、2種巖藻糖基化中性糖和10種酸性低聚糖。2011年,Marino等使用親水作用液相色譜和外糖苷酶消化相結合的方法,對Holstein-Fresian奶牛初乳中的21種不同單糖組成的游離聚糖(2種非巖藻糖基化中性糖、6種巖藻糖基化中性糖和13種唾液化低聚糖)進行了結構鑒定。2012年,Sundekilde等對Jersey和Holstein牛乳中的低聚糖種類進行對比,在兩種牛乳中共檢測到了33種單糖組成不同的低聚糖,其中包括16種非巖藻糖基化中性糖、5種巖藻糖基化中性糖和12種酸性糖。2013年,Aldredge等對Jersey和Holstein牛初乳中的低聚糖進行了全面的結構分析,在牛初乳中確定了22種單糖組成不同的低聚糖,其中15種為中性糖(含4種巖藻糖基化中性糖)、7種為酸性糖。2014年,Albrecht等在Holstein-Friesian牛初乳中檢測到了9種單糖組成不同的中性低聚糖(包含3種巖藻糖基化中性聚糖)、15種酸性低聚糖。2020年,Chen Wenyan等使用泛甲基化修飾結合基質輔助激光解吸飛行時間質譜對市售牛乳中的游離低聚糖進行分析,在牛乳中發現8種巖藻糖基化低聚糖。
Tao等在豬乳中共檢測到了11種單糖組成不同的低聚糖,包括6種中性低聚糖(其中包含2種低豐度的巖藻糖基化中性低聚糖)和5種酸性低聚糖。Albrecht等對Landrace豬的初乳進行檢測,共發現了9種單糖組成不同的中性低聚糖(包括2種巖藻糖基化中性聚糖)、6種酸性低聚糖。2016年,Difilippo等對長白豬的初乳和成熟乳中的低聚糖進行分析,共鑒定到13種單糖組成不同的低聚糖,其中包括6種非巖藻糖基化中性低聚糖、2種巖藻糖基化中性低聚糖和5種酸性低聚糖。Cheng Likun等利用親水作用液相色譜串聯四極桿-飛行時間質譜對Landrace-Large White、Duroc和Yorshire 3個品種豬乳中的低聚糖進行檢測及分析。在這3種豬乳中檢測到的低聚糖共有27種不同的單糖組成,其中包括6種非巖藻糖基化中性低聚糖、5種巖藻糖基化中性低聚糖和16種酸性低聚糖。2017年,張佳妮利用基質輔助激光解吸飛行時間質譜技術對長白豬、大白豬、梅山豬3個品種豬乳中的低聚糖進行結構解析,共檢測到14種單糖組成不同的低聚糖,其中包括5種非巖藻糖基化中性糖、2種巖藻糖基化中性糖和7種酸性糖。
Fukuda等采用H-核磁共振波譜對雙峰駝的初乳和成熟乳中的低聚糖進行了鑒定和表征。結果發現,在初乳中鑒定到了9種單糖組成不同的低聚糖,其中包括3種中性低聚糖(含1種巖藻糖基化中性聚糖)和6種酸性低聚糖,在成熟乳中只鑒定到3種低聚糖,分別為Gal-(1→3)-Gal-(1→4)-Glc、Gal-(1→4)-GlcNAc-(1→6)-(Gal1-3)Gal-(1→4)-Glc和Neu5Ac-(2→)-Gal-(1→4)-Glc),而Gal-(1→4)-GlcNAc-(1→6)-(Gal1→3)-Gal-(1→4)-Glc只在成熟乳中發現而未在初乳中檢測到。