美洲鰣內源性營養階段脂肪酸組成及利用
2022-08-11 12:18:12施永海徐嘉波嚴銀龍張之文劉永士于愛清鄧平平袁新程
海洋漁業 2022年3期
關鍵詞:營養
施永海,徐嘉波,嚴銀龍,張之文,劉永士,于愛清,鄧平平,蔣 飛,袁新程
(上海市水產研究所,上海市水產技術推廣站,上海 200433)
美洲鰣(Alosa sapidissima),又名美洲西鯡,屬于鯡形總目(Clupeomorpha),鯡形目(Clupeiformes),鯡科(Clupeidae),西鯡屬,為洄游性魚類,原來主要分布于北美洲大西洋西岸海洋和通海河流中[1-3],中國于1998年開始從美國引進美洲鰣受精卵進行人工繁養[4]。由于美洲鰣與中國鰣(Tenualosa reevesii)的外形相似、肉質鮮美,美洲鰣也深受國內市場的追捧,銷售價格較高[5-6]。近幾年,隨著美洲鰣全人工繁育技術的突破,國內養殖量也日趨增長,苗種年投放量大概在200多萬尾。雖然美洲鰣人工繁育技術日趨完善,但在美洲鰣苗種培育早期,特別是仔魚內源性營養階段成活率較低,這也成為美洲鰣規模化人工繁育的技術瓶頸。營養支持對魚類早期發育(特別是內源性營養階段和混合營養階段)至關重要,支撐魚類內源性營養階段個體生長發育的營養和能量來源主要是儲存在其卵黃囊中的脂類,而且脂類還參與調節生理活動和構建器官組織[7-8],所以,脂類含量和脂肪酸組分密切關系著魚類早期生命階段的生長、發育及存活[8-10],特別是DHA和EPA是海水魚類早期發育的必需脂肪酸,仔稚魚階段是腦神經和視神經迅速發育的時期,仔稚魚需要大量的DHA等重要營養物質,來滿足腦神經和視神經發育的需求[8,10]。了解魚類早期的營養利用有助于探究魚類在該發育階段的生命生理活動,進而從營養角度分析其生長和存活規律。因此,研究仔魚對脂類的利用和消耗有助于探究美洲鰣早期發育階段成活率低的原因。
迄今有關魚類早期生命階段對脂肪酸利用的研究大多集中于海水或者淡水魚類,如:菊黃東方鲀(Takifugu flavidus)[10]、日本鬼鲉(Inimicus japonicus)[7]、條石鯛(Oplegnathus fasciatus)[8]、黃顙 魚(Pelteobagrus fulvidraco)[9]和白鰱(Hypophthalmichthys molitrix)[11],而海淡水洄游性魚類發育早期對脂肪酸的利用相對比較特殊[12],美洲鰣作為典型的溯河繁育、降海育肥的洄游性魚類,其內源性營養階段對脂類的利用可能更為特殊。近年來,國內外有關美洲鰣的脂類營養狀況和需求的研究主要集中于開口攝食后的仔魚脂肪酸組成[13]、當年魚種的脂肪酸組成變化[4]、幼魚的脂肪需求[14-16]、成魚的肌肉營養成分分析[17-20]等。有關美洲鰣發育早期內源性營養階段脂類代謝的研究目前尚未見公開報道。本文檢測了美洲鰣胚胎孵化期間和仔魚開口前的脂肪酸組成及含量,分析了美洲鰣胚胎出膜前后和仔魚開口前對脂肪酸的利用程度,旨在揭示美洲鰣發育早期對脂肪酸的利用規律,以期為提高美洲鰣發育早期仔魚成活率相關研究提供參考。
1 材料與方法
1.1 樣品采集與處理
