烤煙上部煙葉成熟生理特性研究進展
2022-08-22 08:19:46賈中林尹啟生戴華鑫梁太波張艷玲張仕祥
煙草科技 2022年7期
賈中林,尹啟生,戴華鑫,梁太波,張艷玲,劉 陽,張仕祥
中國煙草總公司鄭州煙草研究院,鄭州高新技術產業開發區楓楊街2號 450001
煙葉成熟度是決定烤煙品質的核心要素,田間煙葉成熟采收是優質煙葉生產的重要環節,直接影響烤后煙葉品質。左天覺[1]、宮長榮[2]認為田間煙葉成熟采收對煙葉品質的貢獻占整個烤煙生產環節的1/3。烤煙上部煙葉包括上二棚和頂葉在內的5~6片葉,占單株總產量的30%~40%,優質的上部煙葉對卷煙香味和風格有很大影響,是生產高檔卷煙的重要原料,在煙葉生產中占有重要地位。長期以來,我國許多煙區上部煙葉存在成熟度不夠、葉片僵硬、化學成分不協調、刺激性大、雜氣重以及工業可用性差等問題,導致多數上部煙葉不能直接用于一、二類卷煙配方中[3]。自2009年朱尊權院士提出烤煙上部4~6片煙葉一次性成熟采收技術后,許多產區相繼開展了提高上部煙葉可用性的技術研究與應用,并取得了顯著效果,部分上部煙葉已應用到高端卷煙品牌配方中[4-5]。然而,目前關于烤煙上部煙葉成熟進程中的物質轉化規律和相關作用機理等仍不明確,這在一定程度上限制了相關技術的應用。為此,從煙葉物質代謝、生物酶活性、基因表達和蛋白功能等方面綜述了近年來烤煙上部煙葉成熟生理特性的研究進展,旨在為煙葉成熟度調控和優質上部煙葉開發提供參考。
1 上部煙葉成熟過程中物質代謝規律
1.1 碳水化合物
1.1.1 碳水化合物種類及測定方法
煙葉碳代謝產物主要是碳水化合物(糖類),按結構組成分為單糖、低聚糖和多糖。20世紀60年代起,國外開始關注煙葉中的碳水化合物,主要采用酸水解法測定煙葉淀粉含量,Somogyi法測定煙葉中可溶性糖含量,離子交換色譜法來分離煙葉中的糖組分[6-7]。20 世紀80 年代起,國內也開始相關研究,起初采用二硝基水楊酸比色法測定還原糖和總糖含量,酸水解-二硝基水楊酸比色法或酸水解-碘比色法測定淀粉含量[8-9]。隨著分析技術的不斷發展,出現了光譜法、液相色譜法、氣相色譜法、離子色譜法和電泳法等更加快速精準的檢測方法[10-11],目前行業內對煙葉中總糖和淀粉的測定一般采用連續流動分析法,測定糖組分一般采用高效液相色譜法(表1)。

表1 煙葉中碳水化合物的檢測方法Tab.1 Detection methods for carbohydrates in tobacco leaves
1.1.2 碳水化合物與煙葉成熟度的關系
煙葉成熟過程中碳代謝產物的研究主要集中于總糖、還原糖和淀粉(表2)。在煙葉成熟過程中,葉片光合生理功能開始衰退,碳水化合物合成減弱而降解增強,物質含量逐漸降低[12]。國外研究表明,隨煙葉成熟度增加,淀粉、總糖和可溶性糖含量大致呈拋物線形變化,國內研究也發現同樣的規律,并認為煙葉淀粉含量的最大值出現在煙葉工藝成熟時期,之后進入過熟階段,煙葉淀粉含量逐漸降低[13-14]。煙葉成熟過程中物質含量的變化可為生產上煙葉成熟度的判斷提供參考,如美國通過提前1周測定煙葉中化學成分含量來判斷煙葉的成熟度,以此來確定煙葉采收期[15]。20世紀80年代起,國內學者也提出將淀粉含量作為煙葉成熟度的判斷指標,淀粉含量達到最高值的時間即為煙葉工藝成熟期[18]。煙葉碳代謝產物在成熟過程中的變化受品種、地域環境等因素的影響。不同品種烤煙成熟過程中,煙葉淀粉和還原糖含量雖然整體上表現為先升高后降低的變化趨勢,但達到峰值的時間存在差異[22]。同一烤煙品種在不同地域環境下,煙葉糖含量在成熟過程中呈現的變化規律也略有差異[23]。目前對煙葉成熟過程中糖含量的研究主要集中在總糖和還原糖總量方面,而對糖組分如葡萄糖、果糖、蔗糖和麥芽糖等的關注相對較少。