2013年,Alhaj等也采用H-核磁共振波譜的方法對市售單峰駝乳中的低聚糖進行了鑒定,共鑒定到了6種單糖組成不同的低聚糖,其中包括2種中性低聚糖和4種酸性低聚糖。2014年,Albrecht等使用超高效液相色譜串聯二級質譜技術對單峰駝乳中低聚糖進行表征。在單峰駝成熟乳中共檢測到了25種單糖組成不同的低聚糖,其中包括 5種巖藻糖基化中性糖、9種非巖藻糖基化中性低聚糖和11種酸性低聚糖。
如圖1A所示,在單糖組成的水平上,在山羊乳中發現了22種中性低聚糖和21種酸性低聚糖,在牛乳中發現了32種中性低聚糖和32種酸性低聚糖,在豬乳中發現了21種中性低聚糖和25種酸性低聚糖,在駱駝乳中發現了14種中性低聚糖和12種酸性低聚糖。
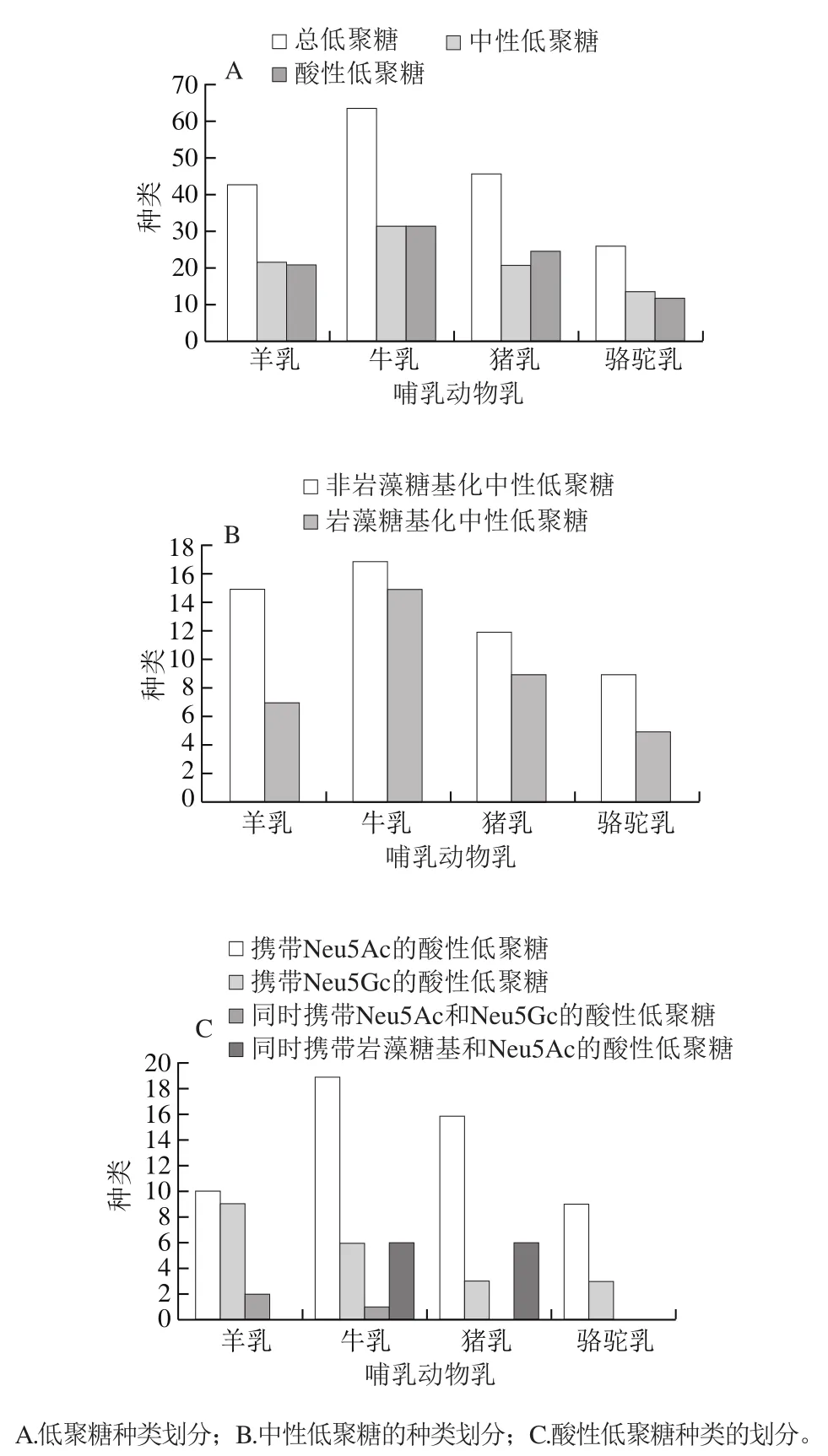
圖1 不同哺乳動物乳中低聚糖的種類[18,22-40]Fig. 1 Types of oligosaccharides in milks from different mammals[18,22-40]
革蘭氏陰性菌多通過細菌內毒素與腸道Toll樣受體4(Toll-like receptor 4,TLR4)結合以激活黏膜炎癥。巖藻糖基化低聚糖被發現可以調節人腸細胞中白細胞分化抗原(cluster of differentiation,CD)14的表達,抑制TLR4信號,從而減輕由細菌內毒素誘導的新生兒壞死性小腸結腸炎等炎癥。從動物乳中提取純化的巖藻糖基化低聚糖,可以作為一種新型的口服預防藥物或治療藥物,以減輕各種黏膜炎癥。如圖1B所示,巖藻糖基化低聚糖在牛乳和豬乳中發現的種類較多,牛乳中發現了15種,單糖組成分別為H1F1N1、H2F1、H2F1N1、H3F1、H3F1N1、H3F2N1、H3F1N2、H3F1N4、H3F1N6、H4F1N1、H4F1N2、H4F1N4、H4F1N5、H4F2N2、H5F1N4;在豬乳中發現了10種,單糖組成分別為H1F1N1、H2F1、H2F1N1、H2F2、H3F1N1、H3F2N1、H4F1N1、H4F1N2、H4F2N1、H4F2N2;在山羊乳和駱駝乳中分別發現了7種和5種巖藻糖基化低聚糖,單糖組成分別為H1F1N1、H2F1、H3F1N1、H3F2N1、H3F1N3、H4F1N2、H4F2N2(山羊乳);H2F1、H2F1N1、H3F1、H3F1N1、H4F1N2(駱駝乳)。其中單糖組成為H4F1N4、H3F1N6、H5F1N4、H3F1N4、H4F1N5、H3F1N2的巖藻糖基化低聚糖目前只在牛乳中發現,單糖組成為H3F1N3的巖藻糖基化低聚糖只在山羊乳中發現,在豬乳發現其特有的H2F2和H4F2N1這兩種單糖組成的巖藻糖基化低聚糖。