實驗用美洲鰣的受精卵和仔魚來自上海市水產研究所奉賢科研基地,于2018年6月底到7月初全人工繁育獲得,親本培育和人工繁育方法參照張根玉等[1]和施永海等[5-6]。選取人工促產、自然交配、產卵受精的魚卵作為實驗材料,魚卵從集卵箱收集、清洗后轉移到孵化池,孵化池為錐形底,水體為1.6 m3,每個孵化池內設置2個氣石,孵化池上方約2 m處覆蓋雙層遮蔭率為90%的遮蔭膜,連續曝氣,靜水孵化,孵化密度為8~10萬粒·池-1,每天換水1次,每次換水總水體的2/3,溫度控制為(18.98±0.32)℃,仔魚孵化出膜后,還繼續在孵化池內暫養至魚苗開口;采用解剖鏡吸管吸取的方法[12]分別獲取胚胎(原腸期)、0日齡和4日齡仔魚(攝食前),隨機取樣,用濾紙吸干表面水分,每個發育階段算一組,每組樣品分3個平行,每個平行按胚胎(15~17 g)和仔魚(2~3 g)取樣,再測算每組樣品單位質量的個體數。樣品用離心管密封保存,先在-20℃的冰箱冷凍24 h,而后轉移到-76℃的冰箱保存。
1.2 樣品測定方法
用SIM冷凍干燥機(型號:FD5-3,產地:美國)采用真空凍干法測定水分,用真空干燥箱(型號:DZF-6020,產地:中國上海)采用氯仿甲醇法提取粗脂肪,用氣相色譜儀(型號:Agilent 6890,產地:美國)測定及色譜峰峰面積歸一法計算出脂肪酸相對含量(GB/T22223-2008)[4]。
1.3 數據處理和統計
所有數據用mean±SD表示,采用Excel 2003和SPSS19.0處理數據。采用oneway ANOVA對數據進行方差分析,如數據是百分數,先用反正弦函數轉換后再方差分析,用Duncan法多重比較,以P<0.05表示有顯著差異[4]。
1.4 計算公式[12]
單個個體的脂肪酸絕對含量(Mn,μg):
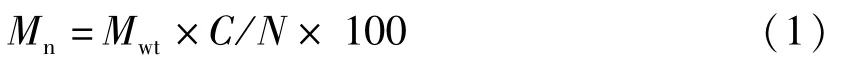
胚胎出膜階段脂肪酸減少量(Mg,μg):

胚胎出膜階段脂肪酸減少率(Rg,%):
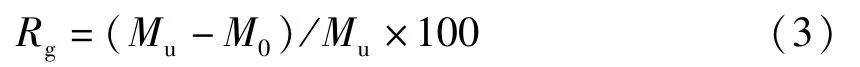
仔魚開口攝食前脂肪酸利用量(Mf,μg):

仔魚開口攝食前脂肪酸利用率(Rf,%):
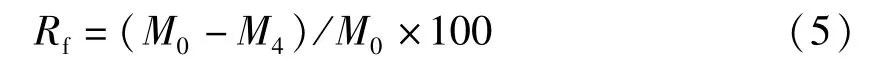
式中,Mwt是單位鮮重的粗脂肪含量(%),C是各脂肪酸的相對含量(%),N是各發育時期每克鮮重的生物體個數,Mu、M0和M4分別是胚胎、0日齡仔魚、4日齡仔魚個體的脂肪酸絕對含量(μg)。
2 結果與分析
2.1 內源性營養階段的水分和粗脂肪含量變化
美洲鰣內源性營養階段水分相對含量隨著個體的發育而顯著降低(P<0.05),每個發育階段的水分含量均顯著高于下個階段(P<0.05)(表1)。美洲鰣內源性營養階段干物質的粗脂肪相對含量隨個體發育微弱升高,各階段之間無顯著性差異(P>0.05)(表1);鮮質量的粗脂肪相對含量從胚胎的0.58%顯著上升至4日齡仔魚的2.25%(P<0.05)(表1);然而,由于個體發育消耗了體內大量的營養物質,單個個體質量迅速下降,導致單個個體的粗脂肪絕對含量隨個體發育呈現顯著下降趨勢(P<0.05),從胚胎的60.04μg顯著下降到4日齡仔魚的41.91μg(P<0.05)(表1)。