表2 國內外煙葉成熟過程中碳代謝產物的變化主要研究結果Tab.2 Domestic and foreign researches on variations of carbon metabolites during tobacco leaf ripening
1.2 含氮化合物
1.2.1 含氮化合物種類及測定方法
煙葉中氮代謝產物主要是含氮化合物,包括蛋白質、游離氨基酸和生物堿等,含氮化合物對煙葉的感官質量和吸食者健康有重要影響。國外對煙葉含氮化合物的研究始于20世紀60年代,最初主要采用凱氏定氮法測定蛋白質,該方法具有操作簡單、測量準確等優勢,也是較長一段時期國內測定蛋白質的常用方法。目前,行業內總氮的測定主要采用連續流動分析法[24]。1958 年,Moore 等[25]發明了氨基酸自動分析的方法,逐漸成為國內外測定氨基酸的常用方法。隨著研究的深入和分析儀器的發展,出現了電泳法、光譜法、色譜法、質譜聯用等更加快速準確檢測煙葉游離氨基酸的方法[26-27](表3)。國外對生物堿的測定方法研究較早,Bertrand 和Javillier 采用硅鎢酸重量法測定了煙堿;之后Markw 提出了1種煙堿比色測定的方法[28]。國內對煙堿含量的測定最初采用酸堿滴定法,之后出現了光度法、電泳法、液相色譜、氣相色譜、液質聯用或氣質聯用等更精準的測定方法[28-30],目前行業內主要采用連續流動分析法大批量檢測樣品中生物堿含量(表4)。
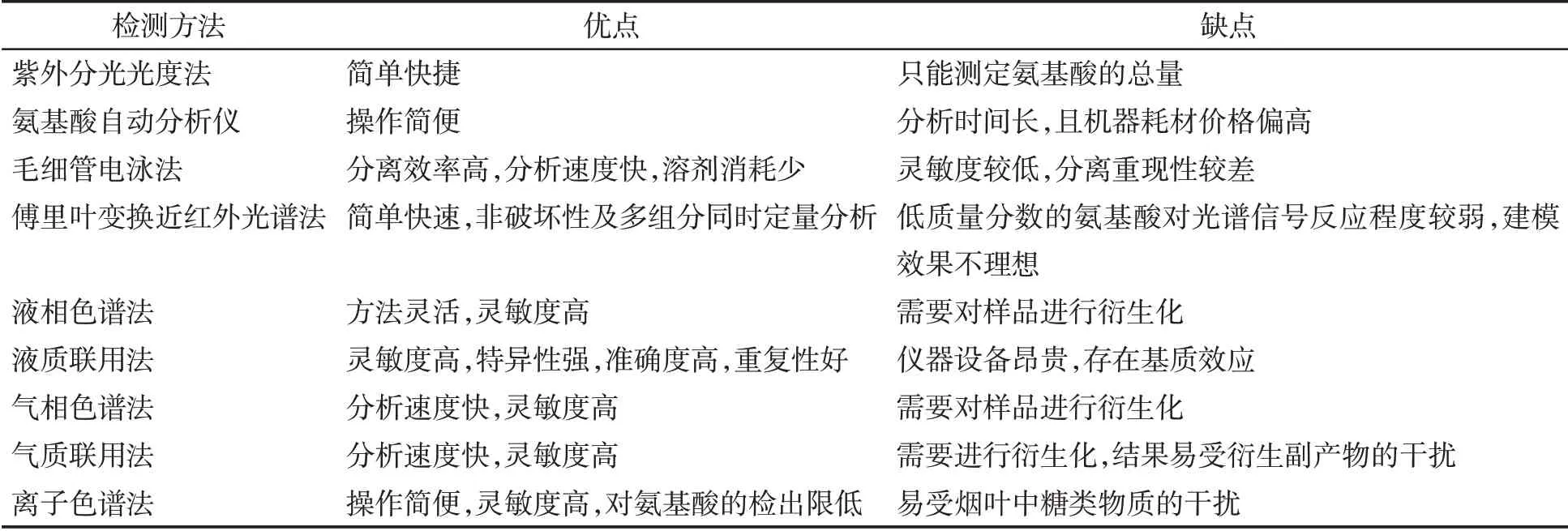
表3 煙葉中游離氨基酸的檢測方法Tab.3 Detection methods for free amino acids in tobacco leaves
1.2.2 含氮化合物與煙葉成熟度的關系
煙葉成熟過程中氮代謝產物的研究多集中在總氮、蛋白質、氨基酸和生物堿(表5)。隨著煙葉成熟度增加,總氮、蛋白質和游離氨基酸含量逐漸降低,這一變化與煙葉衰老過程中氮素的再分配和再利用有關[31]。葉片中的氮素大部分以蛋白質形式存在于葉綠體中,在葉片進入衰老階段,蛋白水解酶活性顯著增強,蛋白質被水解為氨基酸等物質,然后通過韌皮部輸送到新生器官中以維持其生長,因此蛋白質等含氮化合物含量在煙葉成熟衰老過程中逐漸降低[32]。在煙葉生長發育過程中,生物堿在葉片中逐漸積累,各組分(煙堿、降煙堿、新煙草堿和假木賊堿)含量均隨成熟度增加而呈上升趨勢[33]。游離氨基酸可作為前體物參與煙堿的生物合成,隨著煙葉葉齡的增加,氮在游離氨基酸中的分配比例逐漸降低,而在煙堿中的分配比例逐漸提高,成熟后期煙葉游離氨基酸含量降低,也可能與其參與了煙堿的生物合成有關[31,34]。有研究表明在煙葉成熟后期,煙堿含量出現下降趨勢,可能是由于煙葉達到了過熟狀態,葉片開始衰老,導致部分內含物質發生降解而損耗[35]。有研究認為,煙葉游離氨基酸含量與煙葉品質呈負相關,在煙葉成熟過程中游離氨基酸含量呈先降低后升高的變化規律,最低值出現在煙葉工藝成熟時期,煙葉中游離氨基酸含量可以作為判斷煙葉成熟度的依據之一[18]。與碳水化合物相似,煙葉成熟過程中含氮化合物的變化規律在不同品種、生態環境和栽培條件下也略有差異。因此,在實際生產中應針對產區特點,探究煙葉成熟過程中化學成分與成熟度的關系,確定煙葉最佳采收期,將煙葉化學成分控制在適宜范圍內。

