偽狂犬病毒感染對BHK-21懸浮細胞內質網應激和未折疊蛋白反應的影響
2022-08-23 02:40:10倪敏舒莊騰寒郭美錦
中國畜牧獸醫 2022年8期
陳 麗,倪敏舒,徐 悅,鮑 熹,莊騰寒,馮 磊,3,4,5,郭美錦
(1.華東理工大學生物反應器工程國家重點實驗室,上海 200237;2.江蘇省農業科學院動物免疫工程研究所,南京 210014;3.江蘇省動物重要疫病與人獸共患病防控協同創新中心,揚州 225009;4.江蘇大學藥學院,鎮江 212013;5.江蘇省食品質量安全重點實驗室——省部共建國家重點實驗室培育基地,南京 210014)
病毒在感染過程中會把內質網(ER)作為復制場所,并利用宿主細胞內質網的膜和蛋白系統合成自身的病毒蛋白。內質網是真核細胞器的重要組成部分,由脂類和蛋白質構成,是蛋白質合成、翻譯后修飾、加工和折疊的重要場所,同時還參與Ca2+儲存和脂類物質的合成[1]。在缺氧、氧化應激、毒性物質刺激等情況下[2],內質網的微環境發生改變,大量未折疊或者錯誤折疊蛋白在內質網中聚集,使內質網處于生理功能紊亂,進而誘發內質網應激。為緩解應激壓力,細胞會啟動未折疊蛋白反應(UPR)以維持細胞穩態[3]。為了能夠達到在宿主體內增殖,病毒需通過調控UPR以減少對其增殖的不利因素,創造出有利條件。同時,病毒還可以通過調控UPR與細胞凋亡、細胞自噬和病毒先天免疫之間的關系,以應對病毒感染引起的內質網應激,最終達到致病的目的[4-5]。UPR活化由3個不同的內質網跨膜蛋白介導,分別是蛋白激酶樣內質網激酶(PERK)、ER跨膜蛋白激酶1(IRE1)和活化轉錄因子6(ATF6)[6-7]。跨膜蛋白具有抑制病毒蛋白的合成、降解駐留在內質網的病毒蛋白量或誘導促折疊伴侶分子表達等功能[8-9]。活化的PERK催化真核轉錄起始因子2(eIF2α)磷酸化,磷酸化eIF2α(P-eIF2α)上調活化轉錄因子4(ATF4)基因的表達,ATF4激活生長停滯和DNA損傷誘導蛋白(GADD34)、C/EBPα-同源蛋白(CHOP)等相關蛋白的表達[10]。活化的IREl能夠去除X盒結合蛋白1(XBP1)mRNA的26 bp內含子,形成翻譯框移碼,編碼新蛋白sXBP1[11-12]。XBP1被IRE1切除的26 bp核苷酸序列中含有1個PstⅠ酶切位點[13]。剪切形式的sXBP1誘導內質網相關降解途徑(ERAD)中伴侶分子和內質網相關降解蛋白(EDEM)的表達。活化的ATF6(90 ku)轉運至高爾基體后降解為有轉錄活性的ATF6蛋白(50 ku),進入細胞核調節內質網應激響應元件(ERSE)的表達,如葡萄糖調節蛋白78(GRP78)、葡萄糖調節蛋白94(GRP94)、鈣連蛋白(Calnexin)和鈣網蛋白(Calreticulin)等[14]。