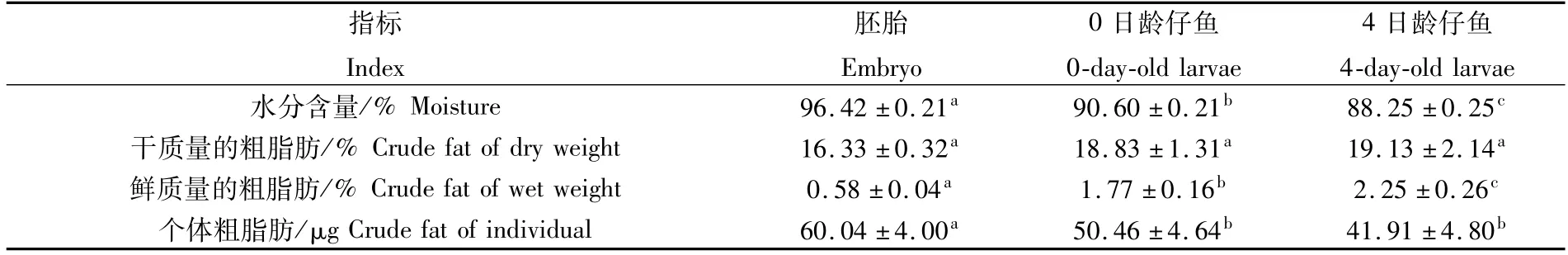
表1 美洲鰣內源性營養階段的水分和粗脂肪含量(n=3)Tab.1 Moisture and crude fat content of Alosa sapidissima during endogenous nutrient stage(n=3)
2.2 內源性營養階段脂肪酸的組成變化
2.2.1 單個脂肪酸相對含量的變化
美洲鰣胚胎、0日齡和4日齡仔魚檢測到的22種脂肪酸相對含量見表2,內源性營養階段各時期相對含量排前4的脂肪酸均分別是C16∶0、C18∶1n9c、C22∶6n3(二十二碳六烯酸,docosahexaenoic acid,DHA)和C18∶2n6c(表2)。
C16∶0相對含量較高(21.65%~23.06%),0日齡仔魚的C16∶0相對含量顯著高于胚胎和4日齡仔魚(P<0.05),且胚胎和4日齡仔魚之間無顯著性差異(P>0.05);C18∶1n9c的相對含量豐富(19.02%~20.93%),其相對含量隨著個體的發育而顯著降低(P<0.05);C18∶2n6c相對含量相對較穩定(12.33%~14.43%),其相對含量隨著個體的發育而顯著降低(P<0.05),4日齡仔魚的C18∶2n6c相對含量顯著低于胚胎和0日齡仔魚(P<0.05),而胚胎和0日齡仔魚之間的差異不顯著(P>0.05)(表2)。
美洲鰣胚胎、0日齡和4日齡仔魚的C22∶6n3(DHA)均較豐富(15.02%~18.20%),其相對含量隨著個體的發育而顯著升高(P<0.05),4日齡仔魚的C22∶6n3(DHA)相對含量顯著高于胚胎和0日齡仔魚(P<0.05),而胚胎和0日齡仔魚之間無顯著性差異(P>0.05)(表2)。美洲鰣內源性營養階段的C20∶5n3(二十碳五烯酸,eicosapentaenoic acid,EPA)雖然相對含量不高(1.05%~1.49%),但其相對含量也隨個體發育而顯著上升(P<0.05)(表2)。DHA和EPA含量均隨個體發育而上升的相似變化規律也使DHA+EPA隨個體發育呈現顯著的上升趨勢(P<0.05),4日齡仔魚的DHA+EPA(19.69%)顯著高于胚胎和0日齡仔魚(P<0.05),而胚胎和0日齡仔魚之間(分別是16.07%和16.63%)的差異不顯著(P>0.05)(表2)。
2.2.2 SFA、MUFA和PUFA相對含量的變化
美洲鰣胚胎的SFA相對含量(32.39%)顯著低于4日齡仔魚的(33.53%)(P<0.05),而0日齡仔魚SFA相對含量(33.39%)分別與胚胎以及4日齡仔魚的均無顯著變化(P>0.05)(表2)。MUFA 的相對含量相對較低(20.92%~22.82%),且隨個體發育而顯著下降(P<0.05)(表2)。美洲鰣內源性營養階段的PUFA含量豐富且穩定(44.76%~45.55%)(表2),胚胎、0日齡和4日齡仔魚之間的PUFA含量無顯著變化(P>0.05)(表2)。
2.2.3 ∑SFA/∑UFA和∑n3PUFA/∑n6PUFA比率的變化