表5 國內外煙葉成熟過程中含氮化合物的變化主要研究結果Tab.5 Domestic and foreign researches on variations of nitrogen metabolites during tobacco leaf ripening
1.3 致香物質
1.3.1 致香物質的種類及測定方法
煙葉致香物質含量是衡量烤煙品質的重要因素之一。國外早在20 世紀50 年代就已經從烤煙中分離鑒定出多種重要的香味成分如茄尼醇、新植二烯和揮發性脂肪酸等[41]。1976年,Lloyd等[42]借助蒸餾萃取和氣相色譜/質譜聯用技術(GC/MS)從烤煙煙絲中分離鑒定出酮類、羧酸類、醇類、醛類、酚類和氮雜環類等共計323 種化合物。國內對烤煙致香成分的研究較晚,1989年,李云等[43]利用氣相色譜技術從烤煙中鑒定出12種低級脂肪酸;1992年,冼可法等[44]從云南烤煙中鑒定出129 種中性致香成分。隨著前處理技術以及檢測技術的不斷發展,同時蒸餾萃取、固相微萃取、液液微萃取、超臨界CO2萃取、加速溶劑萃取以及基于氣質和液質聯用的代謝組學分析手段逐漸應用到煙葉致香成分分析中,大量致香成分被分離和鑒定出來[45]。
1.3.2 致香物質與煙葉成熟度的關系
一般認為在煙葉成熟過程中,類胡蘿卜素、類西柏烷和苯丙氨酸等致香前體物的降解產物以及棕色化反應產物含量總體呈升高趨勢,達到最大值后開始下降,但由于煙葉致香物質受生態因素影響較大,不同產區的研究結果存在較大差異,例如在四川省攀西地區,隨著成熟度增加,煙葉棕色化產物含量逐漸降低[46],而在陜西省延邊地區,煙葉棕色化產物總量則呈上升趨勢[47]。此外,不同品種的致香物質成分組成也存在差異,在烤煙NC89煙葉成熟過程中檢測到糖苷類香氣成分共28種,包括醛、酮、醇、內酯和酚類等成分,而在云煙87煙葉中僅分離鑒定出21種糖苷類香氣成分,主要由脂肪醇、芳香族成分、倍半萜類和多酚類物質組成,兩個烤煙品種煙葉中糖苷類香氣成分總量均隨成熟度的提升而增加[48-49]。目前,關于煙草致香物質的研究大多集中在烤后煙葉中,對于田間鮮煙葉尤其是上部煙葉成熟過程中的變化研究較少,雖然在煙葉烘烤過程中致香物質含量還會發生新的變化,但田間采收煙葉中致香物質及前體物的基礎含量對煙葉品質起著決定性作用。因此,選擇合理的采收期,保持適宜的煙葉致香物質及前體物含量,對改善烤后煙葉香氣品質具有重要意義。
1.4 質體色素
煙葉成熟過程中最直觀的變化是顏色的改變,生產上葉片顏色是判斷成熟度的重要依據。煙葉顏色的變化實質上是質體色素含量變化的外在表現,質體色素主要包括葉綠素和類胡蘿卜素等。葉綠素含量下降而導致的葉片失綠被認為是植物衰老最顯著的特征,其與葉綠體中類囊體膜逐漸崩解有關[50]。觀察細胞超微結構(圖1)發現,在煙葉成熟過程中,細胞內葉綠體首先出現衰老癥狀,具體表現為細胞空隙變大,葉綠體腫脹呈不規則形狀,基粒個數和類囊體數量逐漸減少,類囊體膜結構喪失,淀粉粒和嗜鋨顆粒數量增多、體積增大,并向細胞中部游離[50-51]。
