小麥貴協3號成株期抗條銹病基因的遺傳定位
2022-08-08 09:25:56葛昌斌丁延慶辛智海張立異
麥類作物學報 2022年8期
高 旭,程 斌,高 煜,葛昌斌,丁延慶,曹 寧,辛智海,張立異
(1.貴州省農業科學院旱糧研究所,貴州貴陽 550006; 2.貴州大學農學院,貴州貴陽 550025;3.漯河市農業科學院,河南漯河 462300; 4.貴州大學生命科學學院,貴州貴陽 550025)
迄今為止,已經有34個小麥條銹菌生理小種(CYR1~CYR34)和40多個致病類群(如洛10、雜46、貴農22等)被命名,其中CYR32、CYR33和CYR34為當前中國小麥條銹菌的主要流行小種。截止2020年,全球已有83個抗條銹病基因被正式命名,有40多個抗條銹病基因被暫時命名。但是,貴農22致病類群出現以后,只有少數苗期抗病基因(、、等)對現有的條銹菌仍能保持有效的抗病性。CYR34小種主要分布在中國西南地區和西北甘肅等省份,目前該小種已經擴展至全國大部分麥區,對中國小麥安全生產和抗病育種工作造成嚴重威脅。因此,必須通過挖掘新的抗條銹病基因來拓寬抗性基因遺傳資源。
SNP芯片技術被廣泛應用于小麥研究中,是一種高通量、微型、自動化檢測SNP的手段。混池轉錄組測序(BSR-Seq)技術無需檢測全基因組,且不涉及重復序列,所以對基因組較大的物種而言,BSR-Seq更為簡便有效。Zhang等利用一個RIL群體(SW8588/Thatcher)并結合55K芯片,檢測到7個抗葉銹病QTL和6個抗條銹病QTL,其中有4個QTL對兩種病害兼具抗性。Jia等利用小麥品種濟麥23和泰農18雜交構建的F和F群體進行BSR-Seq,將抗白粉病基因定位在5DS染色體標記YTU3004和Swgi068/Bwm20之間。
貴協3號是利用以色列野生二粒小麥(,AABB,2=28)與光稃野燕麥(L.var.)進行雜交,獲得的F代與貴農22回交選育而成的普通小麥材料。2010―2021年,貴協3號在中國多個麥區對條銹病主要流行小種(CYR32、CYR33和CYR34)均表現為近免疫或高抗,充分說明該品種具有較好的持久抗病性,是一個具有利用價值的條銹病抗源。因此,本研究對貴協3號的條銹病抗性進行研究,準確定位其攜帶的抗條銹病基因,并發掘與抗條銹病基因緊密連鎖的分子標記,以期加快貴協3號在小麥抗病育種中的應用,保障中國小麥安全生產。
1 材料與方法
1.1 試驗材料
