基因編輯技術在蔬菜中的研究進展
2022-08-27 03:52:10唐笑王桂香劉凡韓碩宗梅郭寧段蒙蒙
中國蔬菜 2022年8期
關鍵詞:研究
唐笑 王桂香劉凡 韓碩 宗梅 郭寧 段蒙蒙
(北京市農林科學院蔬菜研究所,農業農村部華北地區園藝作物生物學與種質創制重點實驗室,蔬菜種質改良北京市重點實驗室,北京 100097)
基因編輯是一種可以在基因組水平上對DNA序列進行改造的遺傳操作技術。目前,基因編輯技術已經在植物的遺傳基礎理論研究和生產實踐中得到廣泛應用。通過基因編輯技術進行基因功能研究和利用基因的靶向修飾進行性狀改良是基因編輯應用最廣泛的領域,在模式植物和大田作物中都有大量報道。蔬菜是人們獲取人體所需要的維生素和微量元素的主要來源,基因編輯技術目前已成功應用在多種蔬菜作物中。隨著功能基因組研究日漸深入,通過基因編輯技術進行功能基因研究和性狀改良是蔬菜分子育種的必然方向。
1 基因編輯技術的發展歷程
基因編輯的原理就是構建一個人工核酸內切酶,在預定的基因組位置切斷DNA,產生DNA 雙鏈斷裂(double-strand break,DSB),DSB 在被細胞內的DNA 修復系統修復過程中會產生突變,從而達到定點改造基因組的目的。基因編輯技術應用的關鍵是核酸內切酶的使用,常用的核酸內切酶能夠有效切割DNA,但它們通常在多個位點進行識別和切割,特異性較差。基因編輯的發展歷程就是研究者對不同類型核酸內切酶的發現和改造的過程。基因編輯常用的核酸內切酶主要分為四類:巨型核酸酶(meganuclease,MegNs)(Choulika et al.,1995)、鋅指核酸酶(zinc finger nucleases,ZFNs)(Wah et al.,1998)、轉錄激活因子樣效應物核酸酶(transcription activator-like effector nucleases,TALENs)(Zhang et al.,2014)和成簇規律性間隔短回文重復系統(clusters of regularly spaced short palindromic repetition systems,CRISPR/Cas)(Cho et al.,2013)。
1.1 巨型核酸酶
在20 世紀80 年代后期發現的巨型核酸酶是一種脫氧核糖核酸內切酶,可以識別一段12~40 bp的DNA 序列,這種專一的長序列在DNA 鏈上重復出現的頻率低,保證了其切割的特異性,但由于其對特定長序列的需求和巨大的酶體,導致其剪切效率低,應用難(Silva et al.,2011)。
1.2 鋅指核酸酶
由于巨型核酸酶的缺陷,鋅指核酸酶取代了巨型核酸酶。ZFNs 是一種人工融合蛋白,包括DNA識別域和核酸內切酶I。DNA 識別域由一系列Cys-His 鋅指蛋白(zinc-fingers,ZFPs)串聯組成,每個ZFP 識別并結合一個特異的三聯體堿基。Kim等(1996)首次將3 個串聯的鋅指結構域(能夠識別9 個核苷酸)與I 的C 端內切酶結構域通過一段連接蛋白融合,制造出第一個嵌合型核酸內切酶—ZFNs,并在體外證明該酶對靶DNA 具特異剪切能力。ZFNs 的DNA 識別域能識別特異位點并與之結合,而由I 構成的切割域具有剪切功能,針對目的基因設計特異性ZFPs,再將DNA 識別域與I 結合就可使靶位點產生DSB。細胞可以通過同源重組(HR)修復機制和非同源末端連接(NHEJ)修復機制來修復DNA。HR 修復有可能會對靶位點進行恢復修飾或者插入修復,而NHEJ修復極易發生插入突變或缺失突變,從而達到基因改造的目的。
