新型蛋白互作識(shí)別系統(tǒng)的研究進(jìn)展
2022-08-10 08:08:26于海波
湖北農(nóng)業(yè)科學(xué) 2022年13期
于海波
(天津大學(xué)生命科學(xué)學(xué)院,天津 300072)
蛋白質(zhì)之間的相互作用(Protein-protein interac?tion,PPI)對于許多生物過程至關(guān)重要,已得到了廣泛研究[1]。目前,已經(jīng)開發(fā)出多種傳統(tǒng)的PPI研究方法,這些方法各具優(yōu)勢,但也有其自身的局限性。為了克服傳統(tǒng)PPI 的局限性,近年來開發(fā)出了幾種新型PPI 識(shí)別系統(tǒng),這些系統(tǒng)能夠高效地應(yīng)用于PPI 研究。本研究綜述了傳統(tǒng)PPI 研究方法的優(yōu)勢及其局限性、新型PPI 識(shí)別系統(tǒng)的設(shè)計(jì)以及新型PPI 識(shí)別系統(tǒng)的特點(diǎn),以期為開發(fā)更多高效的PPI 識(shí)別系統(tǒng)提供幫助。
1 傳統(tǒng)PPI研究方法
免疫共沉淀可用于大規(guī)模篩選,然而,這種技術(shù)卻不能區(qū)分直接的相互作用和間接的相互作用[2-4]。酵母雙雜交技術(shù)是一種強(qiáng)大的檢測PPI 的方法,其可以用于全基因組篩選,并有潛力識(shí)別相對較弱的PPI,然而,酵母雙雜交技術(shù)經(jīng)常出現(xiàn)較高的假陽性率和假陰性率[5]。熒光共振能量轉(zhuǎn)移能夠檢測活細(xì)胞中的PPI,但是,該方法需要靶蛋白高度表達(dá)[4]。雙分子熒光互補(bǔ)技術(shù)的檢測步驟相對簡單,報(bào)告蛋白熒光可以用熒光顯微鏡進(jìn)行檢測[6]。由于重組熒光蛋白自身帶有熒光,因此,雙分子熒光互補(bǔ)技術(shù)可以在活細(xì)胞中直接觀察到相互作用,從而避免了用外源性試劑處理細(xì)胞可能引起的一些問題[4]。這種直接觀察也有助于PPI 的亞細(xì)胞定位分析[7]。雙分子熒光互補(bǔ)技術(shù)可以分析活細(xì)胞中穩(wěn)定的PPI,也可以分析活細(xì)胞中短暫的或經(jīng)過特殊處理誘導(dǎo)的PPI[4]。例如,使用雙分子熒光互補(bǔ)技術(shù)證明了Phot1 在黑暗中以單體的形式存在,而在藍(lán)光處理后可以形成同型二聚體[8]。截至目前,已有超過10 種熒光蛋白被用于雙分子熒光互補(bǔ)技術(shù)[9]。然而,目標(biāo)蛋白的功能可能會(huì)受雙分子熒光互補(bǔ)片段的影響;此外,還應(yīng)控制融合蛋白的表達(dá),以排除因融合蛋白過多而導(dǎo)致的雙分子熒光互補(bǔ)復(fù)合物的非特異性形成;雙分子熒光互補(bǔ)技術(shù)的另一個(gè)局限性是發(fā)色團(tuán)成熟緩慢,這意味著該方法不適合檢測瞬態(tài)和動(dòng)態(tài)的PPI[4]。并且,雙分子熒光互補(bǔ)技術(shù)發(fā)色團(tuán)的成熟在體內(nèi)是不可逆的,這可能導(dǎo)致對相互作用蛋白定位的錯(cuò)誤判斷[6]。
2 新型PPI識(shí)別系統(tǒng)的設(shè)計(jì)
2.1 PUP-IT 鄰近標(biāo)記系統(tǒng)
