亞胺還原酶的改造及其催化合成1-苯基-1,2,3,4-四氫異喹啉的性能
2022-08-29 10:52:24曹文斌陳飛飛鄭高偉許建和
華東理工大學學報(自然科學版) 2022年4期
曹文斌, 李 昊, 陳飛飛, 潘 江, 鄭高偉, 許建和
(華東理工大學生物工程學院,省部共建上海生物制造產業協同創新中心,上海 200237)
四氫異喹啉(THIQs)作為具有生物活性的分子骨架在天然產物與藥物中存在非常普遍[1-2],常作為受體拮抗劑[3]、受體激動劑[4]、酶抑制劑[5-6]等。其中1-苯基-1,2,3,4-四氫異喹啉(1-Ph-THIQ)由于對神經系統具有獨特的抑制作用引起制藥領域的廣泛關注[7],而(S)-1-苯基-1,2,3,4-四氫異喹啉((S)-1-Ph-THIQ)是膀胱過度活動癥治療藥物索利那新(Solifenacin)的關鍵結構砌塊[8]。近年來,亞胺還原酶(IRED)催化環形亞胺不對稱氫化的反應路線具有反應條件溫和、立體選擇性高的優點,被越來越多地用于不同結構手性胺的不對稱合成[9-10]。然而,由于亞胺底物1-苯基-四氫異喹啉(1-Ph-DHIQ)中苯基的巨大位阻和苯基對鄰近官能團的共軛效應,使其酶法合成具有較大挑戰性。
迄今為止,對1-Ph-DHIQ 具有S對映體選擇性的亞胺還原酶報道僅有2 例[11-12],一是亞胺還原酶SnIR[11],比活力只有13 mU/mg,對映體選擇性(ee值)為51%;而改造所得的突變體T123I/G228T的比活力增加到56 mU/mg,ee值達91% ;二是武漢大學瞿旭東課題組報道的亞胺還原酶IR45[12],其對映體選擇性具有較高ee值(99%),且對底物1-Ph-DHIQ 具有較好的催化性能,但存在明顯的底物抑制。
為獲得高效催化合成(S)-1-Ph-THIQ 的亞胺還原酶,本文首先對前期篩選獲得的亞胺還原酶(IRED)進行反應性能比較,然后對性能較好的候選酶AdIR1 進行分子改造,并對所得的酶突變體進行動力學與熱穩定性表征,最后嘗試利用優質突變酶進行(S)-1-Ph-THIQ 克級制備實驗。
1 實驗部分
1.1 原料和試劑
瓊脂糖凝膠DNA 回收試劑盒,質粒小量提取試劑盒和PCR 產物回收試劑盒購自上海艾德萊生物有限公司;DNA marker、蛋白質marker、2×Taq PCR Mix購自上海康為有限公司;引物由上海生工生物工程技術有限公司代合成;Ni 柱吸附親和層析柱購自GE Healthcare 公司;蛋白胨和酵母膏購自Oxoid 公司;限制性內切酶購自New England Biolabs有限公司;PrimeSTAR 高保真酶和T4 DNA 連接酶購自Takara有限公司;其余化學試劑均為國產分析純。
PCR 儀、電泳儀、凝膠成像儀、紫外切膠儀、蛋白電泳儀、酶標儀,美國Bio-Rad 公司;超微分光光度計,Nanodrop 2000c 型,美國Thermo Fisher 公司;全自動高壓滅菌鍋,日本TOMY 公司;高速離心機,日本HITACHI 公司;紫外-可見分光光度計,日本SHIMADZU 公司;超凈工作臺,蘇州凈化設備有限公司;恒溫培養箱,上海一恒科技有限公司;恒溫震蕩搖床、深孔板搖床,上海智誠科技有限公司;超聲破碎儀,寧波新芝生物科技有限公司。