基于ZFNs 的基因編輯技術可應用于很多物種及基因位點,具有較好的發展潛力。但是目前有3方面的缺陷制約了該技術的推廣:一是,以現有的策略設計高親和性的ZFNs,費時費力;二是,持續表達ZFNs 對細胞有毒性;三是,雖然三聯體設計具有一定特異性,但仍然存在不同程度的脫靶效應(李想 等,2017)。
1.3 轉錄激活因子樣效應物核酸酶
鑒于ZFN 技術的局限性,人們致力于尋找更為有效、特異性更強的基因編輯工具。2009 年,兩組研究人員同時報道了TALE 蛋白能特異性識別并結合DNA 序列(Boch et al.,2009;Moscou &Bogdanove,2009),TALEN 技術由此應運而生。Christian 等(2010)將TALE蛋白與I核酸內切酶融合構成人工TALEN 核酸酶,一系列TALEN 蛋白串聯起來可組成DNA 識別域,每一個TALEN 蛋白能識別并結合一段14~20 bp 的堿基序列,I 核酸內切酶形成二聚體時可以切割雙鏈DNA。TALEN 技術是一種理論上能夠對任意DNA目的序列進行靶向遺傳修飾的基因編輯技術,是實現基因敲除、基因敲入或轉錄激活等靶向基因組編輯的里程碑。
目前TALEN 技術已經成功應用于酵母、哺乳動物和植物的位點特異性基因打靶,與鋅指核酸酶系統相比有較大的應用優勢,設計更簡單、特異性更高,但仍然有脫靶效應、TALEN 與靶序列的特異性結合受到染色體位置及鄰近序列影響等問題存在(李想 等,2017)。
1.4 成簇規律性間隔短回文重復系統
CRISPR/Cas 系統是繼ZFN 和TANEN 技術之后最新發現的第三代基因編輯技術,更簡單有效,應用前景更廣闊。
CRISPR 是細菌和古細菌基因末端的多組DNA序列與反向序列以及30 bp 左右的空格序列(spacer DNA)串聯成簇的、間隔規律的短回文重復序列。該結構最早是在Ishion 等(1987)對大腸桿菌的堿性磷酸酶基因研究中被發現。隨后,Jansen等(2002)進一步研究了原核生物中的該重復DNA 序列,將該結構命名為CRISPR,同時確定了4 個CRISPR相關基因(-,)。在不同物種中根據Cas 蛋白不同將CRISPR 系統分為3 大類、10 小類,其中依賴Cas9 蛋白的CRISPR 系統屬于第2 類,最簡單常用(Lino et al.,2018)。CRISPR/Cas9 系統由Cas9 核酸內切酶、具有靶向特異性的CRISPR RNA(crRNA)和反式激活CRISPR RNA(trans-activating crRNA,tracrRNA)3 種 元件組成。crRNA 通過堿基配對與tracrRNA 結合形成tracrRNA/crRNA 復合物,通過人工設計這兩種RNA,可以將其改造、整合形成具有引導作用的sgRNA(single guide RNA),能引導Cas9 蛋白對DNA 的定點切割(Jinek et al.,2012)。sgRNA的特異識別功能依賴靶序列下游3′末端的一段保守結構—PAM(proto-spacer adjacent motifs),tracrRNA/crRNA 復合物在PAM 序列上游切割DNA造成DSB,并啟動DNA 損傷修復機制。
CRISPR/Cas9 系統設計簡單,在多基因編輯應用方面更簡單高效,僅需設計sgRNA,就能同時進行多基因編輯,并且靶向精確、脫靶率低、細胞毒性低、更廉價簡便,深受研究人員的青睞。
2 基因編輯技術在蔬菜中的研究應用
目前,基因編輯技術已經在植物的遺傳基礎理論研究和生產實踐中廣泛應用,植物基因的靶向修飾是基因編輯技術應用最廣泛的領域。基因編輯技術可以通過修飾內源基因來幫助設計所需的植物性狀,在模式植物和大田作物中都有大量報道;蔬菜作物種類繁多,基礎研究較大田作物稍落后,主要在茄科、十字花科及葫蘆科這三大科常見蔬菜中研究較多。本文對相關文獻進行匯總,介紹了基因編輯技術在以上三大科主要蔬菜作物中的研究概況(表1)。