PafA 介導(dǎo)Pup 與底物蛋白的賴氨酸結(jié)合[10]。在2018 年,利用細(xì)菌的Pup 蛋白結(jié)合系統(tǒng),將PafA基因與誘餌蛋白(Bait)進(jìn)行基因融合,它會(huì)將脫氨后的Pup(E)連接到附近的Bait 的賴氨酸上,同時(shí),也可以標(biāo)記與Bait 相互作用的獵物蛋白(Prey),從而可以研究PPI,該系統(tǒng)被稱為PUP-IT 鄰近標(biāo)記系統(tǒng)(圖1),在體外試驗(yàn)和細(xì)胞試驗(yàn)中均證實(shí)了PUP-IT對較弱的PPI 的識(shí)別作用。此外,因?yàn)镻up(E)停留在PafA 上,底物上的賴氨酸是否被修飾取決于PafA和Prey 之間的幾何形狀和方向,因此需要在PafA 和Bait 之間添加相對靈活且長的連接肽,以便允許結(jié)合的Prey 能夠以不同的幾何形狀呈現(xiàn),在該系統(tǒng)的所有設(shè)計(jì)中,設(shè)計(jì)者在PafA 和Bait 之間使用了15~20 個(gè)氨基酸的連接肽,同時(shí),設(shè)計(jì)者也建議在未來對分子較大的蛋白質(zhì)進(jìn)行研究時(shí),應(yīng)考慮使用較長的連接肽進(jìn)行有效標(biāo)記[11]。

圖1 PUP-IT 鄰近標(biāo)記系統(tǒng)
2.2 Mito-docking 系統(tǒng)
基于蛋白質(zhì)的重新定位和富集,2019 年選擇線粒體作為錨定平臺(tái),將A 錨定到線粒體外膜上作為Bait,從而捕獲并富集其與蛋白B 的相互作用,而不與蛋白A 相互作用的蛋白C 的定位不會(huì)受影響,這樣當(dāng)?shù)鞍譇 和蛋白B 標(biāo)記不同的熒光蛋白時(shí),通過使用熒光顯微鏡觀察蛋白B 在線粒體上的重新定位和富集,可以簡單有效地確定蛋白A 和B 之間的相互作用,此方法被命名為Mito-docking系統(tǒng)(圖2)[12]。
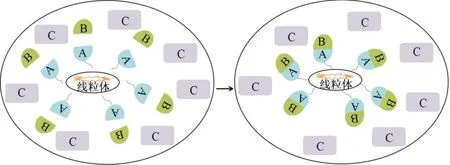
圖2 Mito-docking 系統(tǒng)
設(shè)計(jì)者選擇了外膜移位酶20 kDa 亞基(Translo?case of the outer membrane 20 kDa subunit,Tom20)的膜靶向序列的1~40 位氨基酸和干擾素β 啟動(dòng)子刺激蛋白1(Interferon beta promoter stimulator protein 1,IPS-1)的462~540 位氨基酸分別作為氨基端(Mi?toN)和羧基端(MitoC),將融合蛋白固定在線粒體,首先確定它們是否能將常用的熒光蛋白EGFP 和DsRed2 靶向線粒體,研究發(fā)現(xiàn)融合蛋白在線粒體上的定位是正確的,這表明MitoN 和MitoC 可以有效地將蛋白靶向到線粒體外膜;隨后,設(shè)計(jì)者應(yīng)用該方法檢測到G 蛋白亞基γ2(Gγ2)和亞基β1(Gβ1)之間的已知相互作用,確定了Mito-docking 系統(tǒng)可以在體內(nèi)有效檢測PPI[12]。運(yùn)輸受體與貨物蛋白之間的PPI在蛋白質(zhì)的核運(yùn)輸中發(fā)揮重要作用[13]。但由于這種PPI 是高度動(dòng)態(tài)的,因此通過傳統(tǒng)方法進(jìn)行檢測存在困難,設(shè)計(jì)者使用Mito-docking 系統(tǒng),首次使SV40 核定位信號(hào)和人類核輸入蛋白α 受體的PPI 可視化,有助于進(jìn)一步理解核轉(zhuǎn)運(yùn)機(jī)制[12]。