多種皰疹病毒可以引起內質網應激,如單純皰疹病毒1型(Herpes simplex virus 1,HSV-1)通過PERK通路應對內質網應激[15-16];鴨瘟病毒(Duck enteritis virus,DEV)感染可以激活PERK和IRE1通路,并通過小干擾RNA(siRNA)檢測發現這2個通路有助于誘導細胞自噬[17]。
偽狂犬病(pseudorabies,PR)是一種由偽狂犬病毒(Pseudorabies virus,PRV)引起的急性傳染病[18-19]。PRV屬于α皰疹病毒亞科,具有傳播速度快、死亡率高、病原體頑固等特點[20-21],能夠引起多種家畜和野生動物感染發病,主要的宿主是豬[22-23]。該病廣泛分布于世界各地多個國家和地區,給全球養豬業帶來了巨大的經濟損失。疫苗的免疫接種是防治和消滅偽狂犬病的主要手段[24]。PRV致病機制目前還不明確,內質網應激研究對PRV與宿主細胞的相互作用機制的研究具有重要意義,可為闡明PRV的致病機制奠定基礎。目前有關PRV感染誘導內質網應激的研究較少,Yang等[25]的研究表明,PRV感染PK-15細胞在感染早期可激活IRE1-XBP1通路,但是該通路沒有對病毒復制造成影響,并通過CHOP-Bcl2通路誘導細胞凋亡。作者前期對PRV感染BHK-21懸浮細胞的接毒劑量進行了摸索,考察了MOI為0.1,0.01,0.001,0.0001對病毒滴度影響,最終得到在生物反應器中生產PRV的最佳MOI為0.01(未發表)。本研究以PRV滅活疫苗生產中的工程細胞BHK-21懸浮細胞為宿主細胞,以MOI 0.01接毒,考察PRV感染對BHK-21懸浮細胞的內質網應激及UPR信號通路的影響。
1 材料與方法
1.1 材料
1.1.1 細胞和毒株 BHK-21貼壁細胞、BHK-21懸浮細胞和PRV毒株均由江蘇省農業科學院動物免疫工程研究所細胞工程創新團隊保存。
1.1.2 主要試劑 DMEM培養基、新生牛血清(NBS)均購自Gibco公司;懸浮培養基MD910購自北京清大天一科技有限公司;eIF2α(9722S)、P-eIF2α(3597S)和α-tubulin(2144S)抗體均購自Cell Signaling Technology公司;HRP標記羊抗兔二抗購自Invitrogen公司;內質網應激誘導劑毒胡蘿卜素(Tg)和內質網應激抑制劑牛磺熊去氧膽酸(TUDCA)均購自Sigma公司;PstⅠ內切酶購自TaKaRa公司;RNA提取、逆轉錄、QuantiFast SYBR Green PCR試劑盒和DNA回收試劑盒均購自Qiagen公司;CCK-8試劑盒購自上海碧云天生物技術有限公司。