美洲鰣4日齡仔魚的∑SFA/∑UFA比例(0.50)顯著高于胚胎的(0.48)(P<0.05)(表2)。美洲鰣內源性營養階段的∑n3PUFA/∑n6PUFA比例隨生長發育而明顯上升(P<0.05),其數值從0.69上升到0.94(表2),胚胎和0日齡仔魚的∑n3PUFA/∑n6PUFA比例顯著低于4日齡仔魚的(P<0.05),而胚胎和0日齡仔魚之間的差異不顯著(P>0.05)(表2)。

表2 美洲鰣內源性營養階段的脂肪酸組成及相對含量(n=3,%)Tab.2 Fatty acid composition and content of Alosa sapidissima during endogenous nutrient stage(n=3,%)
2.3 內源性營養階段單個個體的主要脂肪酸的絕對含量變化及利用
美洲鰣內源性營養階段單個個體中各脂肪酸的絕對含量見表3。由表3中各發育階段的脂肪酸絕對含量,依據公式可分別獲得主要脂肪酸在胚胎孵化出膜階段的減少量和減少率,以及在仔魚內源營養階段的利用量和利用率(表4)。
美洲鰣內源性營養階段的單個個體的脂肪酸絕對含量前4的分別為C16∶0、C18∶1n9c、C22∶6n3(DHA)和C18∶2n6c(表3)。隨著美洲鰣的個體發育以及卵膜的脫落,除了C24∶1n9絕對含量先略微下降(P>0.05)再顯著上升(P<0.05),C15∶0的絕對含量呈現顯著的先降后升趨勢(P<0.05),C17∶1絕對含量先略微上升(P>0.05)再顯著下降(P<0.05),C17∶0、C20∶0、C20∶4n6、C20∶5n3(EPA)及C22∶6n3(DHA)的絕對含量無顯著變化外(P>0.05),其余的14個脂肪酸絕對含量均呈現顯著的下降趨勢(P<0.05)(表3);多數脂肪酸下降的疊加效應造成了單個個體的SFA、MUFA和PUFA絕對含量隨個體發育呈現顯著的下降趨勢(P<0.05)(表3)。另外,EPA+DHA和∑n3PUFA絕對含量隨個體發育均無顯著性變化(P>0.05),而∑n6PUFA絕對含量呈現顯著的下降趨勢(P<0.05)(表3)。
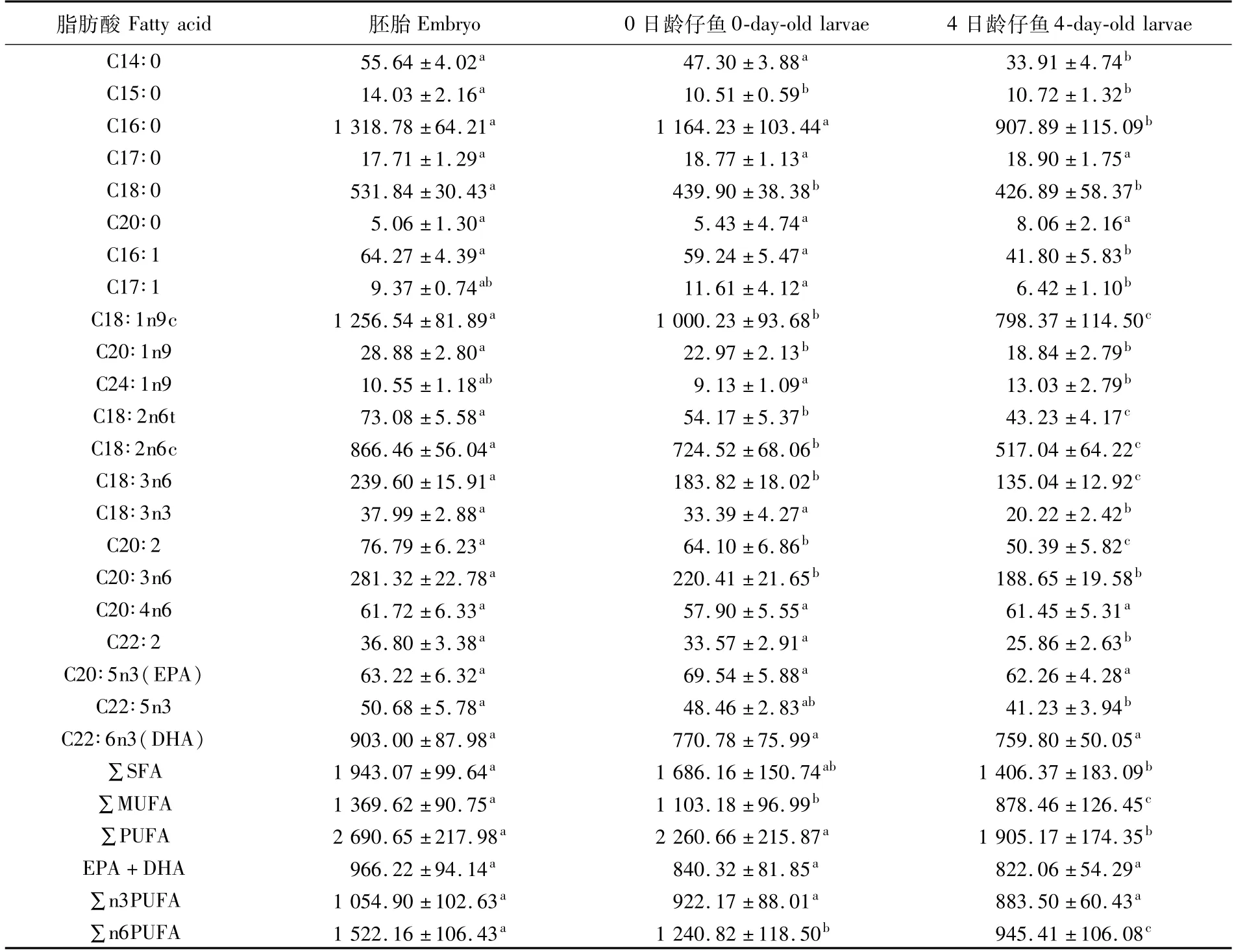
表3 美洲鰣內源性營養階段單個個體的脂肪酸絕對含量(n=3,μg)Tab.3 Fatty acid absolue content of individual Alosa sapidissima during endogenous nutrient stage(n=3,μg)
美洲鰣胚胎孵化脫膜,由于個體發育、出膜以及卵膜的脫落等因素造成主要脂肪酸實際減少量前4的分別是C18∶1n9c(256.31μg)、C16∶0(154.55μg)、C18∶2n6c(141.93μg)和DHA(132.23μg)(表4)。比較單個個體中SFA、MUFA和PUFA 的實際減少量,PUFA 為最高(429.99μg);從各主要脂肪酸的實際減少率來看,除了C20∶5n3(EPA),其他的主要脂肪酸減少率相對較均衡(11.72%~23.28%)(表4);SFA、MUFA和PUFA減少率也較均衡,數值分別為13.22%、19.45%和15.98%。綜合減少量和減少率,胚胎孵化破膜以及卵膜的脫落過程中,C18∶1n9c的減少較多(分別為256.31μg和20.40%),更值得注意的是,C20∶5n3(EPA)減少量和減少率分別為-6.32μg和-10.00%(表4)。
