三峽水庫支流浮游植物演替內外驅動力辨析
2022-08-10 08:07:46陳佳俊張佳磊
湖北農業科學 2022年13期
陳佳俊,張佳磊
(湖北工業大學土木建筑與環境學院/河湖生態修復與藻類利用湖北省重點實驗室,武漢 430068)
三峽蓄水成庫后,因改變三峽水庫的水動力條件,從而影響三峽水庫支流的水溫結構、光照條件和營養鹽遷移轉化規律[1-4],導致水體中浮游植物群落特征發生一系列的改變,由河流型硅甲藻向湖泊型藍綠藻水華演替[5],而多數藍藻在代謝過程中分泌的藻毒素破壞了生態環境,對飲用水源安全產生嚴重影響,引起國內外的廣泛關注[6-8]。
浮游植物是水生態系統中的初級生產者,其群落結構能反映水質狀況的好壞[9]。浮游植物群落的變化受到水溫、光照、營養鹽以及水動力條件的影響[10],水溫對藻類生長速率有一定影響,不同水溫下浮游植物的種類存在顯著差異[11]。不同水文期的水體光學特性存在顯著差異,真光層深度以及光合有效輻射的變化影響浮游植物的生物量[12]。水體變緩,顆粒物攜帶營養鹽沉積效應加強,營養鹽在支流庫灣中的滯留,浮游植物生物量升高[13]。河流、水庫等流動性水體,其浮游植物生境的主要參量是水動力條件,已有研究表明水華暴發與水動力條件呈顯著相關[14-17]。綜上所述,三峽水庫的浮游植物演替是外部環境條件改變所引起的。
但是從生態學角度來看,浮游植物的演替過程是內部競爭與外部環境條件共同驅動下發生的。目前多關注外部因素,而對內部因素效果關注較少,根本原因是內部因素不好分辨。趨同性的核心是指浮游植物群落演替由環境條件所決定;在相同的環境條件下,浮游植物群落將向同一方向進行演替。不同環境條件下,浮游植物群落將向不同的方向進行演替。趨異性是指浮游植物群落的演替由群落的初始結構所決定;在相同的環境條件下,由于初期群落結構的不同,也將向不同的方向演替[18]。為明確三峽庫區浮游植物演替的內外驅動力因素,本研究基于浮游植物群落構建的趨同性和趨異性理論,開展了溫度、光照對浮游植物群落影響的室內控制試驗,以期探究影響河道型水庫浮游植物群落生物量與多樣性的關鍵因子。
1 材料與方法
1.1 藻種來源與試驗裝置
藻種來源為香溪河河水。光照系統采用日光燈組連續提供,確保光照強度分布均勻且穩定在(2 000±150)lx;水溫用可調式絕緣電熱棒控制,精確度為±1 ℃。試驗裝置如圖1所示,將9個長50 cm、寬38 cm、高30 cm 的白色水框平行布置,放置于自然條件下。在每個水框中均勻放入2 個微型水泵,用以擾動水體,避免水體中的藻類自然沉降。微型水泵通過硅膠軟管與空氣壓縮機連接;進氣前端設有孔徑為0.2 μm 的空氣過濾器用以除塵除菌,另設置空氣開關閥用以控制進氣量的大小。