1.2 目的基因克隆
本文使用表1 中所示引物通過PCR 分別從菌株Saccharothrix espanaensisDSM 44229 和Amycolatopsis decaplaninaDSM 44594 的基因組中擴增獲得亞胺還原酶目的基因AdIR1和SeIR2。
PCR 體系(20 μL):基因組模板1 μL (0.2 μg),上下游引物分別加入1 μL(0.5 μmol/L),2 × Taq Mix 10 μL,和ddH2O 7 μL。PCR 擴增步驟:(1) 95 ℃,預變性3 min;(2) 95 ℃,變性10 s;(3) 55 ℃,退火5 s;(4) 72 ℃,延伸1 min,然后步驟(2)~(4)循環30 次;(5) 72 ℃延伸,延伸10 min,冷卻至4 ℃保存。
使用φ=1%的瓊脂糖凝膠對PCR 產物進行電泳分析,采用瓊脂糖凝膠DNA 回收試劑盒回收目的基因片段。目的基因和pET-28a(+)質粒經限制性內切酶NdeI 和EcoR I 雙酶切、T4 DNA 連接酶連接構建重組質粒,并轉化至Escherichia. coli(E.coli) BL21 (DE3)感受態細胞。
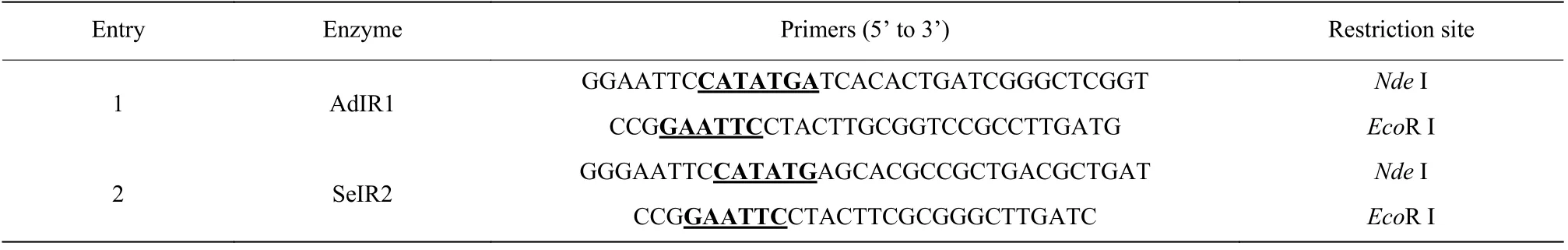
表1 目的基因克隆所用的引物Table 1 Primers used in target gene cloning
1.3 重組菌誘導與表達
從重組菌E. coliBL21 (DE3)甘油保藏管中取樣10 μL 接種至4 mL 的Lysogeny Broth (LB)培養基中(含50 μg/mL卡那霉素),于37 ℃、200 r/min 條件下培養過夜。從菌液試管中轉接1 mL 菌液至含100 mL LB 培養基(含50 μg/mL 卡那霉素)的搖瓶中放大培養,當搖床(37 ℃,220 r/min)培養至吸光度OD 值(OD600)為0.6~0.8 時,加入異丙基-β-D-硫代半乳糖苷(IPTG,終濃度為0.2 mmol/L)于16 ℃、200 r/min條件下誘導表達24 h。最后在4 ℃、7 000g下離心收集菌體,對收集的菌體進行超聲破碎獲得細胞破碎液。
1.4 1-苯基-3,4-二氫異喹啉的不對稱還原反應
1 mL 反應體系包含1 mmol/L 1-苯基-3,4-二氫異喹啉、0.1 mmol/L 還原型輔酶Ⅱ(NADPH)、5 mmol/L葡萄糖、1 mg/mL葡萄糖脫氫酶BmGDH 凍干酶粉[13]、100 mmol/L KPB 緩沖液(pH 7.0)和200 μL 新鮮細胞破碎上清(上清液按1 g 細胞配比10 mL 緩沖液制備)。不對稱還原反應如圖1 所示,其中煙酰胺腺嘌呤二核苷酸(NADP+)是還原型煙酰胺腺嘌呤二核苷酸磷酸(NADPH)的氧化形式,GDH 為葡萄糖脫氫酶。