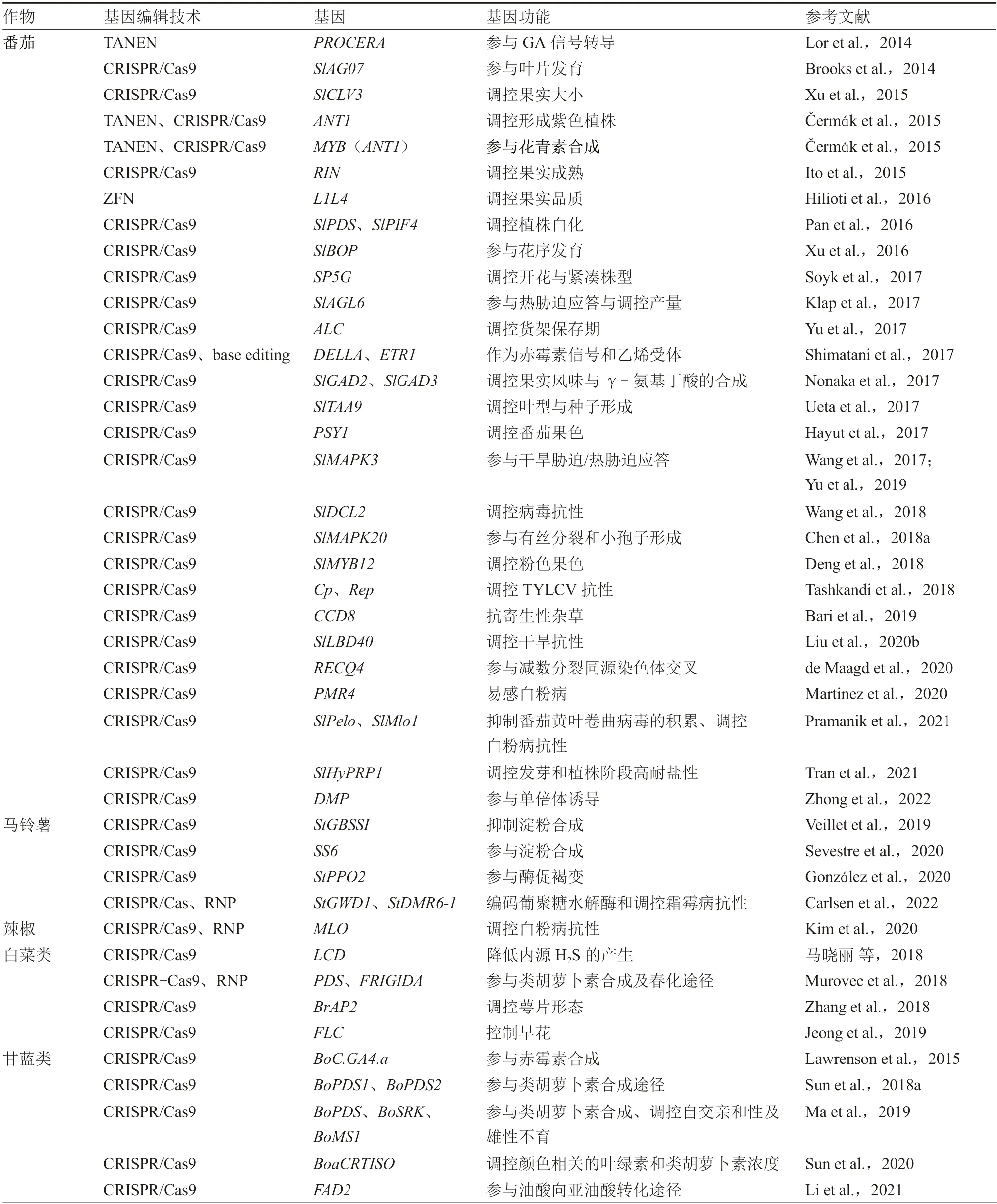
表1 主要蔬菜作物中的基因編輯研究

續表
2.1 茄科蔬菜中基因編輯技術的研究應用
茄科蔬菜的基因編輯技術主要在番茄、馬鈴薯、辣椒上報道較多。
2.1.1 番茄中基因編輯技術的研究 番茄()屬于茄科番茄屬二倍體作物,在全世界廣泛栽培,其基因組已經完成測序,遺傳轉化方法也比較成熟,是理想的蔬菜基因編輯技術研究材料。目前在番茄中已有大量基因編輯相關文獻報道(張愛萍 等,2022)。
Hilioti 等(2016)首次證實了基于ZFNs 的基因編輯技術在番茄上的有效性,通過對番茄--()基因不同位點的編輯改造,證實了的DNA 結合域上游序列的改變可導致包括番茄果實在內的多種表型變異。
Lor 等(2014)首次在番茄基因編輯中成功使用TALEN 技術,該研究的靶基因()是番茄中唯一報道的DELLA 家族基因,而DELLA 蛋白是赤霉素(GA)信號的負調控因子。該研究通過基因編輯產生錯義突變導致部分活性喪失,從而確定在番茄GA 信號轉導中的作用。
Brooks 等(2014)首次利用靶向基因(),將CRISPR/Cas9技術應用于番茄基因編輯,發現敲除的番茄突變體植株葉片呈針狀而不是正常的扁平復葉。?ermák等(2015)比較了TALENs 和CRISPR/Cas9 在番茄上的基因編輯效率,該研究在控制花青素生物合成的基因上游插入一個強啟動子,導致花青素在番茄組織中過表達和異位積累。在這次基因編輯中,超過2/3 的插入是精確的,并且沒有檢測到脫靶修飾,TALENs 和CRISPR/Cas9 都以相似的效率實現了基因靶向編輯。
隨著基因編輯技術的發展,CRISPR/Cas9技術逐漸取代了ZFN和TALEN技術。Danilo 等(2018)以番茄花色苷合成基因為靶基因,利用CRISPR/Cas9 系統在番茄上實現了同源介導(homology directed repair,HDR)的外源基因插入。Li 等(2018a)通過CRISPR/Cas9 系統在番茄中進行多基因編輯,該研究同時敲除了類胡蘿卜素代謝途徑中的多個基因,抑制了番茄紅素向α-胡蘿卜素和β-胡蘿卜素的轉化,促進了番茄紅素的生物合成和積累。隨后,以CRISPR/Cas9 為主的基因編輯技術就在番茄上廣泛應用,在產量、抗病性、非生物脅迫等基因挖掘到性狀改良上都取得了快速進展(Shimatani et al.,2019;Pramanik et al.,2021;Tran et al.,2021)。最近,根據玉米和擬南芥上的研究基礎,Zhong 等(2022)通過CRISPR/Cas9 基因編輯技術改造()基因,首次在番茄上創制了單倍體誘導系,揭開了蔬菜倍性育種的新篇章。
2.1.2 馬鈴薯中基因編輯技術的研究 馬鈴薯(L.)是世界上重要的主食作物之一,屬于茄科茄屬的雜合四倍體作物。目前利用CRISPR/Cas9 技術已經成功在馬鈴薯中實現了定向誘變。
Veillet 等(2019)采用CRISPR/Cas9 技術成功編輯了馬鈴薯的顆粒結合型淀粉合成酶(granulebound starch synthase,GBSS)基因,使淀粉合成受阻。Sevestre 等(2020)利用CRISPR/Cas9 技術敲除了1 個新發現的馬鈴薯淀粉合成酶基因,其結果顯示:在基因第二外顯子末端有一個保守序列(GT1),該序列作為靶點在CRISPER/Cas9 介導的基因編輯中表現出較高的基因編輯效率。