以來自澳大利亞的高感條銹病小麥品種Avocet S為母本,以來自貴州大學張慶勤教授惠贈的高抗條銹病品系貴協3號為父本,雜交得到的F種子自交形成F,F再以單籽粒傳法獲得227份F代重組自交系(RIL)群體,用于田間成株期條銹病的表型鑒定和基因型分析。感病對照品種為銘賢169。
1.2 成株期條銹病表型鑒定
于小麥成株期,在田間自然條件下對RIL群體及其親本進行條銹菌混合生理小種的條銹病抗性鑒定。于2018和2019年的10月中旬,將RIL群體及其親本種植在貴州省農業科學院貴陽基地(26°29′ N,106°39′ E);于2019年11月初,將RIL群體及其親本種植在四川省成都市雙流區試驗基地(30°57′ N,103°92′ E),三個環境分別用18GY、19GY和19CD來表示。每個家系種植1行,每行10株,每隔5行種植感病對照品種銘賢169作為誘發行。待銘賢169及感病親本Avocet S充分發病時,調查并記錄條銹病反應型,參照Cheng等的方法,采用0~4級分級標準,其中0~2級為抗病,3~4級為感病。
1.3 混池轉錄組測序及分析
采用TriZol法分別提取親本Avocet S和貴協3號的RNA,構建親本池。根據RIL群體條銹病抗性鑒定結果,從中選取20個抗病家系(反應型為0)建立抗病池(Br),選取20個感病家系(反應型為4)建立感病池(Bs)。每個家系分別取4粒種子播種于花盆中,置于培養箱(溫度16~25 ℃,光照16 h,光強22 000 lx)中進行培養,待幼苗長至2葉完全張開時,取每個單株的第二片葉中部(葉長4 cm),采用TriZol法分別提取RNA,等量混合,形成抗、感池。RNA樣品檢測合格后,用Illumina高通量測序平臺NovaSeq 6000進行測序,將得到的各樣品的原始數據(raw reads)進行評估,去除測序接頭、低質量堿基、空載體等不可信數據后,得到高質量的質控后數據(clean reads),用序列比對軟件STAR將其比對到中國春2.0參考基因組上。BSR-Seq數據分析由北京麥美瑞生物科技有限公司合作完成。利用STAR軟件將高質量轉錄組測序數據比對到中國春參考序列上并進行過濾,保留只有唯一比對位置且錯配數小于2%的序列比對結果,使用SAM tools軟件挖掘可能的變異位點,然后用Perl程序僅保留比對質量大于phred值15、變異質量大于phred值30、只有2種基因型、總深度在6~100 000之間、參考序列基因型深度大于3、變異基因型深度大于3、參考序列基因型深度比例大于5%以及變異基因型深度大于5%的比對結果。