炎癥過程中,白細胞通過黏附和穿越血管內皮細胞釋放活性氧自由基和酶,造成血管損傷及嚴重的組織損傷。唾液酸化的低聚糖被發現能夠降低白細胞與內皮細胞的黏附從而減輕炎癥反應,其可能作為一種抗炎成分有利于降低母乳喂養嬰兒炎癥性疾病的發病率。如圖1A所示,在單糖組成的水平上,在牛乳和豬乳中發現了種類較多的酸性低聚糖。有相關研究表明,感染小牛和仔豬的輪狀病毒對唾液酸酶敏感,其與細胞的附著和結合被認為依賴于唾液酸,牛乳和豬乳中被發現具有種類較多并且濃度較高的唾液酸化低聚糖,可通過水解釋放末端的唾液酸,與上皮細胞表面競爭輪狀病毒的結合位點,并與病毒結合阻斷其傳染,為小牛和仔豬提供保護。
與牛乳、豬乳和駱駝乳不同的是,山羊乳中發現的攜帶Neu5Gc的酸性低聚糖的種類數與攜Neu5Ac的酸性低聚糖的種類數相近。此外,在山羊乳和牛乳中發現了同時攜帶Neu5Ac和Neu5Gc的低聚糖,H2A1G1這一單糖組成的低聚糖同時也在山羊乳和牛乳中被發現,山羊乳中還發現了其特有H3A1G1這一單糖組成的酸性低聚糖。還有一類低聚糖,同時攜帶巖藻糖基和Neu5Ac基團,它們在牛乳和豬乳中均有被發現,在牛乳中發現了6種,單糖組成分別為H1F1N1A1、H2F1A1、H2F1N1A1、H2F1N1A2、H2F1N2A1、H3F1N2A1;在豬乳中也發現了6種,單糖組成分別為H1F1N1A1、H2F1N2A1、H3F1N1A1、H4F1N1A1、H4F1N2A1、H4F1N1A1。其中H1F1N1A1和H2F1N2A1這兩種單糖組成的低聚糖為牛乳和豬乳中共有。在豬乳和牛乳中都檢測到了多種巖藻糖基低聚糖以及同時攜帶巖藻糖基和Neu5Ac基團的聚糖,這說明豬乳和牛乳低聚糖在結構上可能具有一定的相似性。目前,對攜帶Neu5Gc的酸性低聚糖、同時攜帶Neu5Ac和Neu5Gc的酸性低聚糖以及一些同時攜帶巖藻糖基和唾液酸基團的低聚糖的生理功能的研究較少,有待進一步的研究與討論。
將在山羊、牛、豬和駱駝這4種哺乳動物乳中發現的不同單糖組成的低聚糖種類進行對比,如圖2所示,哺乳動物乳中的低聚糖存在一定的相似性,以牛乳和豬乳最為相似,二者共有37種單糖組成相同的低聚糖,這個數據與上文的分析結果一致。其次是牛乳和山羊乳,共有33種單糖組成相同的低聚糖。第三是牛乳和駱駝乳,共發現了26種單糖組成相同的低聚糖。在不同的哺乳動物乳中也發現了一些其特有的低聚糖。在山羊乳中發現11種其特有的低聚糖,單糖組成分別為H3F1N3、H9、H8、H5、H3A1G1、H4A1、H4N2G1、H2G2、H2N4、H7、H2N1G1;在牛乳中發現18種其特有的低聚糖,單糖組成分別為H3N5、H2F1A1、H3N6、H3N1A2、H6N2A1、H2F1N1A2、H4N4、H2F1N1A1、H4F1N5、H6、H3F1N6、H2N1A2、H3F1N2A1、H3F1N4、H3F1N2、H4F1N4、H5F1N4、H4N5;在豬乳中也發現了8種其特有的低聚糖,分別為H1N2A1、H2F2、H5N1、H4F2N1、H4F1N2A1、H6N2、H3F1N1A1、H4F1N1A1;而在駱駝乳中未發現其特有的低聚糖。