反應條件為30 ℃、200 r/min,每組反應至少重復2 次。采用2 mol/L NaOH 淬滅反應并調節pH 至13,隨后立即用乙酸乙酯進行萃取,并用高效液相色譜分析反應轉化率和產物ee值。1-苯基-1,2,3,4-四氫異喹啉經柱前乙酰化后的高效液相色譜法(HPLC)分析,分析條件為:OD-H 柱(250 mm × 4.6 mm, 5 μm),正己烷和異丙醇體積比為95∶5,流速0.8 mL/min,波長220 nm,溫度30 ℃。(R)-產物出峰時間為20~21 min;(S)-產物出峰時間為22~23 min。
1.5 亞胺還原酶的序列與結構比對
通過CLUSTAL X 程序進行序列比對。通過Swiss-model 在線同源建模[14](以PDB: 4OQY 的蛋白結構作為模板)分別構建得到SnIR 與AdIR1 (與4OQY 的序列一致性分別為43%和46%)的結構模型,再利用Pymol 軟件對SnIR 與AdIR1 進行空間結構比對。

圖1 亞胺還原酶催化的不對稱還原反應Fig. 1 Asymmetric reduction reaction catalyzed by IRED
1.6 AdIR1 酶突變庫的構建與初篩
以母本AdIR1 重組pET-28a(+)質粒為模板,使用PrimeSTAR 高保真聚合酶和表2 所示的目的基因定點突變所用PCR 引物進行PCR 構建單點飽和突變庫。PCR 產物經DpnI 酶消化(37 ℃、2 h)后轉化至E. coliBL21 感受態細胞,并涂布LB 平板(含50 μg/mL 卡那霉素),于37 ℃培養箱過夜培養。
突變體庫篩選步驟如下:
(1) 用牙簽將平板上的突變體轉化子轉移至一級板(單孔有300 μL LB 培養基,含50 μg/mL 卡那霉素)中,于37 ℃、220 r/min 下振蕩過夜培養,每塊板中含有3 個母本和34 個轉化子。
(2) 取50 μL 一級種子液轉接至二級板(單孔有600 μL LB 培養基,含50 μg/mL 卡那霉素)中培養,37 ℃下培養3 h,加入異丙基-β-D-硫代半乳糖苷(IPTG,終濃度0.2 mmol/L),并于16 ℃下繼續培養24 h 誘導突變體表達。
(3) 在3 500 r/min 條件下離心10 min 以收集菌體。
(4) 每孔加入200 μL 裂解液(100 mmol/L KPB 緩沖液,pH 7.0,含750 mg/L 溶菌酶和10 mg/L DNase酶),振蕩使細胞懸浮均勻,然后于37 ℃、200 r/min條件下保溫2 h 破碎細胞。
(5) 在4 ℃條件下,3 500 r/min 離心30 min 后取上清用酶標儀在酶標板中測定活力。
篩選反應條件(200 μL):篩選反應液150 μL (含NADPH 0.27 mmol/L,底物1-Ph-DHIQ 0.13 mmol/L,100 mmol/L KPB 緩 沖 液,pH 7.0),離 心 破 碎 液 上清50 μL。
1.7 AdIR1 突變體的復篩
采用1.3 節所述方法對目標突變體進行發酵培養,使用鎳離子親和吸附柱(His Trap Ni-NTA FF column, 5 mL)對發酵表達的含有His-tag 的重組酶蛋白進行純化[13],并對純酶的比活力進行測定。