在煙葉成熟過程中,葉綠素、類胡蘿卜素和質體色素總量均隨成熟度增加而逐漸降低,其中葉綠素較類胡蘿卜素含量下降速率更快,降解量更大,二者之間的比例變化使不同成熟度煙葉顏色產生差異[52]。由于煙葉葉綠素含量在成熟過程中變化顯著且易于測定,所以常被作為衡量煙葉成熟度的重要指標。葉綠素儀可以快速檢測葉片中葉綠素相對含量(Soil and Plant Analyzer Development,SPAD),生產上可將煙葉田間SPAD值與其外觀顏色特征相結合,作為判斷煙葉成熟度的依據[53-54]。然而,SPAD儀雖然操作簡單、使用方便,但只能進行單點測量,測量誤差和樣本代表性的缺陷,限制了其大范圍推廣使用。近年來,多種現代信息獲取技術如計算機視覺技術、無人機遙感技術、高光譜技術和近紅外光譜技術等逐漸在煙草生產中得到應用,通過建立光譜信息與煙葉成熟度的關系模型,可以實現煙葉成熟度的可視化分析[55-57]。無人機遙感和高光譜等技術雖然具有測量范圍大、精準度高等優勢,但技術門檻和成本較高,現階段并未廣泛應用于煙葉成熟度判斷。因此,根據田間煙葉理化指標的變化選擇更加科學合理的成熟度量化指標和便捷高效的檢測方法、檢測設備,將是今后需重點解決的問題。
2 上部煙葉成熟過程中生物酶活性變化
2.1 碳代謝相關酶
植物的碳氮代謝過程離不開各種生物酶的參與,在煙葉生長發育過程中,參與碳代謝的生物酶有淀粉合成酶(Starch synthase,SS)、淀粉酶(Amylase,AM)、蔗糖轉化酶(Invertase,INV)和蔗糖合成酶(Sucrose synthase,SUS)等[58]。AM 是碳水化合物積累代謝過程中的關鍵酶,可以將煙葉中積累的淀粉降解為麥芽糖和少量葡萄糖,目前關于煙葉AM 活性在成熟過程中的變化主要有3 種觀點:①雙峰變化,但達到峰值的時間還存在分歧[9,12];②先升高后降低的單峰變化[58-59];③先降低后升高的“U”形曲線變化,最低值時間與淀粉含量達到最高值的時間相吻合[21],結果的差異主要與烤煙品種、生態環境及取樣標準等不同有關。蔗糖是植物體中碳水化合物運輸的主要形式,INV 可以催化蔗糖水解為葡萄糖和果糖,其活性大小反映了煙葉對光合產物的利用程度。楊志曉等[59]研究表明,煙葉INV 活性在成熟前期較低,呈緩慢增加趨勢,在成熟期活性迅速上升,不同品種間INV活性存在較大差異。INV活性前期較低,可能是因為煙葉前期氮代謝活動占主導,碳代謝活動強度較弱,進入葉片功能盛期后,氮代謝活動開始減弱,轉為以碳代謝為主,轉化酶活性增強有利于促進光合產物的合理分配。
2.2 氮代謝相關酶
植物的氮代謝包括氮素的吸收、同化、轉運、利用和調節等過程。參與氮代謝的生物酶有硝酸還原酶(Nitrate reductase,NR)、谷氨酰胺合成酶(Glutamine synthetase,GS)、谷氨酸合成酶(Glutamate synthase,GOGAT)和谷氨酸脫氫酶(Glutamate dehydrogenase,GDH)等[59]。1952 年,Evans 等[60]在紅色鏈孢霉中首次發現NR,次年又在高等植物中發現。NR 是植物氮代謝的關鍵酶和限速酶,其作用是將植物吸收的硝態氮還原為亞硝態氮以供后續利用。2000年,Becker等[61]將植物GS分為2 類,細胞質型(GS1)存在于根部細胞質中,主要同化植物根系系統中硝酸還原作用產生的氨;質體型(GS2)存在于綠色組織質體中,負責同化硝酸還原和光呼吸所產生的氨。GS 與GOGAT 聯合作用下,將植物吸收的無機態氮同化為有機態的谷氨酰胺和谷氨酸,其活性大小可反映植物對氮素的同化利用情況[62]。