圖2 山羊、牛、豬、駱駝乳中報道的低聚糖種類的Venn圖[18,22-40]Fig. 2 Venn diagram of oligosaccharide species reported in goat, cow,pig, and camel milk[18,22-40]
通過對4種哺乳動物乳中檢測到的低聚糖種類進行分析可知,牛乳和豬乳中存在種類較多的巖藻糖基化低聚糖和唾液酸化低聚糖,可作為提取動物乳源巖藻糖基化低聚糖和唾液酸化低聚糖的較適乳源。而山羊乳中含有種類較多的攜帶Neu5Gc的酸性低聚糖,可對此進行山羊乳源中特有低聚糖的功能活性研究。
2 哺乳動物乳中低聚糖的含量
由于目前在駝乳方面的研究還較少,只有Zhang Heping等的研究表明Bactrian駱駝初乳中乳糖含量約占總固形物的4.44%,成熟乳中乳糖含量約占總固形物的4.24%,但對低聚糖的含量還未進行深入的研究。因此,本文就目前文獻中報道的山羊乳、牛乳和豬乳中的低聚糖質量濃度,以及6’-SL、3’-SL和GL這3種目前研究較多且質量濃度表現為較高的低聚糖質量濃度進行總結和對比,結果分別如表2~4所示。

表2 山羊乳中報道的低聚糖質量濃度Table 2 Contents of oligosaccharides reported in goat milk
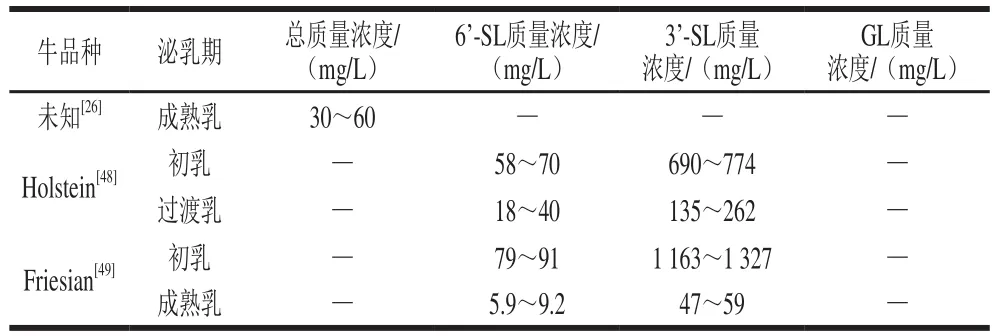
表3 牛乳中報道的低聚糖質量濃度Table 3 Contents of oligosaccharides reported in cow milk
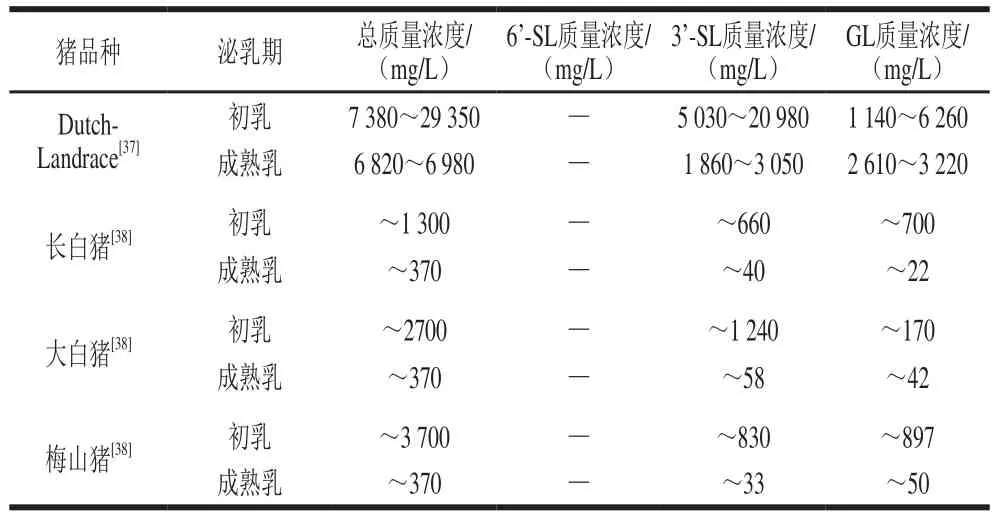
表4 豬乳中報道的低聚糖質量濃度Table 4 Contents of oligosaccharides reported in porcine milk
如表2所示,Martinez-Ferez等對山羊成熟乳中低聚糖質量濃度進行檢測,總質量濃度約為250~300 mg/L。其中酸性低聚糖以6’-SL質量濃度最高(50~70 mg/L),中性低聚糖以3’-半乳糖基-乳糖(3’-galactosyl-lactose,3’-GL)質量濃度最高(30~50 mg/L)。Claps等對兩種意大利山羊(Garganica羊和Maltese羊)的初乳及成熟乳中3種酸性低聚糖(3’-SL、6’-SL和雙唾液基-乳糖(disialyllactose,DSL))的質量濃度進行檢測。結果表明,在初乳中,Garganica羊的3’-SL和6’-SL質量濃度均高于Maltese羊,DSL的平均質量濃度略低于Maltese羊。