1.2 PRV在BHK-21懸浮細胞中的增殖
在高壓滅菌后的1 L生物反應器中加入500 mL細胞培養液(含1% NBS的MD910懸浮培養基),將BHK-21懸浮細胞以5×105/mL接種,培養溫度為37 ℃,攪拌轉速為160 r/min,溶解氧(DO)為50%,pH為7.2。當細胞密度達到4×106/mL后再加入500 mL細胞培養液,以MOI為0.01接種PRV,以不接毒細胞作為對照組,分別在12、24、36、48和56 h收集對照組和感染組細胞,用CCK-8法檢測細胞存活率,按Reed-Muench法[26]測定病毒滴度,篩選病毒接種處理的合適時間。
1.3 PRV感染對內質網應激及UPR通路相關基因及蛋白表達的影響
1.3.1 樣品收集 按1.2方法接種病毒,分別在在12、24、36和48 h收集對照組和感染組細胞,用于檢測內質網應激標志分子GRP78及UPR的3種感受蛋白(PERK、IRE1、ATF6)相關通路中基因和蛋白的表達變化。
1.3.2 實時熒光定量PCR檢測相關基因mRNA的表達水平 使用RNA提取試劑盒提取1.3.1收集細胞的RNA,并反轉錄合成cDNA。參照GenBank中敘利亞倉鼠GRP78、ATF4、GADD34、p58IPK、EDEM、ERp57、PDI、Calnexin和Calreticulin基因序列,用Primer Premier 5.0軟件設計引物,引物信息見表1,引物均由南京擎科生物科技有限公司合成。以β-actin為內參基因,進行實時熒光定量PCR。PCR反應體系20 μL:SYBR Premix ExTaqⅡ 10 μL,上、下游引物(20 μmol/L)各1 μL,cDNA 1 μL,ddH2O 7 μL。PCR反應條件:95 ℃預變性5 min;95 ℃變性30 s,60 ℃退火45 s,72 ℃延伸45 s,共40個循環。采用2-ΔΔCt法計算各基因的相對表達量。
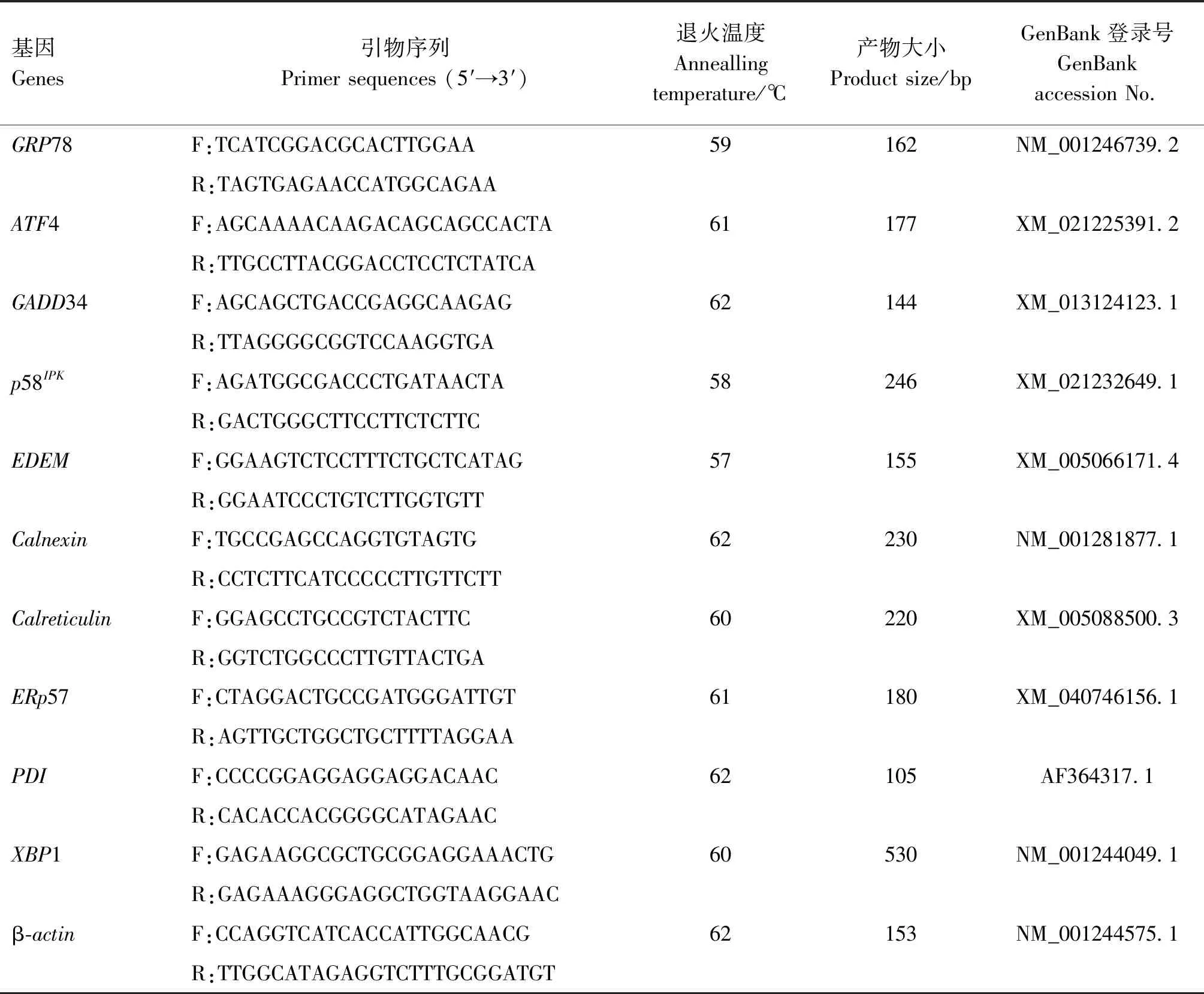
表1 引物序列
1.3.3 Western blotting檢測相關蛋白的表達 用RIPA裂解1.3.1收集的細胞提取總蛋白,12% SDS-PAGE分離蛋白,使用濕轉法將蛋白轉至PVDF膜上。5%脫脂牛奶在37 ℃封閉2 h,4 ℃一抗P-eIF2α(1∶1 000)、eIF2α(1∶1 000)和Tublin(1∶1 000)孵育過夜。然后用TBST洗滌3次,加入HRP標記羊抗兔二抗(1∶5 000)37 ℃孵育1 h,最后用ECL法顯示,并使用Tanon 5200系統成像拍照,ImageJ 2.0軟件統計蛋白條帶灰度值。
1.4 IRE1通路中XBP1剪切分析