2.3 splitFAST 系統(tǒng)
光敏黃色蛋白是一種來自嗜鹽藻的藍(lán)光感光體,Y-FAST 是其工程化變體,Y-FAST 以可逆方式結(jié)合HBR 或HMBR 2 種熒光素,HBR 和HMBR 本身不發(fā)熒光,但是當(dāng)與Y-FAST 結(jié)合時(shí),它們會(huì)在藍(lán)光激發(fā)時(shí)發(fā)出黃光,Y-FAST 不僅能夠特異性地、瞬時(shí)地結(jié)合熒光素,而且結(jié)合還具有高度動(dòng)態(tài)性和完全可逆性,因此,可以簡單地通過添加或去除熒光素來快速打開和關(guān)閉熒光,能夠很好地控制整個(gè)系統(tǒng)[14]。此后,有研究報(bào)道了HBR-3,5DOM 這種新的熒光素可以與Y-FAST 形成緊密復(fù)合物,在綠光激發(fā)下能快速發(fā)出紅光,因此,除了允許對活細(xì)胞中Y-FAST 標(biāo)記的蛋白進(jìn)行多色成像外,因?yàn)閅-FAST標(biāo)記的可逆性,這些熒光素還可以實(shí)現(xiàn)動(dòng)態(tài)顏色切換[15]。有研究對Y-FAST 做了進(jìn)一步的改進(jìn),使其形成綠色和紅色熒光報(bào)告基因,能夠分別比綠色熒光蛋白EGFP 和紅色熒光蛋白mCherry 亮1.6 倍和2倍[16]。基于Y-FAST,在2019 年設(shè)計(jì)者將Y-FAST拆分為2 個(gè)片段,即NFAST(1~114 位氨基酸)和CFAST11(115~125 位氨基酸),這是一種可逆的拆分熒光報(bào)告基因,允許實(shí)時(shí)監(jiān)測蛋白質(zhì)組合的形成和解離,該系統(tǒng)被命名為splitFAST(圖3),通過連續(xù)除去CFAST11 羧基端的殘基得到CFAST10、CFAST9 和CFAST108,可以進(jìn)一步降低Y-FAST 2個(gè)拆分片段的表觀親和力[17]。
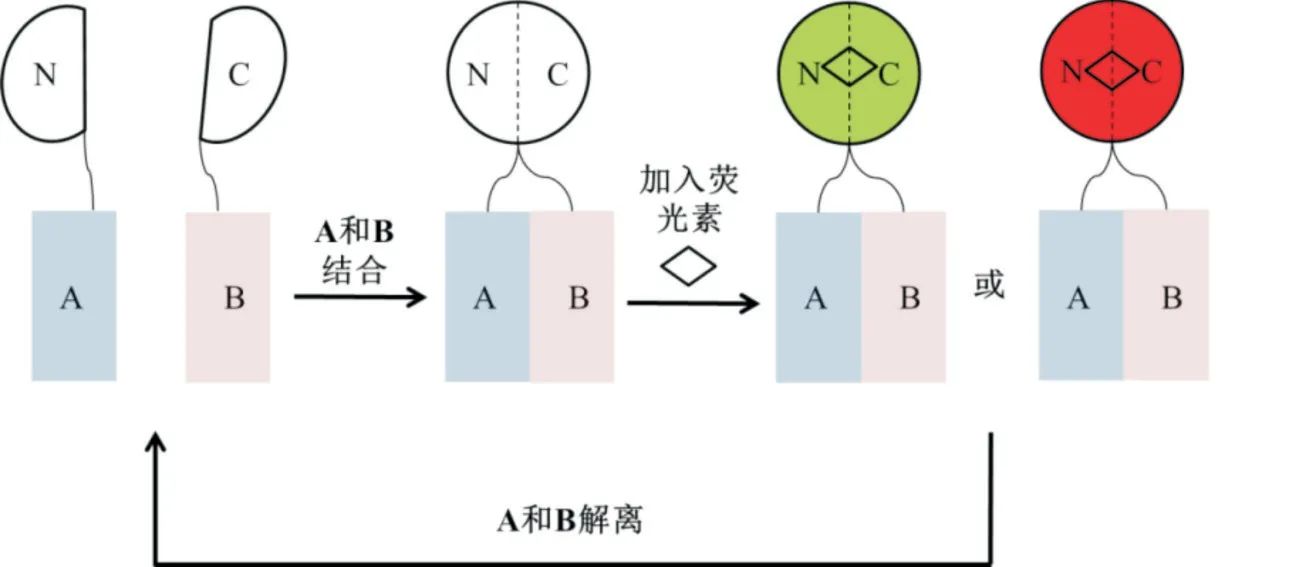
圖3 splitFAST 系統(tǒng)
雷帕霉素是一種天然產(chǎn)物,與FKBP 蛋白結(jié)合,然后,F(xiàn)KBP-雷帕霉素復(fù)合物與FRB 結(jié)合,形成三元 復(fù) 合 物[18]。設(shè) 計(jì) 者 將NFAST 和CFAST10 或CFAST11 分別與FKBP 和FRB 融合,依賴于雷帕霉素誘導(dǎo)的FRB-FKBP 互作,splitFAST 兩片段互補(bǔ)結(jié)合,引起了熒光的大幅增加,證明了splitFAST 系統(tǒng)具有檢測經(jīng)HMBR 或HBR-3,5DOM 預(yù)處理的哺乳動(dòng)物細(xì)胞中PPI 的能力;在胞質(zhì)中表達(dá)FKBPCFAST10 或FKBP-CFAST11 并在質(zhì)膜上表達(dá)FRBNFAST,雷帕霉素的添加導(dǎo)致HMBR 處理的細(xì)胞在質(zhì)膜處快速形成splitFAST 熒光,證明其能夠?qū)崟r(shí)識(shí)別 質(zhì)膜上 的PPI[17]。K-Ras 與Raf1 互作,并將Raf1從細(xì)胞質(zhì)招募到細(xì)胞膜[19]。設(shè)計(jì)者將NFAST 與KRas 融 合 表 達(dá),CFAST10 與mcherry 融 合 后,再 與Raf1 融合表達(dá),研究發(fā)現(xiàn)splitFAST 熒光與mCherry熒光共定位并集中在膜上,使用splitFAST 系統(tǒng)驗(yàn)證了K-Ras 與Raf1 的PPI[17]。