基于基因芯片對癲癇小鼠海馬組織miRNA-204表達的研究及生物信息學分析
2022-08-09 01:38:44呂婷婷刁麗梅
中風與神經疾病雜志 2022年6期
吳 瓊, 呂婷婷, 刁麗梅
癲癇(Epilepsy)是由于大腦神經元異常放電引起短暫大腦功能失調的神經系統疾病,對患者學習和生活產生嚴重影響。全球每年有240萬人被診斷為癲癇,近30%的患者抗癲癇藥物無法控制癲癇發作,急需新的治療方法[1]。MicroRNA(miRNA)是一種長度為20~24個核苷酸的內源性非編碼蛋白的RNA,其通過與靶基因特定序列相互作用,調節靶基因表達的活性。miRNA異常的調節對癲癇的發生發展產生重要的影響,可能成為癲癇的潛在生物標記物和治療靶點。本研究利用基因芯片技術挖掘差異基因,并對差異基因進行生物信息學分析,探索miRNA對癲癇治療靶點作用機制,開拓癲癇診斷與治療新的思路。
1 材料與方法
1.1 材 料
1.1.1 實驗動物 雄性昆明小鼠20只,清潔級,約25~30 g。昆明小鼠由廣西醫科大學動物實驗中心購買。
1.1.2 實驗試劑和儀器 鹽酸匹羅卡品(美國Sigma公司)、氯化鋰(中國醫藥集團上海試劑公司)、硫酸阿托品注射液(鄭州鈴銳制藥有限公司)、Aglient 2100 Bioanalyzer(美國Agilent公司)、Thremo Nanodrop 2000(美國 Thremo公司)、GeneChip Scanner 3000(美國Affymetrix公司)、GeneChip Fluidics Station 450(美國Affymetrix公司)、GeneChip Hybridization Oven 645(美國Affymetrix公司)。
1.2 方 法
1.2.1 模型制備 將制昆明小鼠按隨機數字表法隨機分為造模組和對照組。造模組:昆明小鼠腹腔注射氯化鋰(127 mg/kg),18~24 h后注射阿托品 (1 mg/kg),30 min后腹腔注射匹羅卡品(10 mg/kg),每2 d進行造模實驗1次,共5次,Ⅳ級以上發作為造模成功。對照組:同樣方法注射生理鹽水。