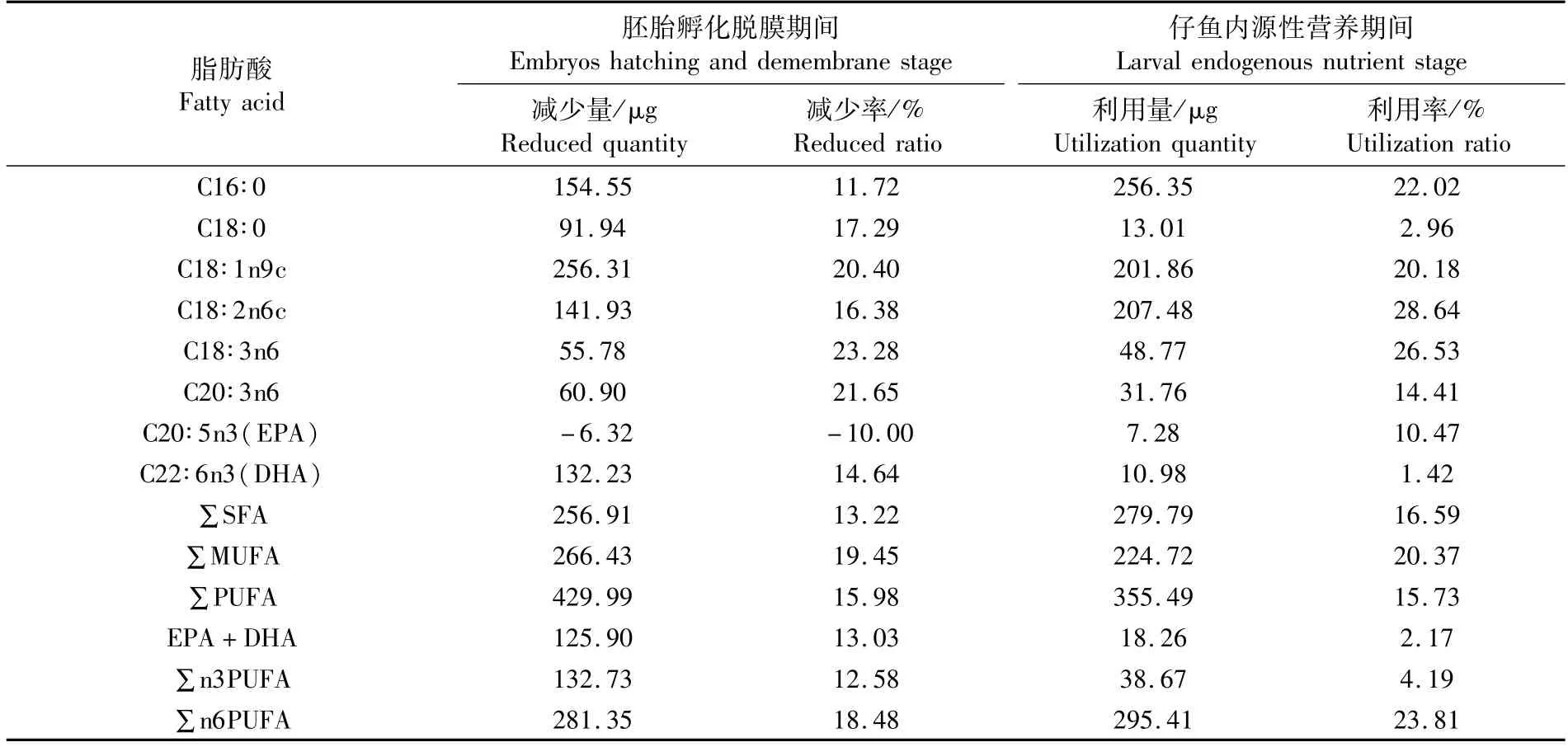
表4 美洲鰣內源性營養階段單個個體的主要脂肪酸的利用程度Tab.4 Utilization degree of main fatty acids of individual Alosa sapidissima during endogenous nutrient stage
美洲鰣仔魚開口攝食前,單個個體的各主要脂肪酸的利用量前3的分別是C16∶0(256.35 μg)、C18∶2n6c(207.48μg)和C18∶1n9c(201.86 μg),利用量最少的兩個脂肪酸是EPA和DHA(分別是7.28μg和10.98μg)(表4);比較單個個體SFA、MUFA和PUFA的實際利用量,PUFA為最高(355.49μg)。單個個體各主要脂肪酸的利用率前4的分別是C18∶2n6c(28.64%)、C18∶3n6(26.53%)、C16∶0(22.02%)和C18∶1n9c(20.18%),利用率最少的3個脂肪酸是DHA、C18∶0和EPA(分別是1.42%、2.96%和10.47%)(表4)。SFA、MUFA和PUFA利用率較均衡,數值分別為16.59%、20.37%和15.73%(表4)。綜合利用量和利用率,仔魚開口攝食前,利用較多的單個脂肪酸是C16∶0、C18∶2n6c和C18∶1n9c;值得注意是,美洲鰣開口前仔魚對∑n6PUFA的占用較多(295.41μg和23.81%),對∑n3PUFA的占用較少(38.67μg和4.19%),尤其對DHA+EPA的占用更少(18.26μg和2.17%)(表4)。
3 討論
3.1 美洲鰣內源性營養階段的粗脂肪含量及脂肪酸組分的特征
美洲鰣受精卵和開口前仔魚的干物質粗脂肪相對含量(16.33%~19.13%)與其他一些海水魚類早期發育階段干物質粗脂肪含量相似:如,條石鯛(9.27%~23.77%)[8]、日本鬼鲉(13.85%~11.66%)[7]和菊黃東方鲀(21.27%~31.70%)[10];美洲鰣內源性營養階段的干物質粗脂肪含量遠低于同是洄游性魚類的長江刀鱭(Coilia nasus)(53.10%~60.97%)[12],但由于美洲鰣內源性營養階段個體較大:受精卵的直徑[(4.03±0.25)mm][2]遠大于長江刀鱭的[(0.91±0.02)mm][21],造成美洲鰣內源性營養階段單個個體的粗脂肪量(41.91 μg~60.04 μg)高于長江刀鱭(16.27μg~43.62μg)。另外,相對于長江刀鱭單個個體在開口攝食前消耗掉62.70%的粗脂肪,美洲鰣單個個體開口前(4日齡仔魚)只消耗掉30.20%的粗脂肪,這說明美洲鰣仔魚開口攝食時,其體內還有較多脂類(41.91μg,占原有的69.80%)被留存,這從理論上解釋了美洲鰣仔魚開口攝食后混合營養階段較長的原因。