MEK1 與ERK2 的PPI 發(fā)生 在 胞 質(zhì) 中[20]。將MEK1 與NFAST 融 合 表 達(dá),ERK2 與CFAST10 融合后,再與mCherry 融合表達(dá),研究能夠觀察到特異性的胞質(zhì)splitFAST 熒光,這說明使用splitFAST 系統(tǒng)也可以驗(yàn)證MEK1 與ERK2 的PPI[17]。核磷酸酶MKP1 能夠?qū)е翬RK2 去磷酸化并失活[21,22]。使用splitFAST 系統(tǒng)能夠檢測到ERK2 和MKP1 之間位于細(xì)胞核的PPI[17]。
2.4 SIMPL 系統(tǒng)
內(nèi)含肽在自然界中廣泛分布,其有著古老的起源,在細(xì)菌、古細(xì)菌和單細(xì)胞真核生物中均有分布,但在多細(xì)胞生物中是不存在的[23]。內(nèi)含肽被認(rèn)為是一種單一的轉(zhuǎn)換酶,其可以在沒有任何外部輔助因子或能量的幫助下從前體蛋白中去除自身[24]。內(nèi)含肽作為一種可動(dòng)的遺傳元件,通過蛋白質(zhì)剪接實(shí)現(xiàn)前體翻譯后自切除,產(chǎn)生活性蛋白[25]。將內(nèi)含肽拆分為2 個(gè)部分,并分別與2 種蛋白相連,在2 個(gè)拆分內(nèi)含肽的作用下,可以對蛋白質(zhì)進(jìn)行反式剪接[26]。GP41-1 拆分內(nèi)含肽是目前已知的反應(yīng)最快的蛋白質(zhì)反式剪接內(nèi)含肽[27]。在2020 年,設(shè)計(jì)者將Bait 在其羧基端與V5 標(biāo)簽和內(nèi)含肽氨基端片段(IN)融合,Prey 在其氨基端與1 個(gè)FLAG 標(biāo)簽和內(nèi)含肽羧基端片段(IC)融合,Bait 和Prey 在選定的哺乳動(dòng)物細(xì)胞中共表達(dá),以研究它們的體內(nèi)PPI,Bait 和Prey 的結(jié)合使IN 和IC 緊密相連,從而使它們能夠重新構(gòu)建功能齊全的內(nèi)含肽,然后催化自身的切除以及Bait 和Prey(包括V5 和FLAG 標(biāo)簽)的連接,可以通過常規(guī)蛋白質(zhì)印跡分析來解析所得的剪接蛋白,而V5 和FLAG 標(biāo)簽的存在允許使用常規(guī)生化技術(shù)可視化或純化蛋白(圖4),設(shè)計(jì)者在該系統(tǒng)中選用GP41-1 拆分內(nèi)含肽,對其進(jìn)行了重新改造,成功地降低了IN 和IC 2 個(gè)片段的內(nèi)在親和力,并具有良好的剪接性能[28]。
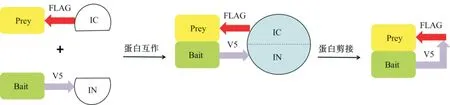
圖4 SIMPL 系統(tǒng)
使用SIMPL 系統(tǒng),在瞬時(shí)轉(zhuǎn)染體系中驗(yàn)證了雷帕霉素誘導(dǎo)的FKBP1A 和FRB 的PPI,且呈現(xiàn)雷帕霉素濃度依賴性,在穩(wěn)態(tài)細(xì)胞系中,經(jīng)過不同劑量的四環(huán)素孵育,誘導(dǎo)細(xì)胞具有不同表達(dá)水平的FRB 和FKBP1A,使用雷帕霉素處理后,在所有樣品中均觀察到FRB 和FKBP1A 的PPI 引起的剪接,這些結(jié)果證明了SIMPL 系統(tǒng)可以用于PPI 研究,并且可以在各種Bait和Prey 表達(dá)水平上研究PPI;為了進(jìn)一步擴(kuò)展系統(tǒng)的檢測能力,設(shè)計(jì)者設(shè)計(jì)了多種SIMPL 的形式,并驗(yàn)證了多種形式的SIMPL 系統(tǒng)均能有效檢測PPI[28]。