1.2.2 實驗過程 分別取造模組和對照組小鼠腦部海馬組織,進行基因芯片檢測,選出模型組和對照組表達差異的基因。采用miRNA靶基因預測工具TargetScan、rna22、miRBase進行預測并取交集。利用DAVID數據庫,將取交集后靶基因基因按功能分為3大類:生物學過程、細胞組分和分子功能3類,后進行KEGG通路分析。
1.2.3 統計學分析數據 使用SPSS 21.0進行處理,組間比較使用t檢驗,P<0.05 為差異有統計學意義。
2 結 果
2.1 差異基因 根據實驗結果,篩選|FC|≥ 1.5的miRNA進行預測分析:相對表達增多的差異miRNA 10個,相對表達減少差異miRNA 18個,結合實驗結果及相關文獻,選取 miRNA-204進行預測分析。
2.2 靶基因預測 采用TargetScan共預測到miRNA-204靶基因6762個,rna22共預測到miRNA-204靶基因14335個,miRBase共預測到miRNA-204靶基因587個,取三者交集得到457個共同的靶基因。
2.3 靶基因GO分析 通過DAVID分析工具對miRNA-204靶基因進行分析發現,miRNA-204靶基因靶基因富集于21個基因功能(P<0.01)。miRNA-204靶基因富集在生物學過程:大腦發育、磷脂酰肌醇代謝過程、離子運輸、蛋白質運輸、蛋白質硫酸化、類固醇激素介導的信號傳導、線粒體去極化、細胞對缺氧的反應、GTP酶介導的信號轉導、肌動蛋白細胞骨架組織等相關生物學過程。miRNA-204靶基因主要作用在:突觸后膜、高爾基體膜、細胞核、細胞質內等細胞組分。miRNA-204靶基因富集的分子功能包括:與蛋白質結合和金屬離子結合、調控類固醇激素受體活性及GTPase活化劑活性(見圖1)。
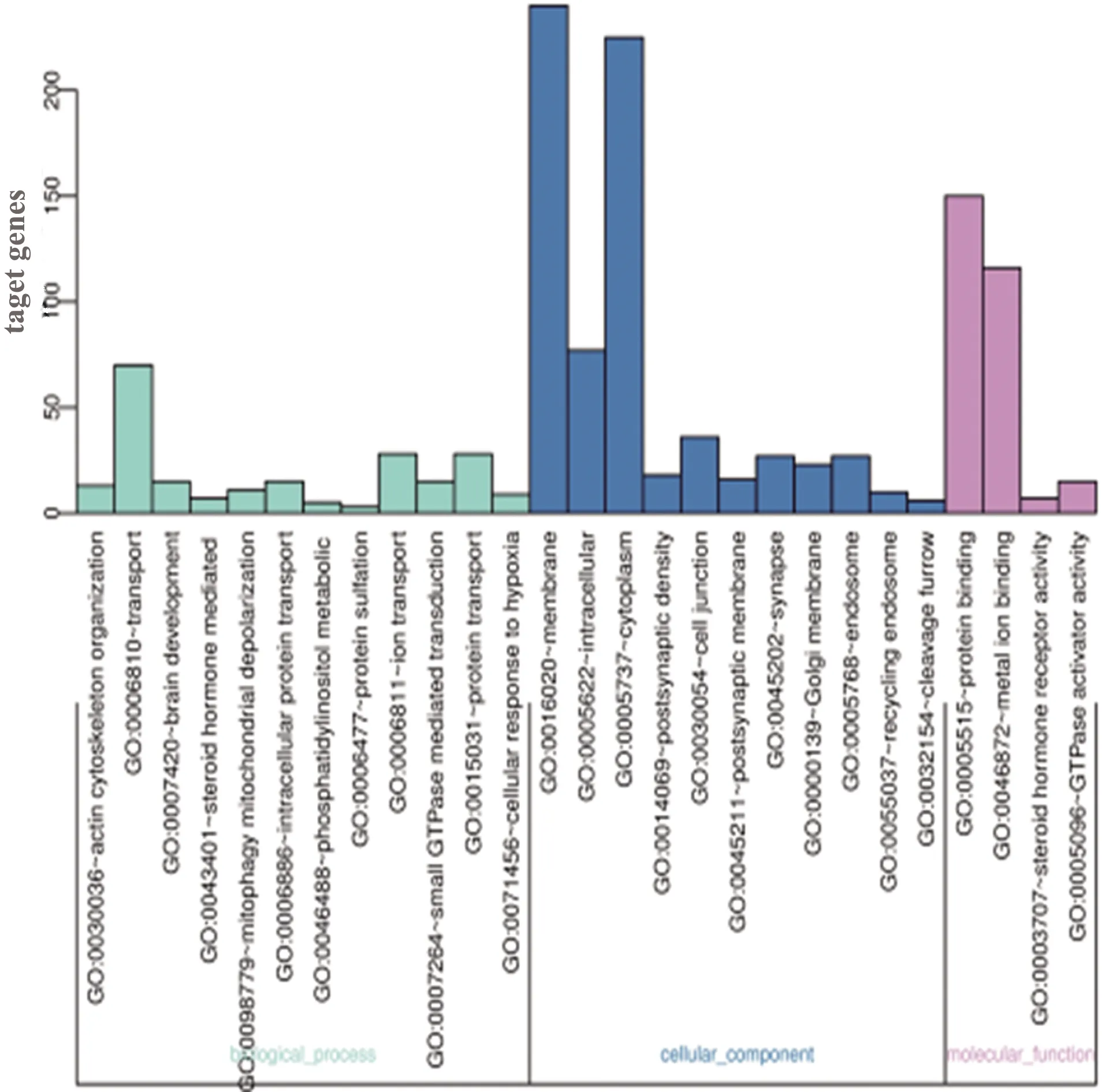
圖1 miRNA-204靶基因富集分析
2.4 miRNA-204靶基因KEGG分析 miRNA-204靶基因主要富集在16條通路中(P<0.05):Wnt信號傳導通路、磷脂酰肌醇信號系統、神經活性配體-受體相互作用通路、鞘脂信號轉導通路、鈣信號通路、Hippo信號通路、cGMP-PKG信號傳導通路、癌細胞膽堿代謝、血管平滑肌收縮、晝夜節律、胰腺分泌、唾液分泌等相關通路(見圖2)。
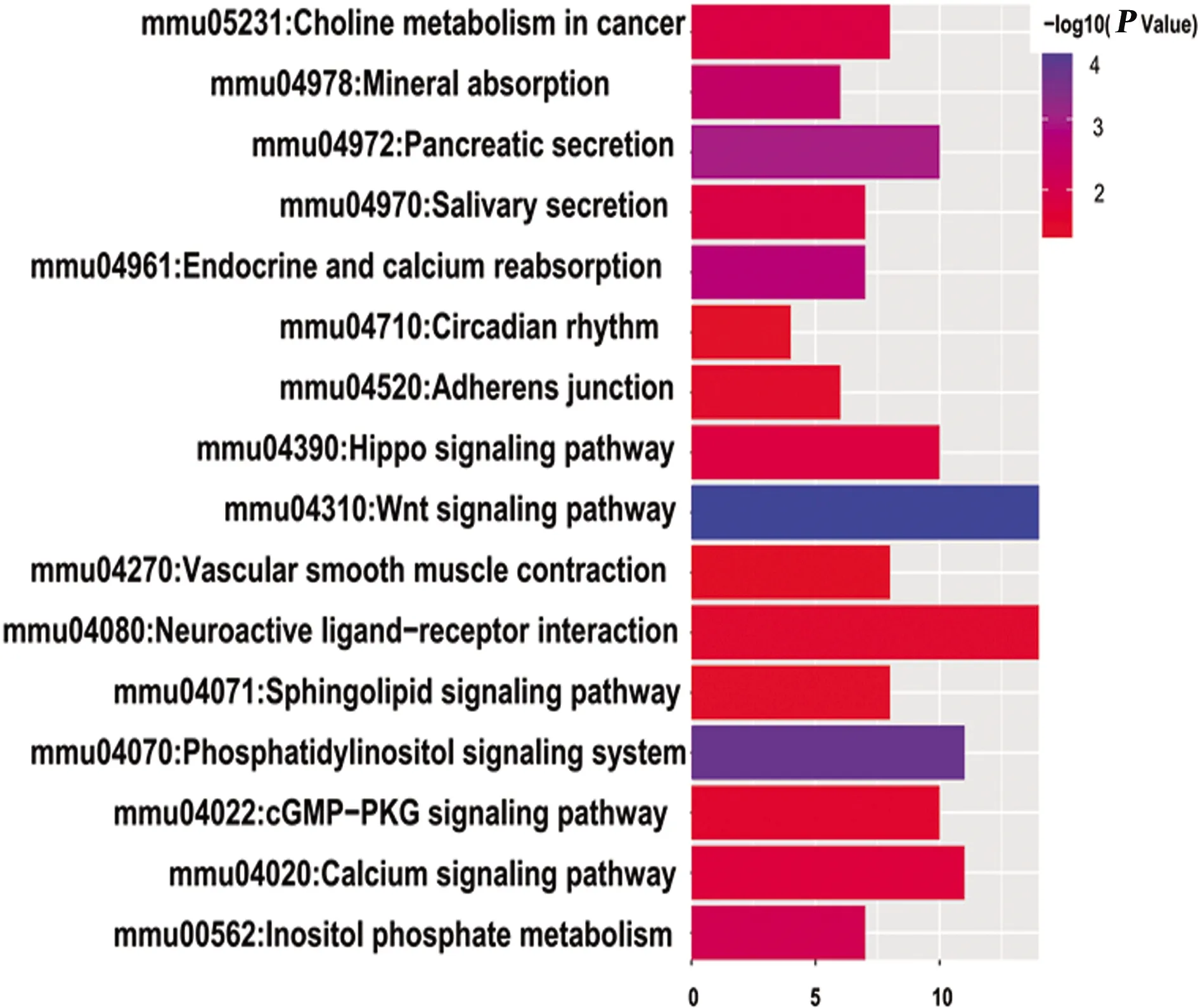
圖2 miRNA-204靶基因KEGG分析
3 討 論