1.4 55K SNP芯片的QTL定位
待RIL群體及其親本的幼苗長到一葉一心期時,用北京天根生化科技有限公司生產的植物基因組DNA提取試劑盒(DP320)提取DNA。用濃度為1%的瓊脂糖凝膠電泳和GelDoxXR凝膠成像系統對提取的DNA進行質量檢測,檢測合格后的DNA樣品利用55K SNP芯片進行基因分型。根據基因型數據的檢出率、最小等位基因頻率(minor allele frequency,MAF)、雜合率等指標對樣品和標記進行基本質控(quality control,QC)。樣品QC標準:DQC>0.82,檢出率≥85%、雜合率≤10%。標記QC標準:轉換類型(Conversion Type)為聚高分辨率(Poly High Resolution)、檢出率≥90%、MAF≥4%、雜合率≤10%、等位基因數=2。對經過QC的基因型數據進行標記位點篩選,只保留親本基因型純合且有差異的位點。利用JoinMap 4.0的Kosambi函數構建遺傳圖譜,將遺傳重組率相同的標記劃分到同一個Bin圖譜中。用QTL IciMapping 4.0軟件的完備區間作圖法(inclusive composite interval mapping,ICIM)進行QTL檢測,染色體步長是0.1 cM,逐步回歸概率為<0.001,尋找LOD值>2.5的QTL,計算出每個QTL的遺傳貢獻率和加性效應。
1.5 多態性SSR標記的篩選
根據BSR-Seq的分析結果和小麥55K SNP芯片的檢測結果,推斷染色體上差異SNP標記分布數目最多、密度最大的區域為貴協3號中抗條銹病基因所在的位置。之后根據Graingenes數據庫中目標區域已知的22個SSR標記(https://wheat.pw.usda.gov/)以及利用PrimerServer(http://202.194.139.32/PrimerServer/)在線設計的103個SSR標記進行標記多態性的篩選。SSR標記由南京金斯瑞生物科技有限公司合成。上述標記首先在親本和抗、感池間進行篩選,獲得的多態性標記在RIL群體中進行驗證,最終獲得6個與目標基因緊密連鎖的多態性SSR標記 (表1)。

表1 6個多態性SSR標記的引物信息Table 1 Primer information of six SSR markers