隨著煙葉成熟度增加,NR活性先升高后降低,在移栽后30~60 d活性較強,在60~70 d后活性逐漸減弱,亞硝酸還原酶活性與NR活性變化規律大致相同[40]。賈保順等[63]研究發現,烤煙K326 和NC71的GS活性在移栽后70~110 d內呈先升高后下降的變化,最高值出現在移栽后80 d,葉片中GDH活性、NH4+濃度與GS活性變化趨勢相同。氮代謝相關酶活性前期升高可能與葉片正處在功能盛期有關,此時氮代謝活動較強,后期碳代謝活動逐漸增強,氮代謝活動開始減弱。在煙葉成熟進程中,碳氮代謝的強度、協調程度以及動態變化會直接影響煙葉中各類化學成分的含量和比例,進而影響煙葉品質。優質煙葉生產的關鍵是在適當發育時期,及時由以氮代謝為主轉變為以碳積累代謝為主。因此,生產上可采用生物、化學和農藝等措施有針對性地對碳氮代謝相關酶活性進行調節,以協調碳氮代謝進程,提高上部煙葉化學成分協調性,改善煙葉品質。
2.3 衰老相關酶
衰老是植物發育過程中細胞、組織、器官和個體死亡的過程。在眾多衰老機制學說中,生物自由基導致衰老的學說比較受重視[64]。植物體內的超氧化物歧化酶(Superoxide dismutase,SOD)、過氧化氫酶(Catalase,CAT)和過氧化物酶(Peroxidase,POD)等是植物應對脅迫時重要的防御酶系,可以有效阻止活性氧自由基的積累[65]。SOD是生物體轉移清除超氧陰離子自由基(O2-·)的酶,能特異性地將O2-·歧化為H2O2和O2[66]。CAT 是一種包含血紅素的四聚體酶,在煙草中分離到的3個CAT同工酶中,Cat1主要負責清除光呼吸產生的H2O2,Cat2 則清除氧化脅迫產生的H2O2,而Cat3 主要清除乙醛酸循環體中產生的H2O2[67]。植物中的過氧化物酶有很多種,最常見的有抗壞血酸過氧化物酶、谷胱甘肽過氧化物酶和辣根過氧化物酶等。利用電泳法從辣根組織中分離出2 種不同的辣根過氧化物酶,在活性氧代謝過程中,POD可以清除光合作用過程中產生的過量H2O2,起到保護葉綠體的作用[68]。
植物衰老過程其實也是抗氧化酶活性下降、活性氧自由基積累、膜脂過氧化不斷加劇的過程。在煙葉衰老過程中,SOD、CAT和POD的活性一般呈逐漸降低的趨勢,但也有研究發現,在煙葉成熟前期抗氧化酶活性有短暫升高的現象[69-70]。此外,一些參與煙株防御體系的生物酶,如脂氧合酶和多酚氧化酶等在煙葉成熟過程中也呈現先升高后下降的變化趨勢[70-71]。成熟前期煙株代謝旺盛,活性氧的少量積累對植物細胞損害較小,反而誘導了抗氧化酶活性的升高,但在成熟后期,煙株生理功能逐漸衰退,活性氧過量積累造成細胞大量損傷,抗氧化酶活性也隨之降低[72]。外界環境因素的改變會影響煙葉衰老進程,如高溫、冷害、鹽脅迫等在一定程度上會加速煙葉衰老。噴施外源調節物質,如水楊酸、硅、2,4-表油菜素內酯等可以調節植物葉片抗氧化酶活性,完善植物體內的防御體系,緩解惡劣環境脅迫對煙株造成的傷害[73-74]。因此,生產上遭遇環境脅迫條件時,可以根據大田煙株生長實際情況,選擇適宜的外源調節物質進行噴施,以增強成熟期煙株的抗逆性。
3 上部煙葉成熟過程中基因表達變化