1.2 試驗方法
設有3 個試驗組,分別為空白對照組、溫度組和光照組。每組設3 個平行,共9 個白色水框。空白對照組為自然條件下的光照及溫度;溫度組為自然條件下的光照,溫度設定為26 ℃;光照組為自然條件下的溫度,光照設定為2 000 lx。
監測時段為2016 年5 月4—26 日,采樣頻率為前14 d 每天采樣1 次,之后每3 d 采樣1 次,共17 次。為了解試驗前各水框的初始狀況,試驗初始測定每個水框中藻類的初始群落結構,以及每個水框中TN、TP 濃度,之后每3 d 測定1 次,滿足藻類生長所需的氮、磷營養鹽閥值(TN>0.2 mg/L,TP>0.02 mg/L)。24 h 不間斷曝氣,促進營養物質循環,盡量避免藻類沉降。
1.3 測定指標及方法
試驗開始后,每天早上09:00 左右取樣。光照強度和水體溫度分別用TES-1339 照度計(臺灣泰仕)和玻棒煤油溫度計(河北光華)測定。TN、TP 濃度利用UV-1700 紫外分光光度計(上海美析)進行測定[19],藻類群落結構利用AlgacountS300 藻類智能鑒定計數儀(杭州迅數)進行鑒定和計數。藻類種類根據文獻[20,21]進行鑒定,藻細胞計數和生物量測算根據文獻[22]。藻種統計至屬即可完全反映藻類群落特征及演替過程,因此本研究藻類鑒定數據統計到屬[23]。
1.4 數據處理
1.4.1 生物量

式中,N為1 L 水樣中浮游生物的數量,ind/L;Cs為框面積,mm2;Fs為視野面積,mm2;Fn為每片計數過的視野數;V為1 L 水樣經濃縮后的體積,mL;V0為計數框容積,mL;Pn為計數的浮游植物個數。
按上述方法進行采樣、稀釋、計數。Cs為400 mm2,Fs為0.156 549,Fn為100,V為1 000 mL,V0為0.10 mL。因此只要計數方法確定,就可以得到N。
1.4.2 演替速率 通常在生境穩定條件下,群落演替的結果是達到頂極群落(Climax)。演替速率是群落結構改變潛勢的客觀測度。Lewis(1978)修正了Jassby and Goldman(1974)建立的群落演替速率計算方法,并在藻類群落生態研究中廣為采用,其計算公式如下。
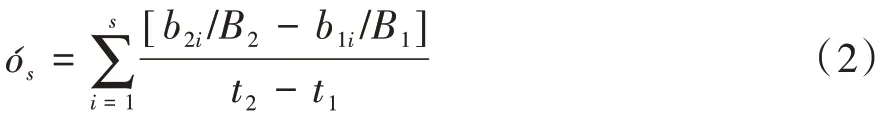
式中,B2、B1分別為群落演替過程中2 個時間狀態(t2、t1)的藻類生物量,而b2i、b1i則為上述相應狀態下群落中第i個藻種的生物量。