本實驗研究利用基因芯片發現miRNA-204在氯化鋰-匹羅卡品癲癇小鼠海馬組織表達減少。已有研究報道與本實驗基因芯片檢測結果相一致:Raoof R等[2]研究發現,miRNA-204在顳葉癲癇及癲癇持續狀態患者的腦脊液中表達顯著減少,Xiang等[3]研究也顯示在體外培養的癲癇持續狀態海馬神經細胞中miRNA-204顯著下調,miRNA-204 通過調節 TrkB 和下游ERK1/2-CREB信號通路在海馬神經元模型中發揮抗癲癇作用。在Xia等[4]在膠質細胞瘤繼發的癲癇腦組織中發現,miRNA-204表達水平明顯低于正常腦組織的表達量,并且發現miRNA-204可以顯著抑制了膠質瘤細胞的生長、遷移和侵襲。
本實驗研究中預測的miRNA-204靶基因的功能GO富集顯示:miRNA-204與大腦發育關系密切,相關研究也證實miRNA-204在神經發育過程中起關鍵作用,包括在軸突誘導控制中的作用和在神經系統發育中的重要作用[5]。miRNA-204在驚厥性腦損傷后表達顯著增加,可減少驚厥誘導的神經細胞凋亡,加速神經細胞軸突的生長與延伸[6]。然而,一些研究也提示miRNA-204可導致大腦海馬神經元突觸可塑性下降,影響海馬組織調控的認知功能改變。隨著年齡增長,海馬中miRNA-204表達上調,而EphB2和NR1表達降低,miRNA-204可通過調控EphB2和NR1影響海馬突觸可塑性下降,可能導致年齡相關認知功能的改變[7]。在KEGG通路分析顯示miRNA-204靶基因富集于Wnt信號通路的有:AF366264、VANGL1、BTRC、PRKCG、FZD3、DAAM1、DAAM2、WNT2、MAP3K7、RAC2、SFRP1、PRICKLE2、PLCB1、FBXW11。
經典Wnt/β-catenin通路調節海馬神經發生、突觸分裂和線粒體調節,對中樞神經系統的發育和功能至關重要[8]。相關的研究發現,β-catenin在對低氧發作反應中出現巨大的轉錄變化,β-catenin在神經元結構的形成和維持以及促進突觸可塑性方面具有重要作用[9]。另一項研究通過使用Wnt通路抑制劑,減少了紅藻氨酸誘導小鼠癲癇發作次數,顯示了Wnt通路治療癲癇潛在有效性[10]。