實驗采用10.0 μL PCR反應體系,包括模板DNA(80.0 ng·μL)1.0 μL,上、下游引物各(10 μmol·L)1.0 μL,2×Taq pcr mix 5.0 μL,ddHO 2.0 μL。PCR反應程序:94 ℃預變性5 min;94 ℃變性45 s,退火溫度視不同引物而定,退火30 s,72℃延伸1 min,35個循環;72 ℃延伸10 min,4 ℃保存。用8.0%非變性聚丙烯酰胺凝膠電泳對擴增產物進行電泳檢測,銀染顯色后觀察拍照。
1.6 數據分析
用Excel軟件對試驗數據進行統計分析,用SPSS軟件進行單因素方差分析和相關性分析。定位到的QTL側翼標記的物理位置和候選基因的相關信息通過小麥多組學網站(http://202.194.139.32)查詢;在Graingenes數據庫查找已經報道的QTL側翼標記或基因信息,并與國際小麥基因組測序聯盟(IWGSC,http://www.wheatgenome.org/)的中國春參考序列進行比對,獲得它們的物理位置;從小麥多組學網站獲得中國春的參考基因組注釋信息。用基于R語言的繪圖包ggplot2(https://ggplot2.tidyverse.org/)和Mapchart 2.32(https://www.wur.nl/en/show/Mapchart.htm)進行QTL峰值示意圖和染色體遺傳圖譜的繪制。
2 結果與分析
2.1 RIL群體及其親本成株期條銹病抗性的鑒定結果
對227個RIL群體及其親本在3個環境下進行成株期條銹病抗性鑒定。結果表明,3個環境下RIL群體條銹病表型數據均呈連續分布,群體中抗、感株系的材料個數比值接近1∶1。方差分析結果(表2)顯示,環境對條銹病的發生有顯著影響。3個環境間的相關系數在 0.337~0.569之間,18GY、19GY與19CD的相關系數均較低,原因可能是四川地區的條銹病小種以CYR34為主,田間流行頻率高于貴州地區,病害更嚴重(表3)。

表2 3個環境下條銹病表型數據的單因素方差分析Table 2 ANOVA for the phenotypic data of stripe rust in RIL population under three environments

表3 3個環境下RIL群體條銹病反應型的相關性分析Table 3 Correlation analysis of stripe rust infection type in RIL population under three environments
2.2 混池轉錄組測序(BSR-Seq)結果