1.4.3 多樣性指數 常用的多樣性指數[23-25]有Mar?galef 指 數、Shannon-Winener 指 數 和Jaccard 指 數。Margalef 指數主要反映群落物種的豐富度,浮游植物種類愈多,S越大。Shannon-Winener 指數反映群落結構的復雜程度,是衡量種類數和均勻度的綜合指標。Jaccard 指數反映樣點間浮游植物群落結構的相似性,J愈大,表明樣點間的浮游群落結構愈相似。
1)Margalef豐富度指數(S)。

2)Shannon-Winener 指數(H)。

3)Jaccard 種類相似性指數(J)。

式中,S為群落中物種數,Pi=Ni/N;Ni為第i種藻的個數,N為觀察到的總個數。J為相似性指數,a為樣點A 的種類數;b為樣點B 的種類數;c為樣點A 和B 共同的種類數。當0<J<0.25 時,樣點A 和B 的浮游植物群落結構達到極不相似水平;當0.25≤J<0.50 時,樣點A 和B 的浮游植物群落結構達到中等不相似水平;當0.50≤J<0.75 時,樣點A 和B 的浮游植物群落結構達到中等相似水平;當0.75≤J<1.00時,樣點A 和B 的浮游植物群落結構達到極相似水平[26]。
1.4.4 數據分析 數據分析利用SPSS 17.0 統計軟件,采取單因素ANOVA 均值分析方法。P<0.05 為顯著,P<0.01 為極顯著。
2 結果與分析
2.1 不同環境條件下藻類群落演變特征
2.1.1 藻類細胞生物量 在試驗期間,對照組、溫度組和光照組的藻類細胞密度存在顯著差異(ANO?VA,P<0.05)。對照組、溫度組和光照組時間變化趨勢一致,均呈先上升后下降趨勢,但是其達到峰值的時間略有差別,分別為5 月13 日、5 月10 日和5 月9日。對照組、溫度組和光照組在試驗期間的生物量平 均 值 為(105.8±82.5)×105、(109.8±90.1)×105、(402.5±289.6)×105ind/L,光照組藻類細胞生物量顯著大于對照組和溫度組(圖2)。
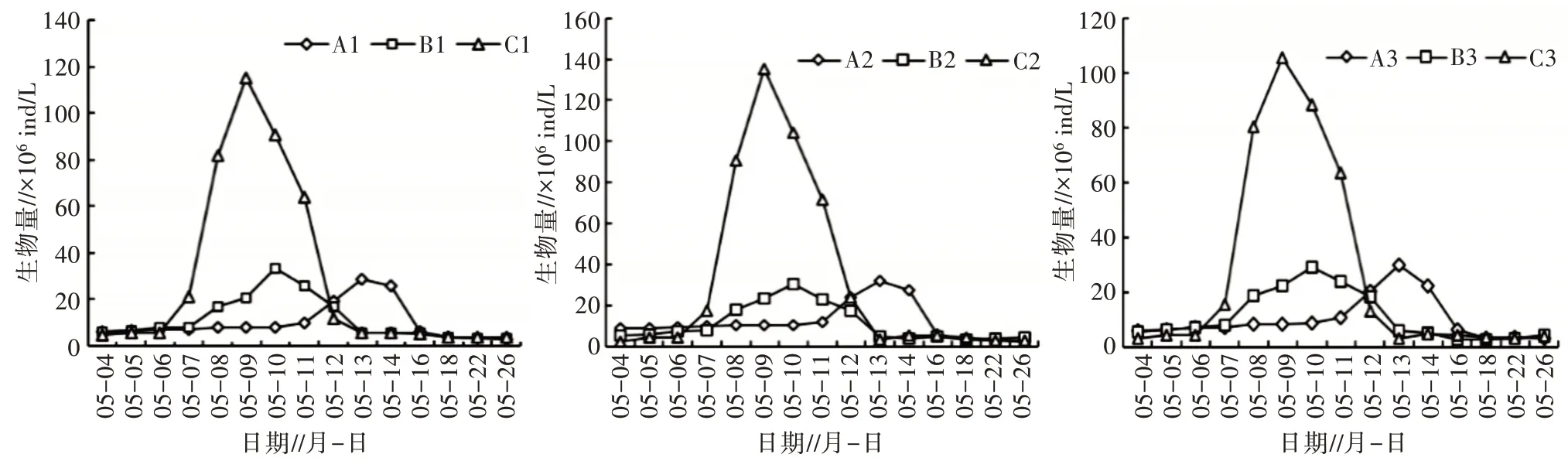
圖2 不同環境條件藻類細胞生物量變化
2.1.2 藻類的群落結構特征
1)不同環境條件藻類群落特征。根據形態鑒定,試驗期間共鑒定出浮游植物27 種,隸屬于4門[20]。其中種類最多的是綠藻門(Chlorophyta)(12種),其次是硅藻門(Bacillariophyta)(9 種)、藍藻門(Cyanophyta)(4 種)和裸藻門(Euglenophyta)(2 種)。