其中Garganica羊初乳中3’-SL質量濃度為253.96~328.54 mg/L,6’-SL質量濃度為174.27~200.96 mg/L,DSL質量濃度為126.44~137.94 mg/L;Maltese羊初乳中3’-SL質量濃度為201.27~249.33 mg/L,6’-SL質量濃度為136.90~144.07 mg/L,DSL為104.91~228.14 mg/L,這一結果與Martinez-Ferez等的研究結果不同。Meyrand等對Alpine山羊成熟乳中的低聚糖進行定量,低聚糖總質量濃度約為1 110~1 350 mg/L,比文獻[26]中所報道的山羊乳中的低聚糖質量濃度高。Thum等對New Zealand Saanen山羊初乳和成熟乳中的低聚糖進行定量,結果表明初乳中低聚糖總質量濃度為236~404 mg/L,成熟乳中低聚糖總質量濃度為201~319 mg/L。
Martin-Ortiz等對Murciano Granadina山羊初乳中的低聚糖質量濃度進行檢測,總質量濃度為251.22~572.24 mg/L,其中酸性低聚糖的質量濃度為83~251 mg/L,中性低聚糖的質量濃度為140~350 mg/L。在酸性低聚糖中,以6’-SL質量濃度最高(28.85~123.76 mg/L),3’-SL次之(3.05~11.99 mg/L);在中性低聚糖中,以GL的異構體質量濃度最高,總質量濃度為124.92~265.77 mg/L。在巖藻糖基化低聚糖中,2’-巖藻基-乳糖(2’-fucosyl-lactose,2’-FL)與Fuc-GalNAc-Glc均顯示較高質量濃度,分別為2.21~31.59 mg/L和3.08~6.15 mg/L。之后,他們對不同泌乳期(泌乳期為2、10、30、40 d和泌乳期為1、7、30、120 d)的Murciano Granadina山羊乳中的低聚糖質量濃度進行研究。結果表明,泌乳期從2 d延長到40 d所收集到的羊乳樣品,低聚糖總質量濃度由199.1 mg/L減少到59.2 mg/L;泌乳期從1 d延長到120 d所收集到的羊乳樣品,低聚糖總質量濃度由488 mg/L減少到112.4 mg/L。在不同泌乳期間,6’-SL的質量濃度均高于3’-SL,泌乳期第一天的羊乳樣品中6’-SL質量濃度最高,為51.4~250.0 mg/L。Marziali等對泌乳期為1~4 d的Murciano Granadina山羊乳中的低聚糖質量濃度進行測定,發現低聚糖的總質量濃度的平均值由第1天的2 398.3 mg/L減少到第4天的702.7 mg/L,泌乳期第1天,在絕對定量的6種低聚糖中,中性Gal-Gal-Glc質量濃度的平均值為390.3 mg/L;酸性3’-SL的質量濃度(平均值為1 421.2 mg/L)高于6’-SL(平均值為221.3 mg/L),這與Martin-Ortiz等的結果相反。