2.2.1 轉錄組測序數據質量控制分析
對親本池和抗、感池分別進行轉錄組測序,結果(表4)表明,原始數據包含144 240 863個reads,質控后數據有139 976 742個reads,有效率為96.37%~98.11%,GC含量為57.00%~ 58.27%,質量值Q20在98.16%~98.36%之間,Q30在94.41%~94.88%之間。
2.2.2 差異SNP的數目和位置
利用RIL群體的BSR-Seq分析結果,在小麥全基因組中共鑒定到277個與條銹病抗性關聯的高置信度SNP或Indel。其中,2AS染色體末端上的SNP數目最多,為258個,占全部SNP總數的93.1%,有8個差異SNP位于2AL、3BL、4BS、5AL和6BS染色體上,有11個差異SNP未被定位在染色體上。因此,推測目標基因位于2AS染色體末端的12.0~59.0 Mb區間。
2.3 遺傳圖譜構建結果
利用小麥55K SNP芯片對227個RIL群體及其親本進行全基因組基因分型。對于雜合率大于10%的群體株系進行了過濾,去除了23個樣品DNA,剩余有效基因型樣本數為204個。在約53 000個SNP中,有16 155個SNP在兩個親本之間具有多態性,過濾后剩余15 559個SNP,其中14 486個SNP有物理位置。利用Joinmap 4.0構建遺傳連鎖圖譜時,剔除不連鎖的分子標記,最終用于作圖的標記數為6 364個。構建的遺傳連鎖圖譜覆蓋了小麥21條染色體(圖1和表5),圖譜總長度為4 363.9 cM,標記密度為1.5 cM。其中,3A染色體的長度最長(333.0 cM),6A染色體的長度最短(33.5 cM)。2A染色體的標記密度最大,為3.2 cM,其遺傳圖譜長度為258.3 cM。
從小麥A、B、D三個基因組來看,B基因組上的標記數最多,為2 744個,占總標記數的 43.1%,D基因組上的標記數最少,為1 083個,占總標記數的17.0%;從小麥七個同源群來看,第二同源群的標記數最多,為1 313個,占總標記數的20.6%,第六同源群的標記數最少,為513個,占總標記數的8.1%;從小麥21條染色體看,平均每條染色體上有303個標記,其中2A染色體上的標記數最多,為828個,占總標記數的 13.0%,6A染色體上的標記數最少,為48個,占總標記數的0.7%(表5)。
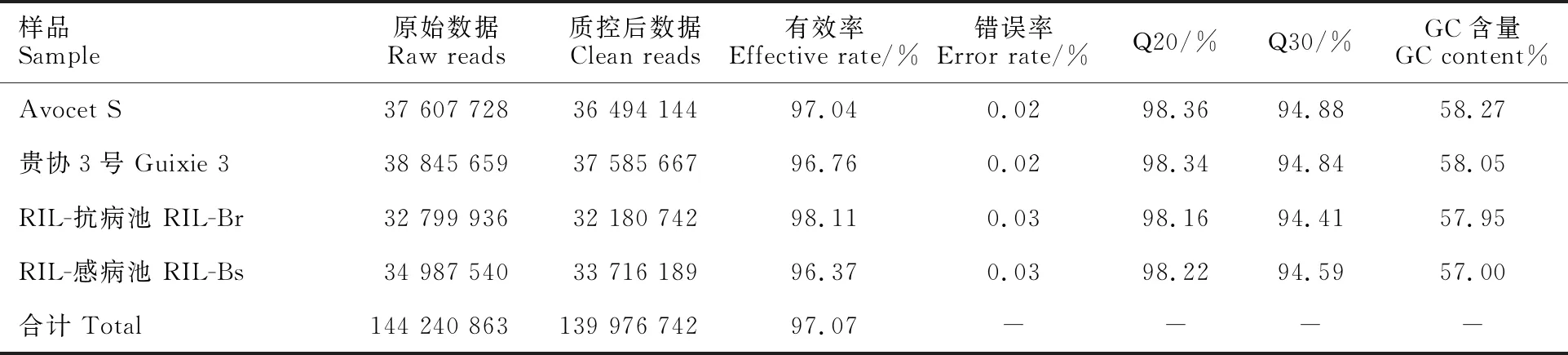