不同的光照和溫度條件對浮游植物群落的演替有明顯的影響,試驗組的優勢種存在一定的差異性。初始培養期,光照組、溫度組、對照組的浮游植物群落結構均由硅藻、藍藻、綠藻和裸藻組成。從初始培養到培養穩定后優勢藻類的種類明顯減少,其中,小環藻屬(Cyclotella)是光照組最主要的優勢種,溫度組主要優勢種是衣藻屬(Chlamydomonas)和小球藻屬(Chlorella),對照組的優勢藻類為微囊藻(Micro?cystis)和色球藻(Chroococcus)(表1)。

表1 各試驗組的藻類演替規律
2)不同環境條件藻類演替速率比較。演替速率是指在某一時間段內藻類群落的演替速率。根據表2 可以看出,光照組的藻類演替速率最快,溫度組次之,對照組生長最為緩慢。所以光照和溫度條件對藻類群落結構的改變有一定影響。
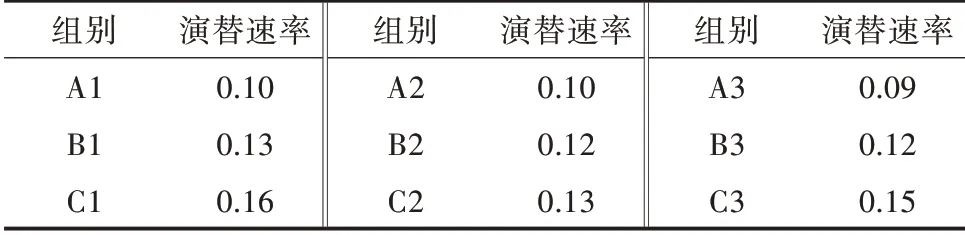
表2 不同環境條件下的演替速率
3)藻類群落結構相似性比較。由圖3 可以看出,試驗期間不同環境條件下試驗組的浮游植物相似性指數在0.27~0.67。試驗初期,各試驗組的浮游植物相似性指數在0.55~0.60,其中對照組與光照組的相似性指數在0.30~0.43,平均值為0.48±0.03,為中等不相似水平;對照組與溫度組、光照組與溫度組群落結構平均值分別為0.54±0.04、0.55±0.05,達到中等相似水平。5 月26 日浮游植物到達演替后期,對照組與溫度組、光照組,溫度組與光照組的浮游植物相似性指數介于0.36~0.45,其種類相似性水平平均值分別為0.39±0.03、0.39±0.01、0.40±0.04,為中等不相似水平。
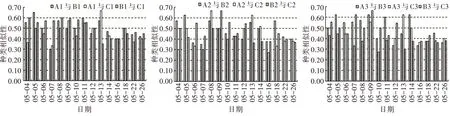
圖3 不同環境條件Jaccard 種類相似性指數變化
4)藻類的物種豐富度及多樣性指數。光照組與溫度組具有極顯著差異(P<0.01),光照組與對照組呈顯著差異(P<0.05),溫度組與對照組差異不顯著(P>0.05)。從藻類的Shannon-Winener 多樣性指數分析結果(圖4)可以看出,各試驗組的多樣性指數呈波動下降后上升趨勢;對照組、溫度組、光照組的平均值分別為0.97±0.50、0.91±0.54、1.27±0.36,藻類群落多樣性大小為C>A>B。

圖4 不同環境條件Shannon-Winener 多樣性指數變化