多酚氧化酶類(polyphenol oxidases,PPOs)是一種含銅酶,廣泛分布于高等植物中,能夠催化多種酚類化合物氧化成各自的醌類物質。醌類物質可以進行自聚合,與蛋白質中的氨基酸或自由基反應,形成深色沉淀物,這一過程即為“酶促褐變”,是水果和蔬菜品質降低的原因。González 等(2020)通過CRISPR/Cas9 系統敲除基因,顯著降低了馬鈴薯塊莖的PPOs 活性,褐變程度也明顯減輕。
馬鈴薯是一種高度雜合的四倍體作物,其品種內和品種間基因組呈現高度雜合性和多態性,也是基因編輯策略和設計中要重點考慮的技術難點。Carlsen 等(2022)發現在原生質體這類細胞池水平上的高編輯效率,對在四倍體中實現全等位基因敲除至關重要,該研究通過聚乙二醇(PEG)介導、CRISPR/Cas 核糖核蛋白顆粒(ribonucleoprotein complex,RNP)瞬時轉化原生質體,獲得編碼葡聚糖水二激酶(GWD)和霜霉病抗性6(-)基因的編輯植株。高編輯效率減少了下游繁瑣而復雜的離體再生程序,而且用兩個或多個RNP 同時靶向兩個靶區中的任意一個,均對基因編輯產生明顯的正協同效應。這些初步發現可能會引發更大規模的研究,以促進和優化植物的精確育種。
2.1.3 辣椒中基因編輯技術的研究 辣椒(L.)屬于茄科辣椒屬,是亞熱帶和溫帶國家最廣泛栽培的茄科物種。辣椒于2014年完成基因組測序,為其基因編輯技術研究奠定了基礎。
Li 等(2020)基于辣椒Zunla-1 參考基因組完成了辣椒全基因組的編輯位點鑒定和特異性分析,確定了辣椒基因組的1 組高度特異的CRISPR/Cas9編輯位點。該套編輯位點具有廣泛的適用性,有助于將辣椒中CRISPR/Cas9 的脫靶率降至最低,有效應對CRISPR/Cas9 系統的脫靶效應。
辣椒的遺傳轉化受組織培養條件的限制,不同基因型組織培養能力差距大。Woo 等(2015)發現Cas9/sgRNA 核糖核蛋白復合物RNP 直接應用于原生質體轉化可以獲得無外源基因插入的編輯植株,減少了前期組織培養步驟,也是辣椒基因編輯技術研究的適宜方向。Kim 等(2020)利用來源于葉片和愈傷組織的辣椒原生質體做受體,進行RNP 轉染,在辣椒CM334 和甜椒Dempsey 中實現了對植物病原尤其是白粉病病原具有廣譜抗性的目標基因的編輯。該研究為在一定程度上突破組織培養的限制、實現RNP 途徑的基因編輯技術在辣椒上的應用奠定了基礎。
盡管目前已有辣椒全基因組序列和基因組編輯工具,但由于缺乏穩定的辣椒轉化體系,致使辣椒的精確基因編輯仍處于起步階段。為此,Park 等(2021)采用3 種農桿菌菌株AGL1、EHA101、GV3101,選擇2 個辣椒品種辣椒CM334、甜椒Dempsey,選用pBAtC 作為CRISPR/Cas9 基因編輯載體,優化了篩選劑磷化氫蓖麻毒素(PPT)濃度,結果顯示:GV3101 在甜椒Dempsey 中得到了更好的轉化編輯效果。
2.2 十字花科蔬菜中基因編輯技術的研究應用
十字花科蔬菜種植最為廣泛的兩個屬分別是蕓薹屬()和蘿卜屬(),其中蘿卜屬的基因編輯研究鮮有報道。蕓薹屬包括多種重要蔬菜作物,主要有3 個二倍體基本種,即白菜(,2=20,AA)、甘藍(,=18,CC)、黑芥(,2=16,BB),以及三者之間相互雜交、經過不斷選擇進化得到的3 個異源四倍體復合種,即甘藍型油菜(,2=38,AACC)、芥菜型油菜(,2=36,AABB)和埃俄比亞芥(,2=34,BBCC)。目前基因編輯研究報道較多的是在白菜、甘藍和甘藍型油菜上。