轉錄組是指細胞中所有參與蛋白質翻譯的mRNA 總和,自20 世紀90 年代起,隨著分子生物技術和高通量測序技術等的快速發展,轉錄組學技術已廣泛應用于各個學科領域,借助轉錄組學分析技術,科研人員在植物葉片衰老的分子機制方面取得了重大進展。1995 年,Gan 等[75]將農桿菌ipt基因在衰老啟動子SAG12 的驅動下在煙草中表達,延緩了煙葉衰老的進程;2001 年,Yang 等[76]發現煙草中兩種堿性亮氨酸拉鏈(b-ZIP)家族的轉錄因子表達會促進葉片衰老。目前,已從煙草和擬南芥等模式植物中克隆出大量與衰老相關的基因,根據這些基因在葉片衰老期間表達量的變化,可將其分為衰老上調基因和衰老下調基因[77]。相關基因在煙葉成熟過程中的功能主要集中在蛋白質降解、碳氮代謝、色素代謝和激素代謝等方面。
3.1 蛋白降解相關基因
半胱氨酸蛋白酶是一種多功能水解蛋白酶,在植物衰老和細胞程序性死亡過程中起關鍵作用,煙草中NtCP1、NtCP2、NtCP23和MC是編碼半胱氨酸蛋白酶的基因[50]。2006 年,Beyene 等[78]從煙草衰老葉片和非衰老葉片中分離得到兩個與衰老相關的半胱氨酸蛋白酶基因NtCP1和NtCP2,NtCP1是一種特異性表達基因,僅在自然衰老的葉片中表達,且不受外界不良條件的誘導,被認為是煙草中良好的衰老標記基因,NtCP2與NtCP1表達模式不同,其在成熟綠葉中表達量較高,而在衰老葉片中表達量顯著降低。在衰老葉片中,包括NTCP23在內的許多編碼蛋白酶基因上調以參與氮素的轉移活動。2000 年,Ueda等[79]從煙草中分離出編碼半胱氨酸蛋白酶的基因NtCP23,其表達模式與NtCP1相似,在衰老過程中上調表達,但在葉片生長發育初期也能檢測到NtCP23的表達。PSA1是編碼26S 蛋白酶體的非催化型亞基,在煙葉衰老過程中其表達量逐漸降低,有學者推測蛋白酶體在植物細胞中的作用可能是調節蛋白水平來控制發育過程,而不是在衰老過程中降解蛋白質[50]。在煙葉衰老過程中,NtCP1、NtCP23、PSA1和MC在不同烤煙品種中的變化規律基本一致,基因NtCP1和NtCP23在衰老過程中上調表達,PSA1和MC基因下調表達。不同烤煙品種之間的差異在于各基因表達量的不同,成熟落黃快的品種NtCP1和NtCP23的表達量較高,PSA1和MC的表達量較低[22]。
3.2 碳氮代謝相關基因
關于煙葉成熟過程中碳氮代謝相關基因的研究主要以編碼生物酶的基因為主。顆粒結合淀粉合成酶(GBSS)和淀粉分支酶(SBE)是淀粉合成的關鍵酶,分別參與直鏈淀粉和支鏈淀粉的合成,他們由相應的淀粉合成酶基因GBSS1、SBE等編碼。在煙葉成熟過程中,GBSS1和SBE基因的相對表達量呈先升高后下降的變化,與淀粉含量在成熟過程中的變化一致[22]。蔗糖合成酶基因(NtSS)、蔗糖磷酸合成酶基因(NtSPS)和蔗糖轉化酶基因(NtINV)參與煙葉的糖代謝過程。在煙葉成熟過程中,NtINV表達量大致呈單峰曲線變化,NtSS和NtSPS表達量均有不同程度上調,且品種之間變化規律一致,說明煙葉進入成熟期后糖代謝活動逐漸增強[80]。在氮代謝方面,NR 是整個氮代謝過程中的限速酶和關鍵酶,其編碼基因在葉片衰老過程中表達量逐漸降低[81]。周健飛等[82]指出,在煙葉成熟過程中,NR 基因的表達豐度僅與NR 活性呈極顯著正相關,對成熟期氮效率影響較小,成熟期煙葉氮素運籌主要受GS同工酶基因表達豐度的影響。在煙葉衰老過程中,GS1 調控氮素的轉移及再利用,GS2 負責氨同化作用[83],GS1 編碼基因表達量呈上升趨勢,GS2 編碼基因表達量則逐漸下降,表明隨著煙葉衰老其氮素轉移能力逐漸加強,而氮素同化能力逐漸減弱[22,82]。GDH在煙葉衰老過程中主要負責脫氫,編碼GDH的基因有GDH1和GDH2,它們的表達量在煙葉衰老過程中均增加,其轉錄水平在衰老后期最高[50],但也有研究發現GDH1的相對表達量在衰老過程中呈先升高后降低的趨勢[81],相關差異的原因還需進一步研究。