從浮游植物的Margalef 豐富度指數分析結果(圖5)可以看出,光照組、溫度組和對照組的Mar?galef 豐富度指數分別為0.13~0.72、0.13~0.51、0.12~0.62,其平均值分別為0.39±0.12、0.28±0.09、0.29±0.09,光照組物種豐富度均在5 月12 日達到最大,在5 月8—26 日維持穩定,不同試驗組間的物種豐富度差異顯著(P<0.05)。

圖5 不同環境條件Margalef豐富度指數變化
2.2 相同環境條件下藻類群落演變特征
2.2.1 藻類細胞生物量 3 個平行組之間的藻類細胞密度不存在顯著性差異(P>0.05),由圖6 可以看出,相同環境條件下的平行組整體變化趨勢一致,隨時間的推進均呈先上升后下降的趨勢。
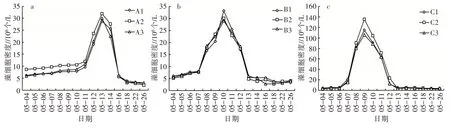
圖6 相同環境條件藻類細胞生物量變化
2.2.2 藻類的群落結構特征
1)藻類群落鑒定結果及演替速率。分別對相同條件各平行組藻類群落形態鑒定(表3),結果表明,穩定期光照組的3 個平行組優勢種(優勢藻以超過總生物量10%為評定標準)均為小環藻屬(56.35%,58.24%,55.69%)。溫度組的3 個平行組優勢種均為衣 藻 屬(58.22%,56.25%,60.36%)和 小 球 藻 屬(36.17%,40.05%,35.14%)。由于對照組3 個平行處于自然條件下,環境因子干擾,其對浮游植物演替不顯著,導致浮游植物的內部競爭更為明顯[18]。因此,對照組的3 個平行組優勢種不一致,A1與A3均為微囊藻屬(40.67%,38.50%),色球藻屬(20.67%,19.16%);A2為 微 囊 藻 屬(42.58%)、小 球 藻 屬(30.61%)。
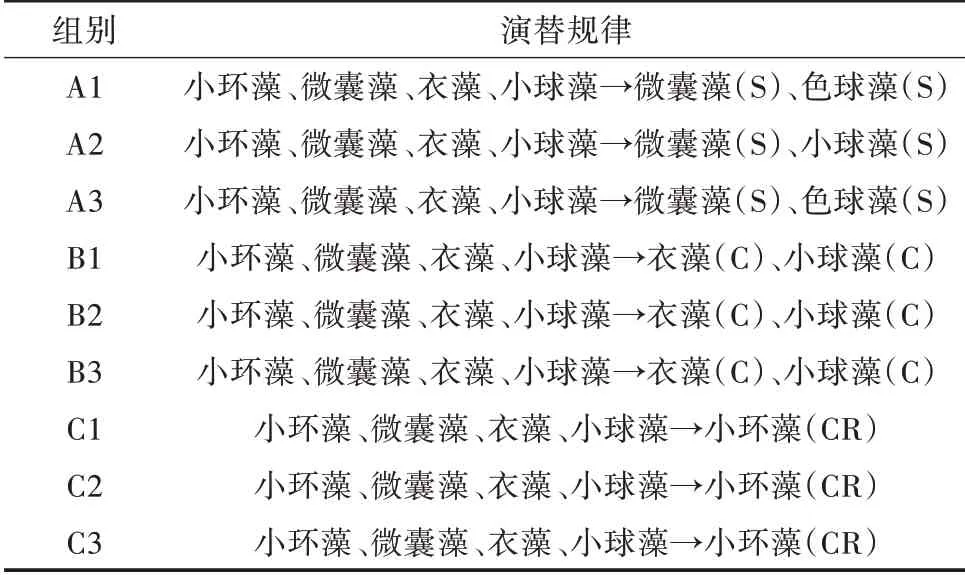
表3 相同條件各平行試驗組藻類演替規律比較
2)相同環境條件下藻類群落相似性比較。由相同環境條件Jaccard 種類相似性指數變化(圖7)可以看出,對照組、溫度組和光照組的初始浮游植物相似性指數分別在0.56~0.71、0.36~0.50、0.43~0.57,其平均值分別為0.62±0.08、0.68±0.16、0.57±0.12。在浮游植物達到峰值時,浮游植物相似性指數介于0.50~0.83,浮游植物群落達到中等相似水平。5 月26 日浮游植物演替后期,各試驗組的浮游植物相似性指數在0.67~1.00,對照組、溫度組和光照組的種類相似性平均值分別為0.78±0.05、0.91±0.08、0.88±0.06,達到中等以上相似水平。
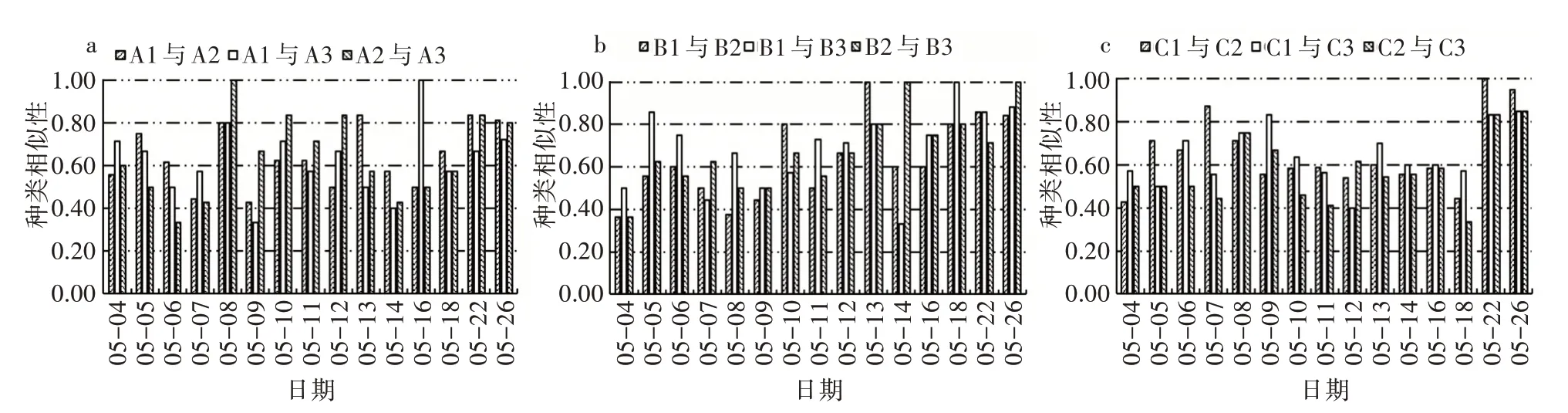
圖7 相同環境條件Jaccard 種類相似性指數變化