表4 轉錄組數據產出質量情況Table 4 Quality of RNA-Seq data

每條染色體內的黑、灰色代表標記所在的位置,顏色越深表明該位置的標記數越多;白色代表沒有連鎖標記的區間。
2.4 抗條銹病QTL的定位結果
根據小麥55K SNP芯片的基因型數據和條銹病表型鑒定數據,利用QTL IciMapping 4.0進行條銹病抗性QTL的定位。結果共檢測到7個抗條銹病QTL,分別分布在1B(1)、2A(2)、2D(1)、5A(2)和6B(1)染色體上,其中可在3個環境中被檢測到,其位于2AS染色體0.00~2.47 cM的遺傳區間內,物理區間為15.87~31.89 Mb(16.02 Mb),其3年的LOD值變化范圍為9.79~14.62,標記區間為AX-108824773~AX-111675237,可解釋的表型變異率為17.07%~34.59%,是一個穩定的主效QTL(表6)。、、和僅在19CD環境下檢測到,LOD值變化范圍為2.79~ 6.65,可解釋的表型變異率為4.48%~11.58%。僅在18GY環境下檢測到,LOD值為 3.62,可解釋的表型變異率是5.50%。僅在19GY環境下檢測到,LOD值為 3.39,可解釋的表型變異率為7.29%。除的加性效應來自母本Avocet S外,其他6個QTL的加性效應均來自父本貴協3號。
2.5 多態性引物的篩選結果
根據QTL定位結果和轉錄組數據,在2AS染色體的目標區段內設計了103對SSR標記引物,同時從GrainGenes數據庫上選擇了該區段內已報道的22對SSR標記引物。利用這125對標記引物對親本和抗、感池進行多態性標記篩選,最終獲得6個與抗條銹病位點緊密連鎖的SSR標記,分別為Xcfd36、Xwmc382、hls-2A-04、hls-2A-17、hls-2A-18和hls-2A-103,這些SSR標記在親本和抗、感池中的篩選結果如圖2所示。進一步在RIL群體中隨機選取40個株系材料進行驗證,發現這些標記均能較好地區分抗、感株系(圖3),因此可用于進一步的圖譜加密。
將6個多態性SSR標記在RIL群體中的基因分型結果與55K SNP芯片檢測結果整合,利用QTL IciMapping 4.0軟件重新進行連鎖分析,并用Mapchart V2.3.0重新繪制2AS染色體的遺傳圖譜(圖5)。綜合3個環境的結果表明,被定位到2AS染色體1.70~4.11cM的遺傳區間,標記區間為hls-2A-103~AX-108824773,對應的物理區間為15.87~18.91 Mb(3.04 Mb)。說明抗條銹病位點的定位區間從16.02 Mb縮小到3.04 Mb的物理區間內(表7)。