3.3 色素代謝相關基因
在煙葉成熟過程中,與類胡蘿卜素合成相關的基因GGPS、PSY、PDS、ZDS和CRTISO均呈現下調表達,相關轉錄因子ORANGE、HY5、COP1和DET1也呈現下調表達。與之相反,類胡蘿卜素轉化基因NCED、ZEP、NXS以及與類胡蘿卜素降解相關的基因LOX、POD、CCD1則上調表達[84]。葉綠素酶是分解葉綠素的初始酶,CLH是編碼葉綠素酶的基因,在煙葉衰老過程中,CLH表達整體增強,說明在煙葉成熟過程中,葉綠素與類胡蘿卜素的分解代謝逐漸增強,類胡蘿卜素的合成逐漸減弱,這與煙葉中質體色素含量降低的變化規律一致[85]。目前,從擬南芥和水稻中鑒定出6種葉綠素代謝酶[86],相關基因在葉片衰老過程中的變化在其他作物上有較多研究[86-87],而在上部煙葉成熟過程中的變化研究則相對較少。
3.4 激素代謝相關基因
內源激素是植物體內重要的信號物質,對葉片的成熟衰老進程起重要調控作用。煙葉成熟過程中,參與乙烯、脫落酸和茉莉酸合成的相關基因NtEFE26、NtNCED和NtPR1b表達量均呈升高趨勢,在衰老煙葉中達到最高值,與相應激素含量的變化趨勢相同[88]。生長素(IAA)和細胞分裂素(CTK)被認為是延緩衰老的激素,在煙葉衰老過程中,IAA負調控因子AuX/IAA與SAUR家族蛋白的相關基因表達量均呈升高趨勢,IAA合成受到抑制,這與IAA含量在煙葉衰老過程中下降的變化相一致[89]。在衰老葉片中CTK含量顯著下降,轉錄組分析結果表明,與CTK降解有關的基因上調表達,而與CTK合成相關的基因則下調表達[90]。NtCHN50是水楊酸應答基因,在煙葉成熟過程中其表達量逐漸降低,預示著水楊酸合成減弱[88],但對擬南芥的研究發現,在衰老葉片中水楊酸的濃度提高了4倍,衰老葉片中較高的水楊酸含量似乎與葉片衰老過程中幾種SAG的上調表達有關[91],差異的原因還需進一步研究。
4 上部煙葉成熟過程中蛋白表達變化
4.1 蛋白組學技術在煙草上的應用
自20世紀90年代起,伴隨著蛋白組學技術體系的不斷發展,蛋白組學技術已廣泛應用于植物不同生長發育階段以及在各種環境因子和逆境脅迫下的蛋白表達差異研究中[92]。2008年,Hebeler等[93]用蛋白組學技術分析了擬南芥早期衰老葉片的蛋白豐度變化,鑒定出13個差異顯著的蛋白。2010年,Bai等[94]研究了牽牛花衰老過程中蛋白組的變化,發現許多衰老上調蛋白參與了防御和應激反應等生物活動。蛋白組學技術在煙草上的應用起步相對較晚,目前主要集中在不同基因型、不同生態環境以及脅迫條件下差異蛋白的研究。2008 年,崔紅等[95]對濃香型和清香型典型生態區的煙葉進行蛋白組學比較分析,發現有51個蛋白在兩個生態區發生了差異表達,在福建煙區高表達的蛋白主要參與葉綠體發育、色素代謝和光合作用,而在河南煙區高表達的蛋白主要參與糖代謝途徑,為從蛋白水平解釋不同生態區煙葉品質差異提供了參考。2016 年,徐瑩等[96]對云煙87、K326和紅花大金元成熟期煙葉蛋白組分析發現,云煙87與烤煙K326煙葉中蛋白質表達圖譜相似,僅存在29 個差異表達蛋白,而云煙87 和紅花大金元之間則有160個蛋白質差異表達,差異蛋白多與光合作用、物質代謝和抗逆性有關。2019 年,Chen等[97]基于同位素相對標記和絕對定量(iTRAQ)的蛋白組學分析研究表明,豫煙6號和云煙87 在水分脅迫下蛋白組表達水平存在差異,與云煙87相比,豫煙6號表現出更強的保水能力、更好的光合性能、更強的抗氧化防御能力,為研究煙草干旱脅迫反應相關的遺傳調控機制奠定了基礎。