1.8 純酶測活與蛋白濃度測定
亞胺還原酶的標準酶活測定體系(1 mL):底物為1-苯基-3,4-二氫異喹啉的二甲基亞砜(DMSO)溶液(10 μL,底物終濃度為1 mmol/L)、NADPH (10 μL,終濃度為0.1 mmol/L),一定濃度的酶液(10 μL)及970 μL KPB 緩沖液(100 mmol/L,pH 7.0)。在30 ℃條件下檢測340 nm 處吸光度的變化。
酶活力(U)單位的定義為:在上述條件下,每分鐘氧化1 μmol 輔酶所需的酶量,酶活計算公式如下所示:
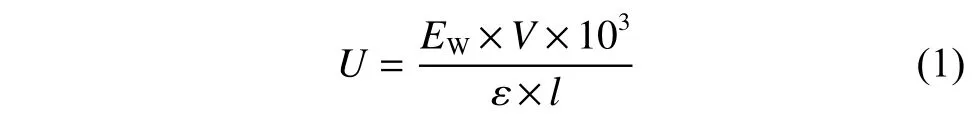
其中:EW代表340 nm 處吸光度在1 min 內的變化值;V代表反應液體系的體積量(mL);ε代表消光系數(L/(mol·cm));l代表光程距離(cm)。
蛋白質含量測定方法:使用Nanodrop 2000c 超微量分光光度計對純酶蛋白含量進行測定,所得測量值除以蛋白的消光系數即得實際蛋白濃度。
1.9 突變體的酶學性質表征
最適催化pH 研究條件:測定亞胺還原酶在pH 3.0~11.0 范圍內不同緩沖液的酶活性,其中pH 3.0~6.0選用檸檬酸-檸檬酸鈉緩沖液(100 mmol/L),pH 6.0~8.0 選用KPB 緩沖液(100 mmol/L),pH 8.0~9.0 選用Tris-HCl 緩沖液(100 mmol/L),pH 9.0~11.0 選用甘氨酸-氫氧化鈉緩沖液(100 mmol/L)。
熱穩定性研究條件:對酶催化活力的半衰期(t1/2)考察,預先用KPB 緩沖液(100 mmol/L, pH 7.0)將AdIR1 突變體酶溶液稀釋至1 mg/mL 后,置于不同的溫度(30、40、50 ℃)下保溫,間隔取樣,按標準測活條件測定酶的殘余活力。以初始活力為基準,將測定得到酶的殘余活力換算為相對活力,相對活力數據通過Excel 線性處理并擬合得到t1/2。

表2 目的基因定點突變所用PCR 引物Table 2 PCR primers used for site-directed mutations of target gene
催化動力學研究條件:測定不同濃度底物(1-甲基-3,4-二氫異喹啉)、不同輔酶(NADPH)濃度下酶的比活力數據,并采用Origin 9.0 軟件根據米氏方程模型擬合獲得酶的動力學參數,分為以下3 種:
(1)無底物抑制條件下的催化模型
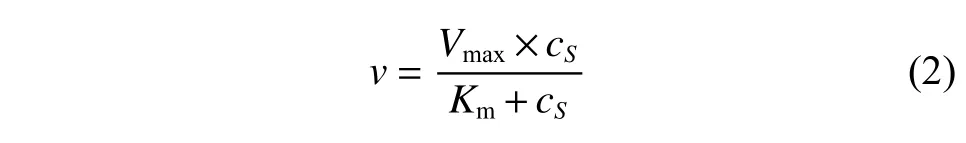
其中:v為酶的比活(μmol/(min·mg));cS為底物或者輔酶濃度(mmol/L);Vmax為催化最大反應速度(μmol/(min·mg)),是底物飽和時的反應速度;Km為米氏常數(mmol/L),是酶達到1/2Vmax時的底物濃度。
(2)底物抑制條件下的催化模型
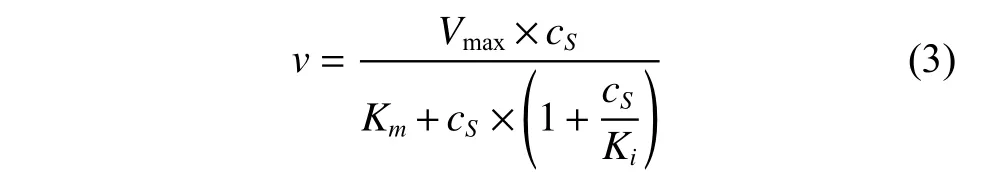
其中:Ki為抑制常數(mmol/L),反映出抑制劑與蛋白質的結合緊密程度。