用1.3.2中合成的各組細胞的cDNA,進行RT-PCR分析IRE1通路中XBP1剪切情況。根據敘利亞倉鼠XBP1基因序列(登錄號:NM_001244049.1),用Primer Premier 5.0軟件設計引物,引物信息見表1,引物由南京擎科生物科技有限公司合成。PCR反應體系20 μL:PremixTaq10 μL,上、下游引物(20 μmol/L)各1 μL,cDNA 1 μL,ddH2O 7 μL。PCR反應條件:95 ℃預變性5 min;95 ℃變性15 s,60 ℃退火30 s,72 ℃延伸30 s,共40個循環;72 ℃延伸300 s。PCR產物用2.0%瓊脂糖凝膠電泳后進行回收,回收產物用PstⅠ酶消化。sXBPl不含有PstⅠ酶切位點而不能被切開,電泳產物為504 bp,而非剪接形式的uXBPl則被PstⅠ酶切形成大小分別為317和213 bp的片段。
1.5 內質網應激相關化學藥物對PRV復制的影響
在BHK-21懸浮細胞上以MOI為0.01接種PRV,同時分別加入0(對照組)、0.001、0.005、0.01和0.02 μmol/L Tg和0(對照組)、20、40、80和160 μmol/L TUDCA,48 h收集細胞,按照1.2中的方法測定細胞存活率和病毒滴度。
1.6 數據統計與分析
用GraphPad Prism 5.0軟件進行單因素方差分析,樣本間差異采用t檢驗。結果以平均值±標準差表示。P<0.05表示差異顯著,P<0.01表示差異極顯著。
2 結 果
2.1 PRV在BHK-21懸浮細胞中的增殖
由圖1可知,PRV在BHK-21懸浮細胞中增殖速度較快,12 h病毒滴度達5.0 lg TCID50/mL,48 h病毒滴度最高,達到8.1 lg TCID50/mL,56 h細胞存活率低于70%,病毒滴度和細胞存活率均出現下降。因此后續試驗對PRV感染48 h內進行研究,分別在接毒后12、24、36和48 h收集細胞。

圖1 PRV感染BHK-21懸浮細胞后病毒滴度和細胞存活率變化Fig.1 Changes of viral titer and relative cell viability of BHK-21 suspension cells infected by PRV
2.2 PRV感染對GRP78表達的影響
由圖2可知,與對照組相比,PRV感染組GRP78 mRNA轉錄水平在36和48 h均極顯著上調(P<0.01)。
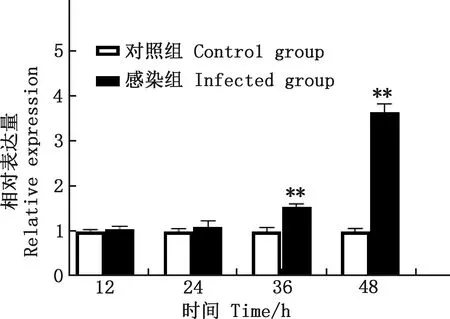