并且,同樣是對Murciano Granadina這個品種山羊的乳低聚糖質量濃度進行研究,Marziali等測定的低聚糖質量濃度高于Martin-Ortiz等的結果,即同一品種山羊乳中的低聚糖的質量濃度存在差異,可能是受畜牧地區、產崽及泌乳的季節氣候和產崽胎次等因素影響,后續的研究可以針對不同地區及不同季節的Murciano Granadina山羊乳中的低聚糖質量濃度進行對比分析。
如表3所示,Martinez-Ferez等檢測到牛的成熟乳中低聚糖質量濃度為30~60 mg/L。Nakamura等對Holstein牛初乳及過渡乳中3種酸性低聚糖(3’-SL、6’-SL和6’-唾液基--乙酰乳糖胺(6’-sialyl--acetyllactosamine,6’-SLN))的質量濃度進行檢測。結果顯示,牛乳中3’-SL質量濃度高于6’-SL和6’-SLN。在牛初乳中,3’-SL質量濃度為690~744 mg/L,在過渡乳中迅速下降至135~262 mg/L。6’-SL和6’-SLN的質量濃度在牛初乳中也最高,分別為58~70 mg/L和93~103 mg/L,而在過渡乳中其質量濃度分別降為18~40 mg/L和64~74 mg/L。Fong等采用外標法對Friesian牛初乳和成熟乳中的酸性低聚糖進行絕對定量。結果表明3’-SL的質量濃度高于其他酸性低聚糖。在產后收集的初乳中3’-SL的質量濃度為1 163~1 327 mg/L,質量濃度隨著哺乳期的延長而減少,在成熟乳中檢測到3’-SL的質量濃度為47~59 mg/L。在產后收集的初乳中,6’-SLN、DSL和6’-SL的質量濃度也較高,分別為112~126、118~134 mg/L和79~91 mg/L;在成熟乳中質量濃度均有所下降,分別為0.11~0.15、1.4~1.6 mg/L和5.9~9.2 mg/L。
如表4所示,Difilippo等對Dutch-Landrace豬初乳和成熟乳中的低聚糖質量濃度進行檢測。初乳中低聚糖總質量濃度為7 380~29 350 mg/L,其中酸性低聚糖質量濃度最高,約占總聚糖總量的77%;酸性低聚糖中又以3’-SL的質量濃度最高,約為5 030~20 980 mg/L。成熟乳中低聚糖總質量濃度較初乳下降,為6 820~6 980 mg/L,其中中性低聚糖的相對含量由初乳的23%提高到了58%,中性低聚糖中以Hex3的質量濃度較高,約為2 610~3 220 mg/L。Cheng Likun等收集了泌乳期為1、5、10、21 d的豬乳,并對乳中的低聚糖進行相對定量分析,結果表明,不同的低聚糖在泌乳過程中表現出不同的規律,大部分的低聚糖在初乳中質量濃度較高,并隨著泌乳期時間的延長質量濃度降低,但也有少數低聚糖的質量濃度增加。