4.2 煙葉成熟衰老過程中的蛋白組學研究
近年來,在蛋白水平上解釋煙葉成熟過程中的成分變化逐漸受到人們的關注。2014 年,張柳等[98]采用蛋白組學技術對云南玉溪地區云煙87旺長期、成熟期和成熟后期煙葉的蛋白組進行了分析,共檢測到432 個差異表達蛋白,主要集中在以下幾個方面:(1)光合作用,如與光反應相關的蛋白質(如PSⅠ和PSⅡ反應中心蛋白等)和與暗反應中CO2同化相關的蛋白(Rubisco及其活化酶等)在衰老過程中均呈下調表達。(2)糖酵解代謝,如α-葡聚糖焦磷酸化酶、α-葡萄糖苷酶類、與糖類轉化及糖核苷酸合成相關酶類均上調表達。(3)蛋白質和氨基酸分解代謝,如天冬氨酸蛋白酶、天冬氨酸轉氨酶等上調表達。(4)逆境反應,如熱激蛋白和分子伴侶總體上調表達。(5)脂類代謝,如甘油酯水解酶類和磷脂類水解酶表達水平升高。這些現象表明,在煙葉衰老過程中葉片光合作用強度逐漸減弱,呼吸代謝、物質分解代謝活動逐漸增強,同時抗逆相關蛋白表達的升高說明煙葉衰老對植物體本身也是一種逆境反應。何聰蓮等[99-100]采用iTRAQ對烤煙K326未熟、適熟和過熟期煙葉的蛋白組進行分析,共鑒定出86個共同差異表達蛋白,這些差異蛋白涉及代謝過程、細胞過程和應激反應等,最終篩選出 包 括Q1WL43、A0A1S4DCM1、A0A1S4C433、A0A1S4AKK6、A0A1S3ZQP6 和A0A140G1P2 在內的6個差異蛋白,可能是煙葉衰老潛在靶標蛋白。
5 總結與展望
目前,我國在上部煙葉成熟度研究和應用方面仍存在一些問題:①關于成熟度與煙葉品質關系的研究,主要集中在田間煙葉成熟度對烤后煙葉品質的影響,而對成熟過程缺乏關注,田間煙葉成熟過程中物質轉化規律和內在生理生化研究不夠深入。②上部煙葉的采收成熟度判斷仍然靠主觀經驗,缺乏統一的特征指標、判斷標準和便捷的測定方法,在生產操作中不易把控,上部煙葉采青現象仍然存在;部分產區由于過分強調成熟度,導致上部煙葉出現物質消耗過度、品質下降等問題。③國內許多產區主要采用延遲采收技術提高煙葉成熟度,由于在煙葉生長后期部分產區氣溫下降較快,導致煙葉表面易形成蠟質層,使煙葉難以成熟,給烘烤帶來困難。煙葉生育后期降水增多帶來的病害問題也限制了該項技術的應用。④在外源調節劑調控上部煙葉開片和成熟度方面研究尚不深入,應用也較少。
針對上述問題,未來可先從典型產區和主要栽培品種入手開展相關研究。①借助轉錄組、蛋白組和代謝組學的相關技術手段,從關鍵酶活性、基因表達、蛋白質和生物小分子代謝物等多角度出發,系統研究上部煙葉成熟過程中的物質代謝規律,篩選煙葉成熟過程中起重要調控作用的關鍵基因和代謝產物,并進行功能驗證和機制解析。②深入開展烤煙上部煙葉成熟調控研究,研制篩選對烤煙上部煙葉成熟具有較好調控作用的生長調節劑,并揭示其作用機理。③選擇適宜種植區域,保證煙株營養均衡,提高煙葉耐熟性,以煙株營養均衡+成熟采收為核心提高上部煙葉品質。④在煙葉田間成熟度特征指標、判斷標準、測定方法和測定設備等方面加強研究,針對不同的生態區域,形成上部煙葉成熟度判斷和采收標準,制定精準的上部煙葉生產技術規范。