2.2.1 白菜類蔬菜中基因編輯技術的研究 白菜類蔬菜包括7 個變種,常見的有大白菜(L.ssp.)、普通白菜(L.ssp.Makino)、菜薹(L.var.Tsen et Lee)等。近年來,隨著基因編輯技術的發展,白菜基因組和育種研究都取得了快速的發展。
大白菜作為一種重要的大宗蔬菜,種植范圍廣泛、歷史悠久。生產者最初選擇了開花晚的大白菜品種來提高產量,但是晚開花的大白菜需要經過冷處理數周才能誘導開花,最終收獲種子。這種晚熟春化誘導開花的特性延長了大白菜的育種年限,阻礙了大白菜相關遺傳研究的快速開展。為了解決這一問題,Jeong 等(2019)利用CRISPR/Cas9 系統將目標突變引入()基因,培育出不需要春化即可早開花的大白菜(和的雙敲除突變體),加快了繁育進程。同時,該研究還發現和之間存在功能冗余。
硫化氫(hydrogen sulfide,HS)是植物細胞內源信號分子,對植物的生長、發育和抗逆性具有重要的調節作用,而L-半胱氨酸脫巰基酶(L-cysteinedesulfydrase,LCD)是合成內源HS的關鍵酶(尚玉婷 等,2018)。馬曉麗等(2018)采用CRISPR/Cas9 基因編輯技術對大白菜基因進行敲除,基因編輯植株中內源HS 含量有不同程度的降低,為研究HS 信號在大白菜中的生理作用及育種應用提供了理論基礎。
姜明亮(2020)選取孤基因(orphan genes)()及其高度同源基因()為靶基因,在大白菜中創制高效靶向突變,降低了果糖、葡萄糖和可溶性糖的含量,但蔗糖含量和蔗糖合成酶活性明顯增加,推測可能依賴蔗糖合成途徑影響可溶性糖代謝。該研究為白菜功能基因組的研究提供了重要技術支撐,同時對深入研究孤基因影響可溶性糖代謝的作用機制提供了材料和理論參考。
菜薹又稱菜心,為不結球白菜類型中以幼嫩花莖為主要食用器官的常規栽培種,雖然在食用廣度和栽培面積上不如大白菜,但其生育周期短,播種至產品收獲僅需40~60 d,是研究白菜類蔬菜轉化技術的理想材料。國家蔬菜工程技術研究中心生物技術室在菜薹原位轉化成功的基礎上(Liu et al.,1998),結合CRISPR/Cas9 系統,以四九菜心為試材,番茄紅素脫氫酶基因()為靶基因,采用真空滲透原位轉化方法,獲得了無外源片段插入的基因編輯植株(宗梅 等,2022)。
2.2.2 甘藍類蔬菜中基因編輯技術的研究 甘藍類蔬菜為十字花科蕓薹屬的一年生或兩年生草本植物,包括多個變種,常見的有結球甘藍(var.L.)、羽衣甘藍(var.DC.)、花椰菜(.var.L.)和青花菜(L.var.Plenck)等。甘藍自交不親和性強,必須通過低溫誘導開花,生育期長,變種內遺傳基礎狹窄,基因編輯技術對高效創制甘藍的遺傳突變、實現優良品種改良起到重要作用。
Ma 等(2019)設計了1 種基于內源tRNA 處理的CRISPR/Cas9 基因編輯系統,實現了對結球甘藍的多位點和多基因突變。該研究以番茄紅素脫氫酶基因、S-位點受體激酶基因和雄性不育相關基因為靶基因,采用串聯tRNA-sgRNA 結構的載體表達多個sgRNA,結果顯示:基因敲除的植株表現白化矮小,作為可視標記直接驗證該基因編輯系統的可行性;基因突變完全抑制了結球甘藍的自交不親和性,將自交不親和系轉化為自交親和系;而基因突變產生了1 個雄性不育突變體,由于同時發生基因突變,該突變體在開花期與其非突變等位基因雜交兼容,從而使該雄性不育系能夠通過蜜蜂介導的異花授粉進行繁育。
Sun 等(2018a)首次報道了CRISPR/Cas9 技術在羽衣甘藍上的應用,該研究以類胡蘿卜素合成相關的基因為靶點,使羽衣甘藍兩個同源基因同時發生定點突變,為羽衣甘藍基因功能研究和分子育種提供了技術支撐。該課題組在此技術基礎上,對羽衣甘藍類胡蘿卜素異構酶基因()進行編輯,共獲得13個突變體,突變率高達81.25%;雙等位基因突變株和純合突變株的總類胡蘿卜素和葉綠素含量均降低,總水平下降了11.89%~36.33%,葉片顏色從綠色變為黃色,創制了黃色羽衣甘藍新種質(Sun et al.,2020)。