張佳妮以長白豬、大白豬、梅山豬為研究對象,使用麥芽五糖對豬乳中的低聚糖進行外標定量,將各個泌乳期豬乳中低聚糖的總質量濃度進行比較。結果顯示,3個品種豬乳中初乳的低聚糖總質量濃度存在差異,其中梅山豬初乳中的低聚糖質量濃度最高,約為3 700 mg/L,大白豬次之(約2 700 mg/L),長白豬豬乳中的低聚糖質量濃度最低,約為1 300 mg/L。隨著哺乳期的延長,這3個品種的豬乳中的低聚糖質量濃度差異逐漸減小。且初乳中的低聚糖總質量濃度高于成熟乳,大約是其3.5~12.5 倍。
通過對比表2的數據可知,豬乳中的低聚糖質量濃度遠高于山羊乳和牛乳,且在張佳妮的研究中發現,梅山豬乳中的低聚糖質量濃度高于大白豬和長白豬,梅花豬的初乳可能成為提取純化動物乳源低聚糖的首選。Murciano Granadina山羊乳和Alpine山羊乳中也含有較高質量濃度的低聚糖,在以山羊為主要飼養家畜的地區,可選擇山羊乳作為工業化提取低聚糖的乳源。同時,乳低聚糖的質量濃度可能隨地區、氣候、季節及產崽胎次的變化而變化,可針對某一較適宜物種,進行不同地區、不同產崽及泌乳季節和不同產崽胎次個體乳的低聚糖質量濃度比較,以選取最佳乳源。而目前的研究結果表明,單從低聚糖的質量濃度方面考慮,牛乳中的低聚糖質量濃度較山羊乳和豬乳低,可能不是提取低聚糖的較佳乳源。
3 結 語
乳低聚糖在醫療保健和食品工業中的應用前景十分廣闊。例如巖藻糖基化低聚糖可以抑制細菌和受體的結合,有潛力作為一種不同于目前抗生素作用機制的新型預防或治療抗菌藥物。相關研究表明,使用在嬰幼兒配方奶粉中添加2’-FL的復合乳粉喂養的嬰兒的血液和體外炎癥細胞因子水平與母乳喂養組顯示的水平接近,且均低于添加半乳糖的嬰幼兒配方奶粉的復合乳粉喂養組。2’-FL的添加有望使配方奶粉更接近母乳,從而進一步實現配方奶粉的母乳化。唾液酸低聚糖被發現具有抗炎的作用,有助于降低母乳喂養嬰兒的炎癥性疾病的發病率。動物乳源唾液酸化低聚糖也被證明了具有抵抗炎癥疾病的生理功能,有相關研究表明,與對照組(不口服山羊乳低聚糖)相比,口服500 mg/(kg·d)山羊乳低聚糖的大鼠結腸病變較少,腸道菌群組成更為正常,說明山羊乳低聚糖具有抗炎作用,有助于恢復結腸黏膜。此外,動物乳可能是膳食補充唾液酸的極好的來源。從動物乳中分離出來的唾液酸化低聚糖可作為腦活化因子應用于嬰兒配方奶粉和其他功能食品中。
動物乳初乳中低聚糖的種類較成熟乳豐富,低聚糖的含量也較成熟乳高,可作為提取純化低聚糖的首選來源。近年來研究者對動物乳中的低聚糖開展了大量研究,但大多數研究僅以表征低聚糖的結構和含量為目標,而對其生理功能研究較少。雖然已有研究表明從動物乳源提取的低聚糖和人乳低聚糖具有相似的功能特性,但是否具有相同的生物利用度仍然是一個有待證實的問題。其次,動物乳中低聚糖的含量較低,需要高通量、高純度分離純化低聚糖的方法來滿足工業化生產的要求。
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