EGFR 發(fā)生自磷酸化后,可以招募SHC1[29]。EGFR 突變體(L858R)活性增強(qiáng),而EGFR 突變體(D855A)是激酶死亡突變體[30,31]。在表皮生長因子刺激前,EGFR 以非活性單體或二聚體形式存在,在表皮生長因子刺激后,EGFR 活性二聚體或多聚體占主導(dǎo)地位[32]。SIMPL 系統(tǒng)能夠有效地檢測到EG?FR 和SHC1 的PPI,并且PPI取決于EGFR 的活性,因?yàn)镋GFR 突變體(L858R)增強(qiáng)了PPI 信號(hào),突變體(D855A)消除了PPI。然而,在沒有表皮生長因子處理的情況下,在瞬時(shí)過表達(dá)EGFR 和SHC1 的細(xì)胞中可以發(fā)生剪接,并且在表皮生長因子刺激后信號(hào)僅略有增強(qiáng),這可能是由于EGFR 的過表達(dá)誘導(dǎo)了非配體依賴性的自發(fā)激活,并隨后招募SHC1,剪接是不可逆的。因此,會(huì)導(dǎo)致剪接蛋白積累,這種積累的剪接蛋白可能掩蓋了表皮生長因子刺激誘導(dǎo)的信號(hào)。克服這個(gè)問題的關(guān)鍵是減少Bait 和Prey 表達(dá)的水平和持續(xù)時(shí)間,通過構(gòu)建穩(wěn)態(tài)細(xì)胞系可以有效地解決這一問題;SIMPL 系統(tǒng)也能夠檢測K-Ras 和Raf1 的PPI;SIMPL 系統(tǒng)能夠?qū)δ繕?biāo)蛋白激酶及底物的短暫和相對較弱的PPI 進(jìn)行識(shí)別,識(shí)別率高于
50%[28]。
3 新型PPI識(shí)別系統(tǒng)的特點(diǎn)
3.1 PUP-IT 鄰近標(biāo)記系統(tǒng)
PUP-IT 鄰近標(biāo)記系統(tǒng)不可避免地會(huì)產(chǎn)生不必要的自我修飾,這可能會(huì)使酶失活,消耗底物,并在質(zhì)譜試驗(yàn)中引入背景信號(hào),更精確的時(shí)間和局部控制可能會(huì)減少標(biāo)記背景;PafA 酶的連接不能保證被標(biāo)記的蛋白質(zhì)會(huì)直接與Bait 相互作用,任何在PafA標(biāo)記半徑內(nèi)的間接相關(guān)蛋白也可以被修飾[11]。
BioID 是對原核生物素連接酶BirA 進(jìn)行了突變(R118G)的酶,BioID 能夠催化生物素和ATP 生成反應(yīng)性生物素-AMP(bioAMP),bioAMP 被釋放并可以與相鄰的胺類(例如賴氨酸)發(fā)生共價(jià)反應(yīng)[33]。在H2O2存在的情況下,抗壞血酸過氧化物酶(Ascor?bate peroxidase,APEX)將生物素酚氧化為苯氧基,從而生物素化附近的蛋白質(zhì)[34]。在體外試驗(yàn)和細(xì)胞試驗(yàn)中,PUP-IT 鄰近標(biāo)記系統(tǒng)比BioID 更活躍,能夠提供更具特異性的標(biāo)記和富集[11]。PUP-IT 鄰近標(biāo)記系統(tǒng)不像APEX 那么活躍,因此可能不適合用于快速信號(hào)循環(huán)中的PPI 研究[11,35,36]。