單堿基編輯技術(base editing)是在CRISPR/Cas9 系統基礎上添加一些酶,實現對DNA 的單個堿基進行替換,具有高效而又精確的基因編輯能力,可在動植物細胞內引入點突變,用于培育具有理想表型的基因編輯動植物。例如:Cas9 蛋白與胞嘧啶脫氨酶組成融合蛋白可以實現胞嘧啶(C)到胸腺嘧啶(T)的單堿基轉換,被稱為BE3(APOBEC1-XTEN-nCas9-UGI,第三代堿基編輯器)系統(Tian et al.,2018);腺嘌呤單堿基編輯器(簡稱ABE),可以對目標位點實現高效腺嘌呤(A)到鳥嘌呤(G)的單堿基轉換(Liu et al.,2022)。北京市農林科學院蔬菜研究所生物技術研究室在花椰菜基因編輯技術體系的基礎上,通過BE3 系統對花椰菜的乙酰乳酸合成酶(acetolactate synthase)基因和著絲粒特異組蛋白變體(centromerespecific histone H3 variant)基因進行單堿基編輯突變,突變率為22%~87%,創制了高抗除草劑的花椰菜新種質(Wang et al.,2022)。
2.2.3 甘藍型油菜中基因編輯技術的研究 甘藍型油菜(L.)作為重要的油料作物,基因組和基因編輯相關研究較多,在產量、品質、抗性、花色、育性等多種性狀中都有研究報道(楊文文等,2021)。通過CRISPR/Cas9 系統,Karunarathna等(2020)同時敲除甘藍型油菜中含油量相關的和兩個基因,獲得了種子含油量明顯增加的突變體植株;Khan 等(2021)敲除CYP78A6 家族基因,突變體植株角果和種子變小,但單株種子數量和質量增加;Sriboon等(2020)敲除開花調控基因的5 個拷貝,發現敲除其中1 個拷貝()后,植株出現早花表型,而敲除其他拷貝,則對開花時間沒有明顯影響;Liu 等(2020a)發現,敲除甘藍型油菜中兩個參與類胡蘿卜素合成的玉米黃質環氧化酶基因和,改變了花瓣中類胡蘿卜素的積累,紫黃素含量降低,葉黃素含量升高,從而導致了橘色花的產生;Xin 等(2020)發現,敲除Y127 株系中的Ms5 基因或敲除Westar 株系中的Ms5 基因,均會導致雄性不育,進一步研究顯示,Ms5 通過與Ms5 或Ms5 形成無功能的異源二聚體而顯性抑制Ms5 或Ms5 的功能,從而導致雄性不育。
Wu 等(2020a)利用基于鼠源胞嘧啶脫氨酶的單堿基編輯器BE3,將基因197 位密碼子上的C 轉化為T,獲得的P197S 油菜突變體表現出耐除草劑苯磺隆的表型。Cheng 等(2021)利用基于人源胞嘧啶脫氨酶的單堿基編輯器,突變油菜基因,發現和的P197F(根據擬南芥的ALS 序列編號)突變均能賦予油菜抗苯磺隆除草劑的能力。且這兩個拷貝同時突變的油菜植株比突變單個拷貝的植株能夠耐受更高濃度的除草劑,說明突變介導的除草劑抗性具有一定的劑量效應。
2.3 葫蘆科蔬菜中基因編輯技術的研究應用
葫蘆科是世界上重要的可食用植物科之一,其重要性僅次于禾本科、豆科和茄科,其中包括黃瓜、南瓜、絲瓜、西瓜等常見的蔬菜和瓜果。然而,這些作物中大多數尚未建立高效的遺傳轉化體系。葫蘆科蔬菜中基因編輯的研究報道大多在黃瓜和西瓜上。
2.3.1 黃瓜中基因編輯技術的研究 黃瓜(L.)屬于葫蘆科黃瓜屬的二倍體蔬菜作物,其基因編輯的相關報道大都是利用CRISPR/Cas9技術。