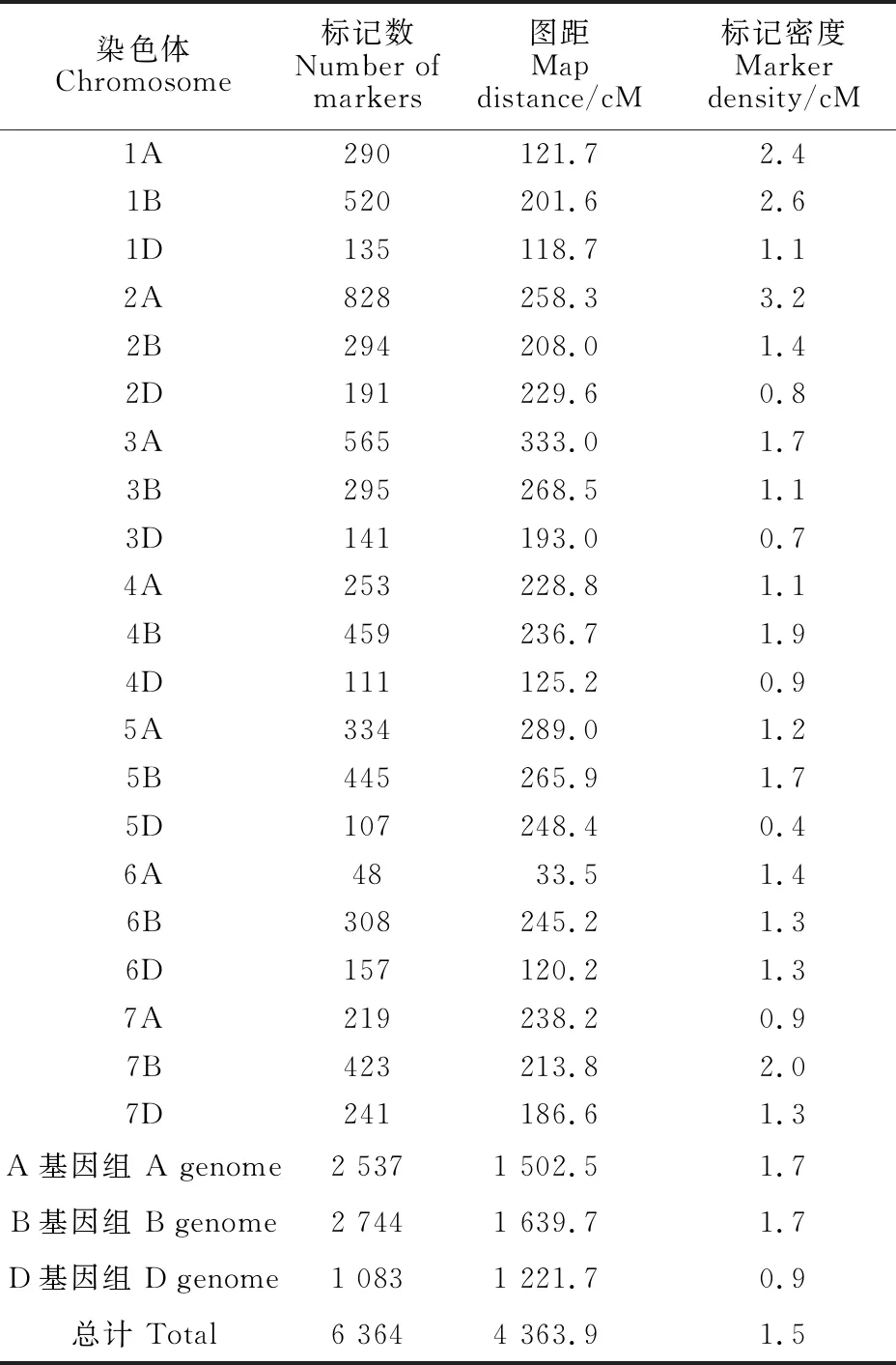
表5 RIL群體的21條染色體圖譜長度及SNP標記密度Table 5 Mapping length and SNP marker density on the 21 chromosomes of RIL population
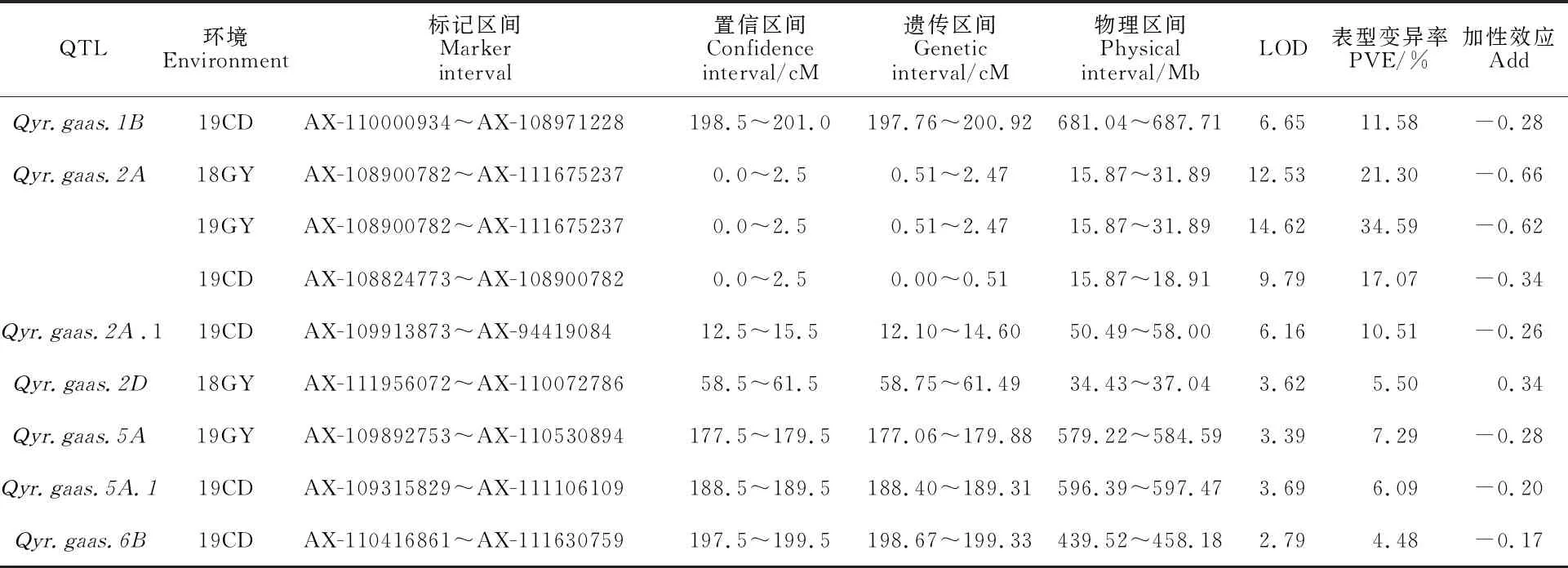
表6 3個環境下檢測到的抗條銹病QTLTable 6 QTLs for stripe rust resistance detected under three environments

G:貴協3號;A:Avocet S;R:抗病池;S:感病池。
2.6 候選基因預測分析
根據上述結果,貴協3號中的抗條銹病位點被定位于2AS染色體15.87~ 18.91 Mb的物理區間范圍內。利用小麥多組學網站與中國春2.0版本進行比對,識別小麥基因ID、注釋及相應序列。結果在上述區間內共預測到13個候選基因(表8),其中TraesCS2A02 G042400.1與抗病蛋白(NBS-LRR)相關,TraesCS2A02G040300.1與糖基轉移酶相關,TraesCS2A02G041200.1與谷胱甘肽S轉移酶相關。
3 討 論
3.1 抗條銹病基因的遺傳定位
本研究利用BSR-seq技術,將貴協3號抗條銹病位點初步定位于2AS染色體12.0~59.0 Mb的物理區間內(約47.0 Mb);而利用小麥55K SNP芯片則將定位到更小的物理區間(約16.02 Mb),該區間包含在BSR-seq的定位區間內,這說明55K SNP芯片技術的定位結果更加精確。
目前,在2AS染色體上已報道的抗條銹病基因有、、、和。其中,來源于硬粒小麥Wollaro,來源于美國軟紅粒小麥Pioneer 26R61。是從人工小麥滲入系CH7086中定位到的苗期抗條銹病基因,其抗性來源于彭提卡偃麥草。的抗病性來源于普通小麥品種中麥175。推測與、、和不同。來源于偏凸山羊草(),是一個苗期抗病基因,攜帶該基因的單基因系表現為高感,研究發現,載體品種Trident對CYR34小種表現為高感;但通過田間觀察發現,另一個載體品種VPM1對CYR34小種表現為中抗,推測VPM1中可能不僅含有,還含有其他未知的抗條銹病基因。Wang等報道,和也是基因的等位變異。為了明確和之間的差異,我們利用的特定CAPS標記URIC/LN2無法在貴協3號中擴增出標記目標條帶,所以推測與不同。為了進一步明確貴協3號中的與上述基因/QTL的相關性,還需要利用等位性檢測等手段進行判定。