H2O2處理細(xì)胞可能會(huì)影響細(xì)胞的氧化狀態(tài)并導(dǎo)致細(xì)胞應(yīng)激,而PUP-IT 鄰近標(biāo)記系統(tǒng)不需要使用H2O2處理細(xì)胞,并且PUP-IT 鄰近標(biāo)記系統(tǒng)有潛力應(yīng)用于動(dòng)物模型,其他的標(biāo)記方法需要將化合物輸送到細(xì)胞中,這在活體動(dòng)物中很難實(shí)現(xiàn),而PUP-IT 鄰近標(biāo)記系統(tǒng)的所有成分都可以在細(xì)胞中表達(dá),因此,可以通過遺傳方法進(jìn)行修飾[11]。
3.2 Mito-docking 系統(tǒng)
設(shè)計(jì)者也同時(shí)使用了雙分子熒光互補(bǔ)技術(shù)和傳統(tǒng) 的共定位 方法對Gγ2和Gβ1之間 的PPI 進(jìn) 行了檢測,分析比較后發(fā)現(xiàn),雙分子熒光互補(bǔ)技術(shù)會(huì)產(chǎn)生假陽性結(jié)果,這可能是由拆分熒光蛋白片段的不可逆互補(bǔ)和拆分片段具有自發(fā)進(jìn)行重組的趨勢所導(dǎo)致的,傳統(tǒng)共定位方法檢測到mCherry-Gβ1與EGFPGγ2在細(xì)胞質(zhì)中的共定位,而在對照細(xì)胞中,EGFP 與mCherry-Gβ1共定位,mCherry 與EGFP-Gγ2共 定位,Mito-docking 系統(tǒng) 通 過MitoN 將EGFP 或EGFP-Gγ2錨定到線粒體外膜上,可特異性檢測到mCherry-Gβ1和MitoN-EGFP-Gγ2共表達(dá)的細(xì)胞在線粒體上的相互作用,與正常表達(dá)mCherry-Gβ1和MitoN-EGFP 或mCherry 和MitoN-EGFP-Gγ2的細(xì)胞線粒體上的信號(hào)明顯不同,這說明了Mito-docking 系統(tǒng)優(yōu)于雙分子熒光互補(bǔ)技術(shù),并且與傳統(tǒng)的共定位方法相比Mitodocking 系統(tǒng)具有更特異性的共定位效果[12]。
3.3 splitFAST 系統(tǒng)
用HMBR 或HBR-3,5DOM 預(yù)處理哺乳動(dòng)物細(xì)胞,設(shè)計(jì)者使用splitFAST 系統(tǒng)檢測細(xì)胞中PPI時(shí),發(fā)現(xiàn)加入雷帕霉素后的延時(shí)成像在幾分鐘內(nèi)顯示出熒光飽和,這說明splitFAST 系統(tǒng)可以實(shí)時(shí)監(jiān)測蛋白質(zhì)復(fù)合物的形成[17]。雷帕霉素能夠解離AP1510 誘導(dǎo)的FKBP 同源二聚體[37]。設(shè)計(jì)者將共表達(dá)FKBPNFAST 和FKBP-CFAST10 或FKBP-CFAST11 的 細(xì)胞與AP1510 一起孵育2 h 以形成FKBP 同源二聚體并用HMBR 處理,雷帕霉素的添加導(dǎo)致80%~90%的splitFAST 熒光損失,證明了splitFAST 組裝的可逆性,加入雷帕霉素后觀察到在幾分鐘內(nèi)熒光快速喪失,證明當(dāng)2 種蛋白質(zhì)解離時(shí)splitFAST 的快速分解[17]。在受到刺激后,ERK2 從細(xì)胞質(zhì)轉(zhuǎn)移到細(xì)胞核,在細(xì)胞核中誘導(dǎo)增殖和分化等生理過程,ERK2能夠通過核輸出機(jī)制返回到細(xì)胞質(zhì)[38,39]。