3)藻類的物種豐富度及多樣性指數。相同環境下,平行組的藻類多樣性指數無顯著差異(ANOVA,P>0.05)。相同環境下各水框中浮游植物群落的Shannon-Winener 指數與Marglef 豐富度指數(圖8、圖9)隨時間的變化規律相似。各平行試驗組藻類多樣性指數呈相同的變化趨勢,總體上表現為先減小后增大的變化規律。光照組、溫度組和對照組分別為0.50~2.07、0.09~1.83、0.23~2.22,其中光照組在5 月8 日多樣性指數最小,在5 月18—26 日趨于穩定;溫度組在5 月10 日多樣性指數最小,在5 月16—26 日浮游植物多樣性指數趨于穩定;對照組A1與A2的多樣性指數呈先增大后減小再增大的變化規律,A3表現為先減小后增大的變化趨勢。
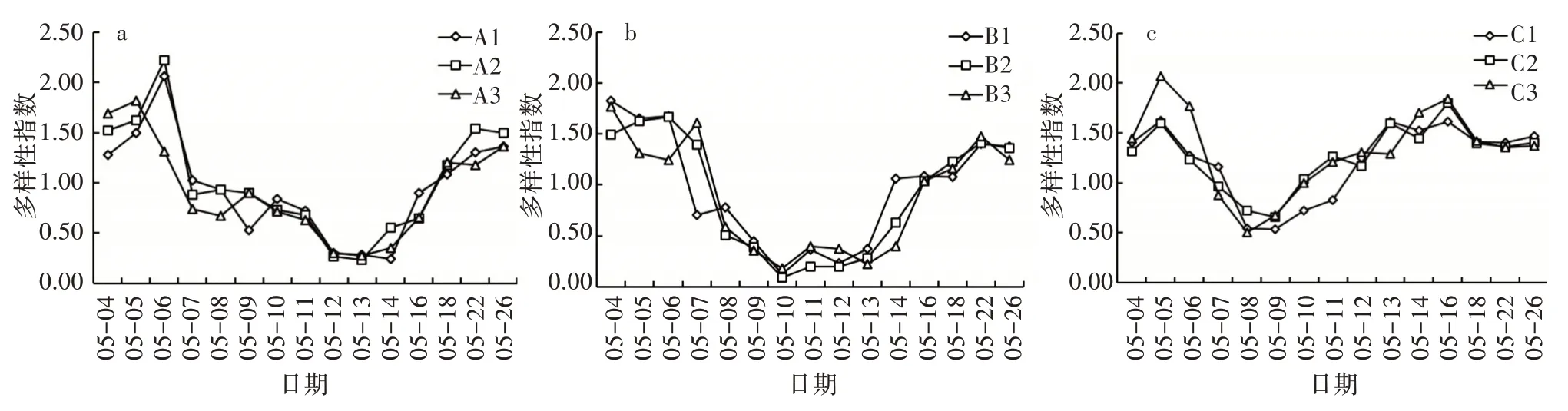
圖8 相同條件下Shannon-Winener 多樣性指數變化

圖9 相同條件下Margalef豐富度變化
3 討論
3.1 環境因子對浮游植物群落結構的影響
試驗期間,各試驗組水體處于富營養化水平(TN>0.2 mg/L,TP>0.02 mg/L)。鑒于浮游植物自身含有營養鹽,氮、磷、營養鹽含量與浮游植物生物量的變化有很強的自相關性,并且各試驗組水體為富營養化水體,因此營養鹽對各試驗組浮游植物群落結構變化的影響不是主要因素,驅動浮游植物群落改變的主要外界因素是溫度和光照[27]。
浮游植物群落演替表征為生境變化下,不同浮游植物隨時間的變化呈相互取代,是自然界發生且可辨識的[28]。在富營養化水體中,浮游植物生物量的增加將導致其群落多樣性的下降[29,30]。浮游植物在適宜的生境下往往具有較高的生長速率,從而生物量能在充足的時間內得到積累而形成優勢;不適浮游植物的生境將因其生長速率受到抑制,在此生境下不具備優勢,而使其被其他浮游植物所取代[10]。在不同環境下3 個試驗組的浮游植物生物量在不同時間節點達到峰值,并且其藻類生物量大小相差較大。光照組最先達到峰值,溫度組次之,對照組最后達到峰值。在浮游植物生物量達到最大時,光照組的優勢種演替為小環藻,溫度組的優勢種演替為衣藻,對照組的優勢種演替為微囊藻。在試驗后期,光照組的優勢種均為小環藻;溫度組的優勢種均為衣藻;對照組光熱條件較差,浮游植物的生長受到光熱條件限制較為明顯,具有低光強的微囊藻、小球藻以及色球藻在該生境條件下具備生存優勢,在試驗期間演替為優勢種。但是當外界環境因子對浮游植物演替不顯著時,浮游植物的內部競爭更為明顯[18]。導致其3 個平行的優勢種不一致,A1與A3組為微囊藻、色球藻;A2組為微囊藻、小球藻。不同環境條件下Jaccard 種類多樣性指數達到中等不相似水平,Margelef 豐富度指數與Shannon-Winener 多樣性指數隨時間的變化呈顯著差異(ANOVA,P<0.05)。由此可見,不同環境條件下浮游植物群落演替表現為趨異性。