與對照組相比,*,差異顯著(P<0.05);**,差異極顯著(P<0.01);無*,差異不顯著(P >0.05)。下同Compared with control group,*,Significant difference(P<0.05);**,Extremely significant difference(P<0.01);No *,No significant difference(P>0.05).The same as below圖2 各組細胞不同時間GRP78的相對表達量Fig.2 The relative expression of GRP78 in each group at different time
2.3 PRV感染對PERK通路相關基因及蛋白表達的影響
由圖3可知,與對照組相比,ATF4在PRV感染36、48 h均極顯著升高(P<0.01),GADD34在PRV感染36 h顯著升高(P<0.05)、48 h極顯著升高(P<0.01);PRV感染36、48 h PERK通路中宿主細胞的P-eIF2α蛋白表達水平均極顯著增加(P<0.01)。
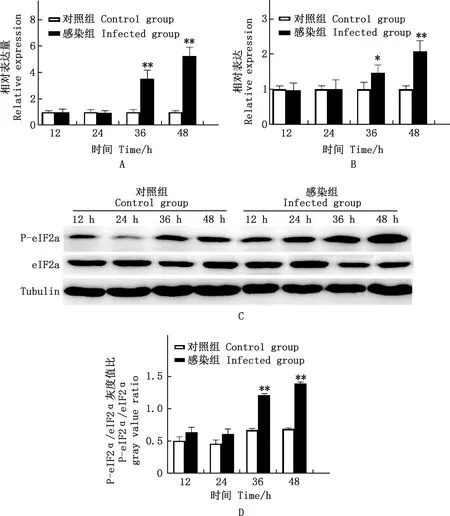
A、B,實時熒光定量PCR分析ATF4、GADD34相對表達量;C,Western blotting分析PERK通路P-eIF2α和eIF2α蛋白表達;D,P-eIF2α/eIF2α灰度值比A and B,The relative expression of ATF4 and GADD34 by Real-time quantitative PCR;C,The expression of P-eIF2α and eIF2α proteins in the PERK pathway by Western blotting;D,The ratio of P-eIF2α/eIF2α gray value圖3 各組細胞不同時間PERK通路相關基因及蛋白表達Fig.3 The related gene and protein expression of the PERK pathway in each group at different time
2.4 PRV感染對IRE1通路相關基因表達的影響
由圖4可知,與對照組相比,PRV感染組細胞36 h后多以剪切形式sXBP1存在,同時下游p58IPKmRNA水平在36和48 h顯著或極顯著增加(P<0.05;P<0.01),EDEM mRNA水平在48 h極顯著增加(P<0.01)。