魚類為變溫動物,其脂肪中含有能降低脂肪酸熔點的n3PUFA,可維系細胞膜的滲透性和流動性[22]。海水魚類因需要調節滲透壓,其脂肪中含有較高n3PUFA以更好地維系良好的細胞膜通透 性[10,23]。相對于淡水魚類,海水魚類的∑n3PUFA/∑n6PUFA較高,在內源性階段,其比值一般高于2,如:日本鬼鲉(2.74~3.98)[7]、條石鯛(6.18~7.62)[8]和菊黃東方鲀(5.26~6.55)[10]。而美洲鰣發育早期內源性階段的∑n3PUFA/∑n6PUFA較低(0.69~0.94),其特征更接近淡水魚類,如:黃顙魚的受精卵(1.22~1.35)和開口前仔魚(1.19~2.26)[9]、鰱(Hypophthalmichthys molitrix)受精卵及仔魚(0.65~2.68)[11]、刺鲃(Spinibarbus caldwelli)魚卵(0.45)[24]、鯉(Cyprinus carpio)魚 卵(0.43)[25]和鯽(Carassius auratus)魚卵(0.88)[25],美洲鰣內源性營養階段較低的∑n3PUFA/∑n6PUFA比值也符合其生態洄游的習性,美洲鰣親魚從海里上溯到江河淡水環境中繁育,其早期發育階段均生活在淡水環境中,一般到當年的秋冬季,幼魚隨江河流水進入海洋越冬[4]。但美洲鰣幼魚進入海洋越冬前,為了適應外界鹽度和低水溫,需要更多的n3PUFA來維系細胞膜良好的滲透性和流動性。因此,美洲鰣仔稚幼魚發育期間有必要增加投喂富含n3PUFA的飼料,讓魚體內提前積累n3PUFA,提高美洲鰣對外界鹽度和低水溫的耐受力。
3.2 美洲鰣各生長發育階段的脂肪酸組分比較
美洲鰣魚卵[26]、胚胎、開口前仔魚、當年魚種魚體[4]、江蘇養成的成魚肌肉[20]之間的大多數脂肪酸組成及含量相近,而與攝食飼料的仔魚、攝食鹵蟲的仔魚[13]以及廣東養殖的成魚肌肉[18]差異較大:美洲鰣魚卵、胚胎、開口前仔魚、當年魚種魚體、江蘇養成的成魚肌肉之間的脂肪酸含量較多的均是C18∶1n9c、C16∶0、C18∶2n6c和C18∶0[4,20,26]。仔魚分別攝食鹵蟲和配合飼料后,其脂肪酸組分差異較大,如:C18∶1n9c(分別為32.54%和19.66%)、C16∶0(分別為10.41%和24.52%)和DHA(分別為0.26%10.71%)[13],美洲鰣當年魚種魚體和江蘇養殖的成魚肌肉的∑ n3PUFA、∑ n6PUFA、∑ SFA/∑ UFA、∑n3PUFA/∑n6PUFA(分別是3.65%~6.99%和3.65%、21.22%~27.12%和18.04%、0.40~0.58和0.45、0.15~0.29和0.20)[4,20]與廣東養殖成魚肌肉[18](分別是0.93%、44.06%、0.85和0.02)相差較大,同時廣東養殖成魚肌肉脂肪酸主要是C16∶0和C18∶2n6c,兩者加起來的百分含量達到67.66%,其C18∶0和C18∶1n9c的百分含量非常少,分別是0.79%和0.95%[18],造成這些現象的原因可能與飼料的脂肪酸組成有關。