Wnt信號通路參與癲癇急性期和慢性期,誘發的神經發生和神經元死亡[11],急性期癲癇發作時Wnt信號通路激活,導致了短暫的神經代謝的改變,隨著癲癇發作的持續,Wnt信號通路將激活mTOR通路,導致癲癇慢性發作。癲癇發生的早期有一個治療窗口,Wnt信號通路啟動了短暫的代謝,這一系列的代謝重組變化可能影響神經異常發生和分化,從而影響癲癇敏感性[12]。同時值得注意的是,長鏈非編碼RNA Uca1可以通過降低miRNA-375的水平來促進Sfrp1的表達,進而通過抑制Wnt/β-catenin通路的激活,抑制癲癇小鼠神經元細胞的異常增殖,預防癲癇發作[13]。長鏈非編碼RNA Peg13可通過miRNA-490-3p/Psmd11軸抑制Wnt/β-catenin通路,減緩小鼠癲癇進展[14]。
癲癇是最常見的神經系統疾病之一,在臨床中有許多患者對目前所有可用的治療藥物都有一定的耐藥性。現有的抗癲癇藥物治療主要是減少興奮和預防癲癇發作,但不能影響疾病的潛在病理生理。最近,有許多研究工作集中在確定抗癲癇治療的機制靶點,以防止慢性癲癇的進展。在本實驗中預測miRNA-204的靶基因富集功能和KEGG通路部分已經被證實,其中Wnt/β-catenin信號通路可能是未來抗癲癇治療的十分有前景的癲癇治療靶點。本課題組將在以后的研究中對miRNA-204靶基因和Wnt/β-catenin信號通路完善相關實驗研究,進一步驗證其對癲癇發生發展的影響。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
中國民間療法(2021年5期)2021-06-09 09:21:04
學苑創造·A版(2020年9期)2020-10-13 09:41:02
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
飲食科學(2017年5期)2017-05-20 17:11:53
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
作文與考試·小學高年級版(2015年17期)2015-05-30 10:48:04
西南軍醫(2015年4期)2015-01-23 01:19:30
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