以色列科學家采用CRISPR/Cas9 技術首次在黃瓜中開展抗病毒研究(Chandrasekaran et al.,2016)。該試驗設計了兩個sgRNA,以黃瓜(真核翻譯起始因子4E)基因為靶基因,獲得了一系列靶點缺失和插入突變的植株。用黃瓜葉脈黃化病毒(CVYV)、小西葫蘆黃花葉病毒(ZYMV)和番木瓜環斑型花葉病毒(PRSV-W)接種突變株和野生型黃瓜植株,基因編輯純合突變株表現出廣泛的病毒抗性。
Hu 等(2017)利用CRISPR/Cas9 系統編輯了抑制黃瓜心皮發育的基因,成功獲得了全雌系材料,提高了產量。短瓜基因編碼1 種葫蘆科特異的環型E3 連接酶,其突變可導致自身泛素化和降解增強,同時造成乙烯合成速率限制酶ACS2 的積累,Xin 等(2019)成功通過CRISPR/Cas9 技術獲得了和這2 個基因的編輯突變體,并研究了其調控乙烯合成和黃瓜果實伸長的分子機制。Zhang 等(2020a)成功實現了對短瓜基因的CRISPR/Cas9 編輯,驗證了通過促進細胞增殖調控黃瓜果實伸長。基因屬于TCP(PROLIFERATING CELL FACTORS)基因家族,Yang 等(2020)成功獲得基因的CRISPR/Cas9 突變體,揭示了基因通過直接調控乙烯的合成來控制黃瓜莖卷須的形態和攀緣。
毛狀根誘導系統因其生長快、遺傳穩定性高,可作為研究基因表達和功能的有效方法,也被證實是評估CRISPR/Cas9 系統用于基因組編輯活動的有效工具(Jacobs &Martin,2016)。Nguyen 等(2022)對黃瓜根瘤菌介導的毛狀根轉化系統進行了優化,為驗證植物轉化載體、CRISPR/Cas9 構建活性以及選擇黃瓜基因編輯的靶向gRNA 提供了有效工具,建立的毛狀根轉化系統為黃瓜和其他植物基因組的編輯技術優化和進一步研究提供了重要信息和新的思路。
2.3.2 西瓜中基因編輯技術的研究 西瓜()屬于葫蘆科西瓜屬一年生蔓生藤本植物。西瓜是葫蘆科中第2 個基因組編輯成功的物種(Tian et al.,2017,2018)。
Tian 等(2017)采用CRISPR/Cas9 技術對西瓜的(phytoene desaturase)基因進行精確編輯。該研究獲得的轉化苗幾乎達到了100%的編輯效率,所有的轉基因西瓜植株都帶有S 突變,并表現出明顯的或鑲嵌式的白化表型。隨后,Tian等(2018)利用基于CRISPR/Cas9 的單堿基編輯器特異性突變乙酰乳酸合酶(acetolactate synthase,ALS)基因的Pro190 位點,獲得了抗除草劑苯磺隆的西瓜新種質。Zhang 等(2020c)采用CRISPR/Cas9 系統敲除了編碼植物磺肽素(Phytosulfokine,PSK)前體的基因,以增強西瓜對尖孢鐮刀菌(,FON)的抗性。
雖然西瓜遺傳轉化取得了一些進展,但從組織培養到再生仍然具有挑戰性。Feng 等(2021)根據小麥中的研究結果,通過共表達生長調節因子4(GRF4)和GRF-相互作用因子1(GIF1),改善西瓜再生,在西瓜中實現了高效轉化,并與基因組編輯工具相結合,在西瓜中實現了高效的基因編輯,成功地創制了二倍體無籽西瓜。該研究為今后西瓜遺傳轉化的研究和育種提供了有力的理論基礎。
3 總結與展望