魚卵脂類不僅是內源性營養階段的主要能量來源,還參與構建機體的組織和調節生理活動,因此,脂類及脂肪酸的含量與內源性營養階段的發育及存活有著密切的關系[10]。同時,美洲鰣(特別是親本)肌肉的營養蓄存與其所產魚卵的營養水平存在密切關系[10]。美洲鰣內源性營養階段(魚卵[26]、胚胎和開口攝食前仔魚)的∑n3PUFA含量(13.16%~21.16%),特別是EPA+DHA的含量(12.02%~19.69%)遠遠高于江蘇[20]和廣東[18]養殖成魚肌肉的含量(分別為3.65%和0.93%、0.69%和0.65%),同時美洲鰣魚卵[26]、胚胎和開口前仔魚的∑n3PUFA/∑n6PUFA雖然已經很低(0.51~0.94),但還是明顯高于其養殖成魚肌肉的(0.02~0.20)[18,20](表5)。因此,美洲鰣人工繁殖前,要挑選營養狀況優良的成魚作為后備親本,同時在親本培育期間,增加喂食富含n3PUFA(如DHA)的飼料,以加強美洲鰣親本營養積累。
3.3 美洲鰣胚胎出膜階段脂肪酸變化的特點
魚類的胚胎發育和出膜期間,脂類下降包括兩個方面:一方面是胚胎發育及出膜等生理和生命活動需要消耗大量體內能量,脂類作為魚卵中能量儲存的主要營養物質也隨之消耗;另一方面是卵膜的脫落造成結合在卵膜的脂類也跟隨丟失[11-12]。本研究中,美洲鰣干物質的粗脂肪含量在胚胎和0日齡仔魚之間的差異不顯著,而個體的粗脂肪含量顯著下降。從單個脂肪酸來看,主要脂肪酸中實際減少量前4的分別是C18∶1n9c、C16∶0、C18∶2n6c和DHA。綜合減少量和減少率,胚胎孵化破膜過程中,C18∶1n9c的減少較多(分別為256.31μg和20.40%),DHA也有較大程度的減少(分別為132.23 μg和14.64%);更值得注意的是,C20∶5n3(EPA)有所增加(減少量和減少率分別為-6.32μg和-10.00%)。因此,可以推斷,美洲鰣胚胎出膜階段,EPA被先行留存,而DHA被大量利用。雖然有研究表明有些魚類品種的發育初期有先期留存n3PUFA的征象,如:大黃魚(Pseudosciaena crocea)[23]、菊黃東方鲀[10]和長江刀鱭[12]等;但美洲鰣胚胎發育期間出現EPA暫存、DHA被大量利用的現象還比較少見,這也說明DHA在美洲鰣胚胎發育中的重要性。另外,EPA有所增加的原因,除了由于脂肪消耗對EPA的蓄留外,也可能是美洲鰣胚胎具有把短鏈的18C-PUFA轉變成長鏈的20C或22C-HUFA的功能[9,11]。
3.4 美洲鰣仔魚開口攝食前階段對脂肪酸利用的特點
研究證實,海水魚類自身一般不能合成n3PUFA,n3PUFA是需要通過食物鏈來攝食獲取的,n3PUFA 被認為是海水魚類的必需脂肪酸[10],如在魚類發育早期就必須要有足夠量的DHA等脂肪酸來滿足神經系統快速發育的需求[10]。