A,RT-PCR分析XBP1剪切水平;B、C,實時熒光定量PCR分析p58IPK和EDEM相對表達量A,The level of XBP1 splicing by RT-PCR;B and C,The relative expression of p58IPK and EDEM by Real-time quantitative PCR圖4 各組細胞不同時間IRE1通路XBP1剪切水平及相關基因的相對表達量Fig.4 The relative expression of XBP1 splicing and the related gene of the IRE1 pathway in each group at different time
2.5 PRV感染對ATF6通路相關基因表達的影響
由圖5可知,與對照組相比,PRV感染組12、24、36、48 h ATF6下游相關效應分子ERp57、PDI、Calnexin和Calreticulin的表達均無顯著變化(P>0.05)。

A~D,分別為ERp57、PDI、Calnexin和Calreticulin基因A-D,ERp57,PDI,Calnexin,Calreticulin genes,respectively圖5 各組細胞不同時間ATF6通路相關基因的表達Fig.5 The related gene expression of the ATF6 pathway in each group at different time
2.6 內質網應激相關化學藥物對PRV復制的影響
由圖6可知,BHK-21懸浮細胞藥物處理48 h后,與對照組相比,0.01和0.02 μmol/L Tg極顯著降低細胞存活率(P<0.01),0.001和0.005 μmol/L Tg及所有試驗濃度的TUDCA組細胞存活率均無顯著差異(P>0.05);與對照組相比,0.005和0.01 μmol/L Tg均極顯著增加了PRV病毒滴度(P<0.01),20、40、80、160 μmol/L TUDCA均極顯著降低病毒滴度(P<0.01),且呈劑量依賴性降低。
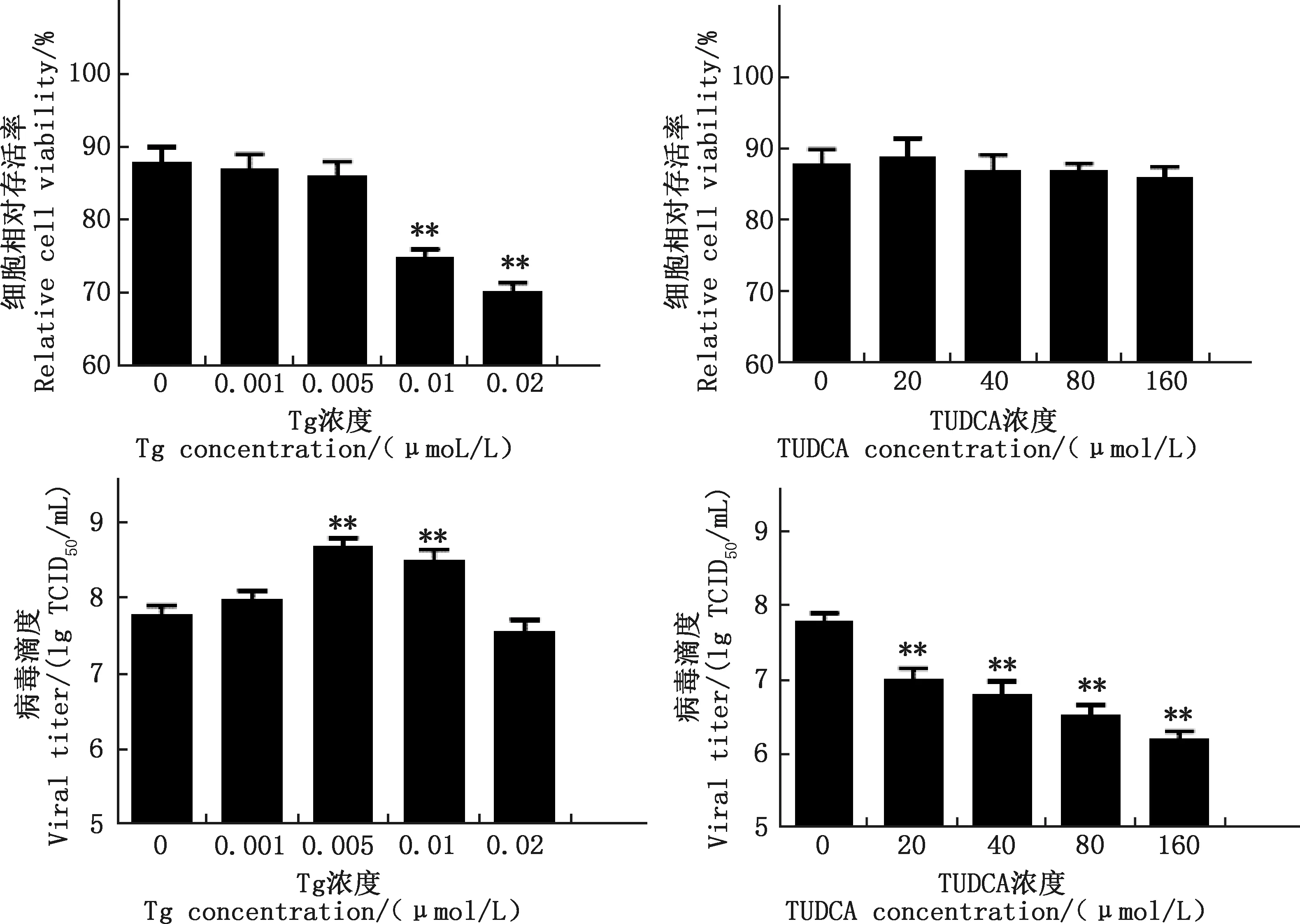
圖6 不同濃度Tg和TUDCA組細胞相對存活率及病毒滴度Fig.6 The relative cell viability and viral titers in different concentration groups of Tg and TUDCA
3 討 論
內質網是宿主蛋白合成的重要場所,病毒感染會引起宿主細胞內質網應激,為緩解內質網應激,宿主細胞會啟動UPR[27]。乙型肝炎病毒、流感病毒、豬圓環病毒、皰疹病毒等均可誘導內質網應激[28]。PRV在宿主細胞內可以快速地增殖。本研究發現,BHK-21懸浮細胞以MOI 0.01接種PRV,在48 h可達到最高效價,隨后細胞死亡率不斷增加,病毒滴度也隨之下降。