在用表皮生長因子刺激細(xì)胞后,觀察到splitFAST 熒光減少和mCherry 熒光的核積累,這說明了mCherry-ERK2-CFAST10 從MEK1-NFAST 解離并轉(zhuǎn)移至細(xì)胞核,mCherry-ERK2-CFAST10 的核積累是短暫的,split?FAST 熒光和胞質(zhì)mCherry 熒光的同時(shí)增加,說明了mCherry-ERK2-CFAST10 返回細(xì)胞質(zhì)并與MEK1-NFAST 重新組裝,這些結(jié)果證明了splitFAST 系統(tǒng)能夠?qū)崟r(shí)觀察信號(hào)通路中的動(dòng)態(tài)PPI,splitFAST 系統(tǒng)對鈣調(diào)蛋白和M13 肽之間鈣離子依賴性的瞬時(shí)PPI的檢測,進(jìn)一步證明了其具有檢測快速、瞬時(shí)PPI 的能力,并且splitFAST 系統(tǒng)在細(xì)胞生物傳感器設(shè)計(jì)方面也具有巨大潛力[17]。
3.4 SIMPL 系統(tǒng)
SIMPL 系統(tǒng)具有檢測短暫和相對較弱的PPI 的能力;SIMPL 系統(tǒng)可以使用酶聯(lián)免疫吸附試驗(yàn)來對蛋白質(zhì)剪接進(jìn)行檢測,因此,該系統(tǒng)能夠?qū)崿F(xiàn)高通量的、可量化的PPI 檢測;SIMPL 系統(tǒng)還可以被用作一種基于活細(xì)胞的藥物篩選測定方法[28]。SIMPL 系統(tǒng)能夠識(shí)別線粒體中參與氧化磷酸化、轉(zhuǎn)運(yùn)、嵴形成等過程的蛋白之間的PPI,識(shí)別率為80%[28,40-42]。但是,由于需要Bait 和Prey 的異源表達(dá),因此,除非通過基因編輯將標(biāo)簽整合到目標(biāo)基因中,否則SIMPL系統(tǒng)無法測量內(nèi)源性PPI,另外,由于SIMPL 系統(tǒng)的作用具有不可逆性,因此該系統(tǒng)不能追蹤蛋白質(zhì)的解離[28]。
4 展望
傳統(tǒng)PPI 研究方法已經(jīng)得到較為廣泛的使用,但也存在其各自的問題,近年來開發(fā)出的新型PPI識(shí)別系統(tǒng)能夠較好地彌補(bǔ)傳統(tǒng)PPI 研究方法的局限性。總體而言,這幾種新型PPI 識(shí)別系統(tǒng)使用了鄰近標(biāo)記、蛋白質(zhì)共定位、拆分蛋白的設(shè)計(jì)思路。盡管仍然有很多問題需要繼續(xù)進(jìn)行改進(jìn),理解這幾種新型PPI 識(shí)別系統(tǒng)將對未來研究開發(fā)更高效的PPI 識(shí)別系統(tǒng)具有很大的指導(dǎo)價(jià)值。
猜你喜歡
工業(yè)設(shè)計(jì)(2022年8期)2022-09-09 07:43:20
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
軍民兩用技術(shù)與產(chǎn)品(2021年10期)2021-03-16 06:05:30
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
北京測繪(2020年12期)2020-12-29 01:33:58
裝備制造技術(shù)(2019年12期)2019-12-25 03:06:46
中國洗滌用品工業(yè)(2019年4期)2019-05-11 09:27:34