本研究中,美洲鰣仔魚開口攝食前,對脂肪酸的消耗是有先后順序的:n6PUFA>MUFA>SFA>n3PUFA,特別是n6PUFA中的C18∶2n6c和C18∶3n6、SFA中的C16∶0、MUFA中的C18∶1n9c優先被仔魚利用,證明n6PUFA對美洲鰣開口前仔魚提供能量的重要性,而n3PUFA利用量和利用率極低(38.67μg和4.19%),尤其是EPA+DHA(18.26μg和2.17%),造成這種現象的原因可能有兩個方面:一方面是美洲鰣早期仔魚對DHA和EPA的暫存,DHA和EPA未被動用可能因為它是細胞膜的必需成分,對維系細胞膜結構和功能的完整性極為重要[9];另一方面是美洲鰣仔魚可能啟動了生物轉化功能(即把短C鏈的脂肪酸轉化合成為長C鏈的PUFA),仔魚把18C-PUFA(如:C18∶2n6c、C18∶3n6)轉變成20C或22C-HUFA[9],美洲鰣開口前仔魚 對C18∶2n6c、C18∶3n6的利用量和利用率數據上也有體現:C18∶2n6c(207.48μg和28.64%)和C18∶3n6(48.77μg和26.53%)。美洲鰣仔魚的生物轉化能力在其他淡水和河口性魚類上也已經有發現,如黃顙魚[9]和鰱(Hypophthalmichthys molitrix)[11],但海洋魚類是不具備的[11]。這也說明美洲鰣早期仔魚具有對脂肪酸的生物轉化能力,呈現了淡水魚類的特點。
3.5 美洲鰣內源性營養階段脂肪酸利用特點與仔魚存活
DHA與EPA是公認的動物生長發育的必要脂肪酸[27],魚類內源性營養階段是魚類神經系統發育的快速期,此階段就需要消耗大量的DHA來維系和支持其需求[9]。本研究中,美洲鰣胚胎發育期間對DHA的大量消耗,可能會造成初孵仔魚體內DHA的匱乏,較低水平的DHA可導致剛出膜的仔魚對環境響應水平的下降,以至于仔魚的死亡[28],這或許是造成美洲鰣胚胎出膜后到仔魚開口前的階段成活率較低的一個原因。因此,在美洲鰣親本培育和促產期間,要投喂富含DHA的飼料,以及時將DHA補充到親本體內及轉化到卵巢及魚卵中。
4 小結
美洲鰣內源性營養階段,單個個體的粗脂肪含量較高,仔魚開口前只消耗掉其中的30.20%。各時期的相對含量排前4的脂肪酸均分別是C16∶0、C18∶1n9c、C22∶6n3(DHA)和C18∶2n6c,其∑n3PUFA/∑n6PUFA較低。胚胎出膜階段,EPA被先期暫存;開口攝食前,仔魚對脂肪酸利用的先后順序為:n6PUFA >MUFA >SFA >n3PUFA,n3PUFA被優先保存,尤其是EPA+DHA。因此,建議美洲鰣親本培育和促產期間,增加投喂富含∑n3PUFA(特別是DHA)的飼料,以增強親本營養積累和及時補充∑n3PUFA(特別是DHA)到親本體內,增加魚卵營養儲備,進而提高美洲鰣開口前仔魚的成活率。
猜你喜歡
中老年保健(2022年5期)2022-11-25 14:16:14
今日農業(2021年11期)2021-08-13 08:53:34
今日農業(2021年4期)2021-06-09 06:59:58
今日農業(2020年24期)2020-03-17 08:58:18
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
廣東第二課堂·小學(2017年2期)2017-02-20 15:10:08
廣東第二課堂·小學(2017年1期)2017-02-20 14:47:45
少兒科學周刊·兒童版(2015年12期)2016-05-16 03:38:08
小學閱讀指南·低年級版(2016年8期)2016-05-14 13:24:45