內質網應激時通過UPR可使GRP78大量表達,GRP78可作為內質網應激的標志性分子。本研究中,在PRV感染BHK-21懸浮細胞36 h后,內質網應激標志分子GRP78表達量極顯著升高,說明出現內質網應激,UPR被激活。與其他報道相比,本研究中PRV感染BHK-21懸浮細胞后GRP78上調時間延遲,這可能與較低的MOI有關。本研究模擬使用規模化培養的生物反應器為容器,BHK-21懸浮細胞為宿主細胞,生產增殖PRV的MOI為0.01,而其他研究者大部分使用方瓶培養貼壁細胞,接毒的MOI為1.0[29-30]。
活化的PERK主要通過降低蛋白合成速率來緩解內質網應激。本研究檢測了PERK通路中eIF2a蛋白的磷酸化水平以及下游分子ATF4、GADD34轉錄表達水平,結果發現上述分子在PRV感染BHK-21懸浮細胞36 h后均出現表達量上升的現象。活化的IRE1主要通過核酸內切酶對XBP1進行剪切,降解內質網錯誤折疊蛋白以維持內質網穩態。在PRV感染宿主細胞36 h后,多以sXBPl剪切形式存在,同時下游調節分子p58IPK和EDEM轉錄水平均有所提高,但是ATF6通路中相關效應分子ERp57、PDI、Calnexin和Calreticulin的表達量并沒有明顯變化。這些結果說明PRV感染能夠激活BHK-21懸浮細胞的PERK和IRE1通路,對ATF6通路沒有影響。Yang等[25]報道PRV感染PK15細胞后可激活IRE1通路,但是并沒有明確激活PERK通路,造成這種現象的原因可能是不同的宿主細胞對內質網應激產生的反應不同。
病毒感染會產生內質網應激,并通過調節UPR創造對增殖有利的條件,以達到增強自身有效復制的目的。傳染性胃腸炎病毒(TGEV)感染可激活ST細胞和IPEC-J2細胞的UPR 3條信號通路,并通過PERK通路抑制TGEV復制[31]。Xu等[32]報道PRV體外感染小鼠神經細胞N2a可以誘導自噬來增強其自身復制。Catanzaro等[33]報道藍耳病(PRRSV)感染激活UPR的3條通路,但內質網應激抑制PRRSV的復制。經典豬瘟病毒(CSFV)利用內質網應激促進病毒增殖,其中IRE1通路起到了關鍵作用[34],并且CSFV非結構蛋白p7可以與宿主蛋白相互作用,調節內質網鈣離子的通透性[35]。這些研究表明,內質網應激在病毒增殖中具有重要作用。本研究在PRV感染過程中,添加內質網應激誘導劑Tg可明顯提高病毒滴度,而添加內質網應激抑制劑TUDCA則抑制病毒增殖,說明PRV可利用內質網應激促進病毒增殖。
4 結 論
PRV感染可以誘導BHK-21懸浮細胞內質網應激和UPR,激活PERK和IRE1信號通路,說明PRV感染可通過調節UPR以利于其在宿主細胞體內增殖。