在相同環境下,組間浮游植物演替具有相似的變化趨勢,浮游植物群落的構建表現為趨同性。3個試驗組的浮游植物生物量均在同一天達到峰值,其生物量較接近。Jaccard 種類相似性達到中等以上相似,Margelef 豐富度指數與Shannon-Winener 多樣性指數(ANOVA,P>0.05)隨時間的變化無顯著差異。根據以上比較可以看出,外界的環境因子對浮游植物群落演替的驅動力較大。
3.2 基于生態學理論辨析三峽水庫浮游植物群落結構演替驅動力因素
水生環境浮游植物生物量和群落結構的改變取決于物理(水文,水動力條件)、化學(營養鹽)以及生物因素(浮游動物捕食作用,浮游植物內部競爭)的相互作用[10,31-33]。不同湖庫的浮游植物群落結構演替因素有所區別,有研究表明,水溫、光輻射強度以及營養鹽濃度是淺水湖泊產生群落結構演替的主導因子[34,35]。對于深水型水庫的調查結果顯示,水體下光熱結構(光照和水體層化模式)、水體滯留時間、外源營養鹽輸入等環境因子的改變導致浮游植物群落結構的演替[36,37]。三峽水庫的特殊水動力學造成了支流水生生境的改變,有研究表明,水位波動、水溫、光熱結構和水體穩定系數是改變三峽水庫支流浮游植物群落結構的關鍵因素[38-41]。三峽水庫支流的浮游植物群落結構演替的驅動力主要是外界的環境因子。
根據“生態學理論”,Reynolds[10]認為浮游植物生境特征與物質、能量的供給相關,物質供給主要為浮游植物對水體營養鹽的利用程度,能量供給則為溫度和光照2 個方面。在穩定的環境中,水體中的浮游植物群落會相互競爭,導致群落多樣性下降,生物量達到最大,演替速率達到最低。對于外界環境條件影響較大的光照組和溫度組生物量相繼達到峰值,此時的群落多樣性指數達到最低值。
4 結論
不同外界環境條件下,光照組浮游植物群落結構演替速率最快,相比于溫度組和對照組,其生物量具有明顯的升高趨勢,光照條件對浮游植物群落演替的驅動更為顯著。 不同試驗組的優勢種呈現出一定的差異性。浮游植物在生物量達到峰值時,光照組的群落結構以小環藻占優,溫度組的優勢種為衣藻和小球藻,對照組的優勢種為微囊藻和色球藻,在外界環境因子影響較為顯著時,浮游植物群落結構向同一方向演替。