3.2 候選基因預測分析
目前,已經克隆的多個小麥銹病基因均具有NBS-LRR結構,例如小麥抗條銹病基因和,抗葉銹病基因和,以及抗稈銹病基因和等。本研究根據所在2AS染色體末端 15.87~18.91 Mb的區間范圍,共預測到13個候選基因,其中TraesCS2A02G042400.1與抗病蛋白(NBS-LRR)相關,推測其可能是的候選目的基因。TraesCS2A02G040300.1與糖基轉移酶相關,糖基轉移酶在植物抗病過程中不僅可通過修飾植物激素來調節植物抗病能力,還是植物超敏反應抵御病害的必要因素。前人研究發現,UDP-糖基轉移酶(UGT)通過將小麥中的脫氧雪腐鐮刀菌烯醇(DON)糖基化為DON-3-葡萄糖苷(D3G),有助于增強赤霉病的抗性。值得注意的是,TraesCS2A02G041200.1與谷胱甘肽S轉移酶相關,而小麥抗赤霉病基因可編碼一種谷胱甘肽S-轉移酶(GST),所以這兩個基因也有可能是的候選基因。后期我們將繼續開發新的標記,擴大遺傳作圖群體,構建剩余雜合群體,實現抗條銹病位點的精細定位,進一步縮減目標區段,最終確定目標基因。
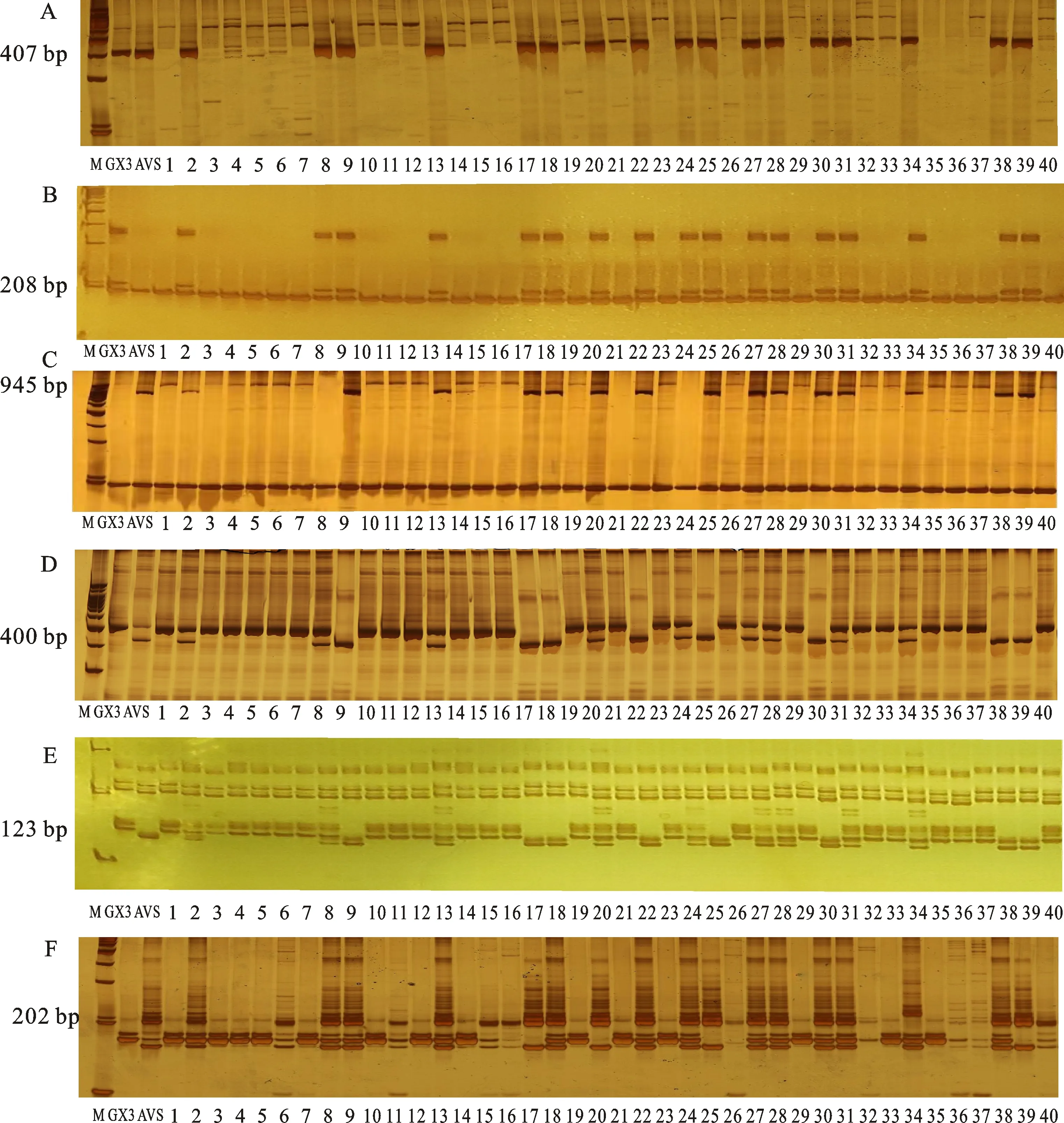
A~F分別代表6個多態性標記hls-2A-04、hls-2A-17、hls-2A-18、hls-2A-103、Xcfd36和Xwmc382,M代表標記DL2000,GX3代表貴協3號,AVS代表Avocet S,1~40代表RIL群體內隨機挑選的40個家系。
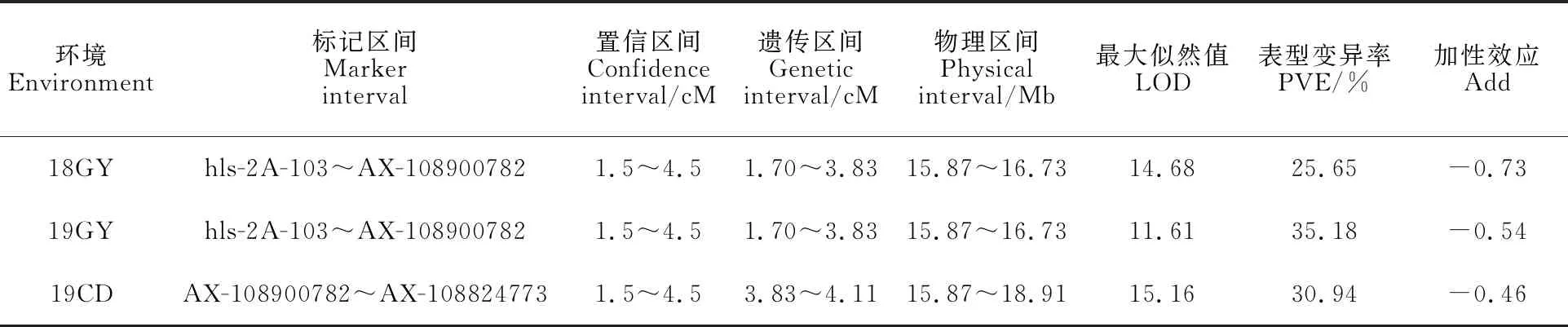
表7 利用開發的SSR標記重新定位 Qyr.gaas.2ATable 7 Relocation of Qyr.gaas.2A using the developed SSR markers
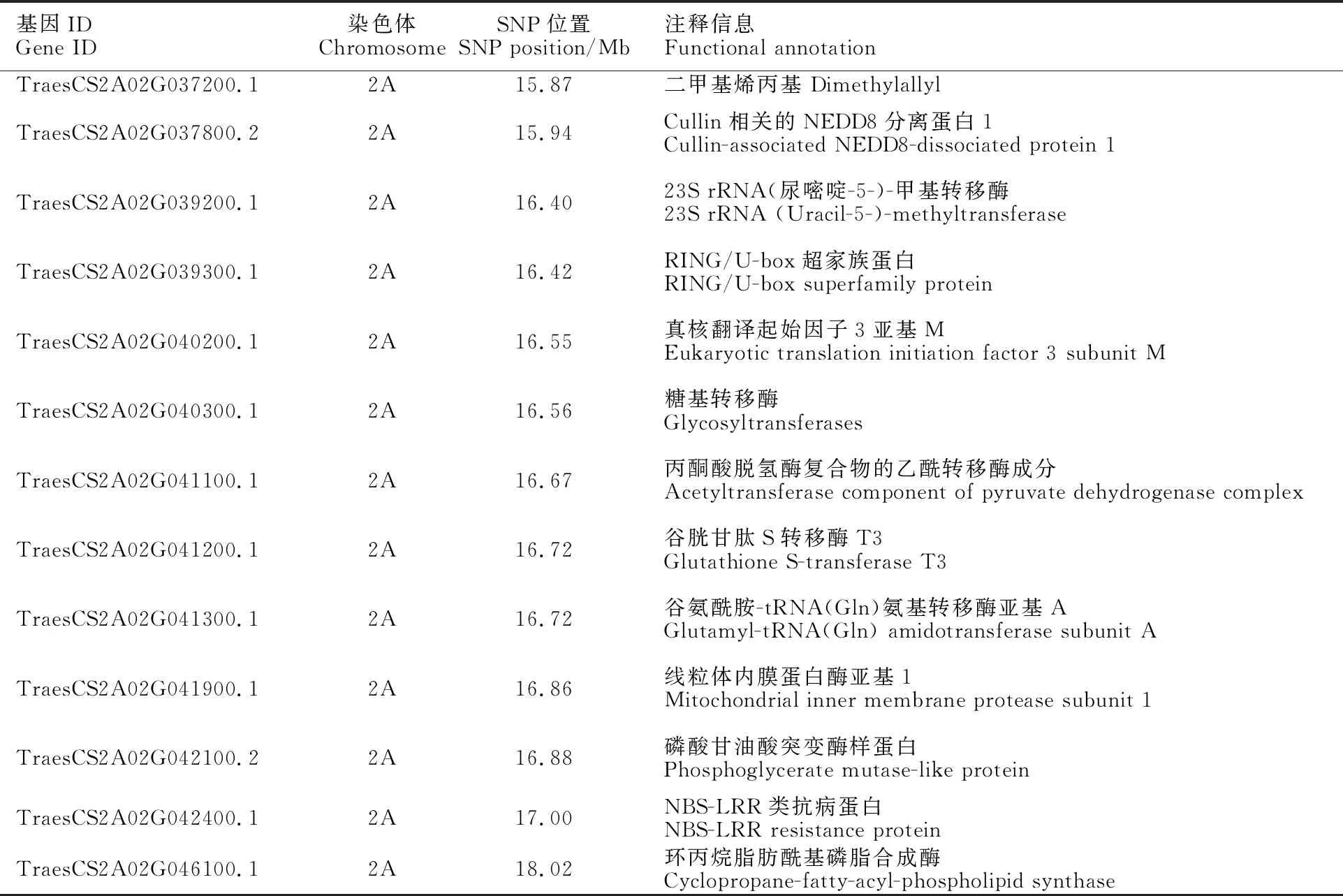
表8 候選區間內差異SNP的小麥基因功能注釋Table 8 Functional annotation of wheat genes with differential SNPs in candidate intervals