基因編輯技術作為生命科學的顛覆性技術,已在農業領域得到了廣泛應用。CRISPR-Cas 系統的出現加速了基因編輯技術的發展,基因編輯工具的開發和應用日新月異,具有更廣泛的目標、更高的效率和特異性,以及更高的精度。目前,基因編輯技術已被應用于植物中各種可遺傳的基因組修飾,例如:隨機插入/缺失(InDels),點突變或核苷酸替換,同源基因和多基因突變,堿基編輯器(base editor),先導編輯器(prime editor),HDR 介導的基因替換、點突變、DNA 插入和刪除,以及有針對性的染色體重排等(Liu et al.,2022)。在技術的持續拓展及延伸下,基因編輯將在更廣泛的領域帶來更大的社會價值與經濟價值。蔬菜作為人們日常飲食中必不可少的食物之一,是提供人體必需的多種維生素和礦物質的主要來源。蔬菜中的VK、VC、有機硫化物和硝酸鹽對慢性疾病的預防有益,鉀、鎂、膳食纖維、類胡蘿卜素等成分有利于預防心腦血管疾病。針對蔬菜的這些特征定向育種,對一些優良性狀進行聚合和改良,提高蔬菜的產量、抗性、品質和口感一直是育種家和消費者共同追求的目標。傳統育種方法不但易受種間生殖隔離的限制、不良基因連鎖的影響,而且育種年限很長。基因編輯技術優勢明顯,可以克服以上困難,在性狀改良上表現出良好的靶向性和易用性,發展空間巨大,應用前景廣闊。
蔬菜作物種類繁多,基因組學研究基礎差距大,較大田作物在遺傳轉化和基因編輯研究上都相對滯后。但基因編輯工具是相通的,最新的基因編輯工具已經很快應用到蔬菜作物中,如花椰菜和西瓜中的單堿基編輯器(Tian et al.,2018;Wang et al.,2022),番茄和甘藍型油菜上的同源基因和多基因編輯(Zhai et al.,2020;Tran et al.,2021),同源介導基因編輯的外源基因插入(Danilo et al.,2018),以及RNP的應用(Kim et al.,2020),等等。雖然蔬菜基因編輯研究上取得了很多亮點,但在一些蔬菜中還存在缺乏高效遺傳轉化技術的瓶頸問題,如辣椒、西瓜、南瓜、白菜、茄子等,缺乏適用于多種基因型的高效遺傳轉化體系,同時可用于編輯的功能基因研究也是核心問題。
圍繞當前基因編輯技術在蔬菜作物中存在的問題,無轉化體系的蔬菜作物應盡快建立適宜的轉化體系。首先可以根據不同蔬菜作物的特性,研究建立不同外植體、不同轉化途徑的技術體系。例如:番茄以子葉、葉片和下胚軸為外植體,花椰菜以下胚軸和花序軸為外植體都有成功的轉化體系建立(Brooks et al.,2014;Hilioti et al.,2016;Wang et al.,2022)。對一些難以轉化和再生的作物,如辣椒、茄子、西葫蘆等,可探索以大孢子、小孢子、原生質體、花粉粒等為受體的轉化編輯體系。同時不依賴遺傳轉化、直接獲得無外源基因插入的基因編輯材料顯得更具優勢,例如:通過農桿菌介導的基因的瞬時表達(Iaffaldano et al.,2016;Chen et al.,2018b),以及RNP 原生質體侵染等(Woo et al.,2015;Murovec et al.,2018)。本課題組也在蕓薹屬菜薹上探索不依賴組織培養的原位轉化編輯體系,并取得了一些進展(宗梅 等,2022)。因此,建立和完善不同蔬菜作物的遺傳轉化技術,努力探索不依賴于組織培養的轉基因途徑是蔬菜作物基因編輯技術廣泛應用的研究方向。
另外,隨著基因組的大規模測序,蔬菜功能基因組的研究也迅速發展。通過基因組測序和大數據分析,根據大田和模式作物的研究基礎快速高效鑒定蔬菜中對應的功能基因,針對蔬菜作物性狀特點結合正向遺傳和反向遺傳明確農藝性狀的控制基因,再結合基因編輯技術鑒定基因功能和作用機制。可編輯的功能基因研究和基因編輯技術的發展應用是相互促進、相輔相成的。
雖然基因編輯技術在蔬菜上的研究已經取得了大量進展,但在不同種類蔬菜作物上的廣泛應用還有很多問題需要解決。隨著蔬菜基因組研究的逐漸深入和發展,技術的不斷完善和延續,基因編輯技術作為最具潛力的基因組改造工具必將在蔬菜作物功能基因研究和分子育種應用中發揮重大作用。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