(3)催化常數kcat計算方程
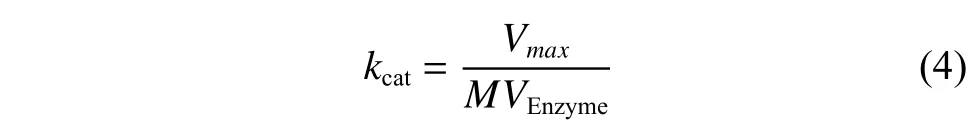
其中:kcat為催化速率(s-1),是一個酶(或者一個活性部位)在底物飽和狀態下反應快慢的參數;MVEnzyme為單位質量酶的分子數量(mmol/mg)。
1.10 1-苯基-1,2,3,4-四氫異喹啉的克級制備
1.10.1 最適反應溫度考察 1 mL 反應體系包含5 mmol/L 1-苯基-3,4-二氫異喹啉、0.1 mmol/L NADPH、25 mmol/L 葡萄糖、1 mg/mL BmGDH 凍干酶粉、5 mg/mL 亞胺還原酶凍干酶粉、100 mmol/L KPB 緩沖液(pH 7.0)和 φ =1%的DMSO。反應條件為轉速200 r/min,溫度分別為30、40 ℃。
1.10.2 最適助溶劑體積分數考察 10 mL 反應體系包含5 mmol/L 1-苯基-3,4-二氫異喹啉、0.1 mmol/L NADPH、25 mmol/L葡萄糖、1 mg/mL BmGDH 凍干酶粉、5 mg/mL 亞胺還原酶凍干酶粉、100 mmol/L KPB 緩沖液(pH 7.0),反應條件為轉速200 r/min,40 ℃,助溶劑體積分數分別為1%、2%、5%及10% 。
1.10.3 克級制備反應 在以上篩選得到的最適溫度以及最適助溶劑濃度條件下,以8 g/L 的突變體凍干酶粉為催化劑,其余反應條件參照1.4 節按比例擴大至2 L 反應體系下進行1-苯基-1,2,3,4-四氫異喹啉的克級制備反應。反應過程定期取樣檢測反應進程。反應結束后進行淬滅、萃取通過硅膠柱層析法(流動相:乙酸乙酯和石油醚體積比為2∶1)對目標產物進行純化。
2 結果與討論
2.1 1-苯基-3,4-二氫異喹啉的不對稱酶促還原反應
通過以上反應條件,本文探索了AdIR1 和SeIR2分別對1-苯基-3,4-二氫異喹啉的不對稱還原反應效果,結果如表3 所示。可以看出,AdIR1 是一個具有較優催化效率的亞胺還原酶,經過24 h 的反應,實現了98%的轉化率(C),合成S構型產物,ee值達98%,結果明顯優于SeIR2。因此,后續以反應效果更優質的AdIR1 作為分子改造的起始酶。

表3 亞胺還原酶不對稱還原1-苯基-3,4-二氫異喹啉的反應Table 3 Asymmetric reduction of 1-phenyl-3,4-dihydroisoquinoline by IRED
2.2 AdIR1 與SnIR 的序列與結構比對
在前期研究中發現,T123I、F178L、G228A 3 個位點的突變會顯著影響SnIR 酶催化1-芳基二氫異喹啉不對稱催化產物的ee值,同時也導致了活力的下降。因此,文中將SnIR 與AdIR1 進行序列及結構比對,以期找到與SnIR 的T123、F178 和G228 位點相對應的AdIR1 關鍵殘基,結果如表4 以及圖2 所示。

表4 亞胺還原酶的關鍵位點比對結果Table 4 Alignment results of key residues of the imine reductases
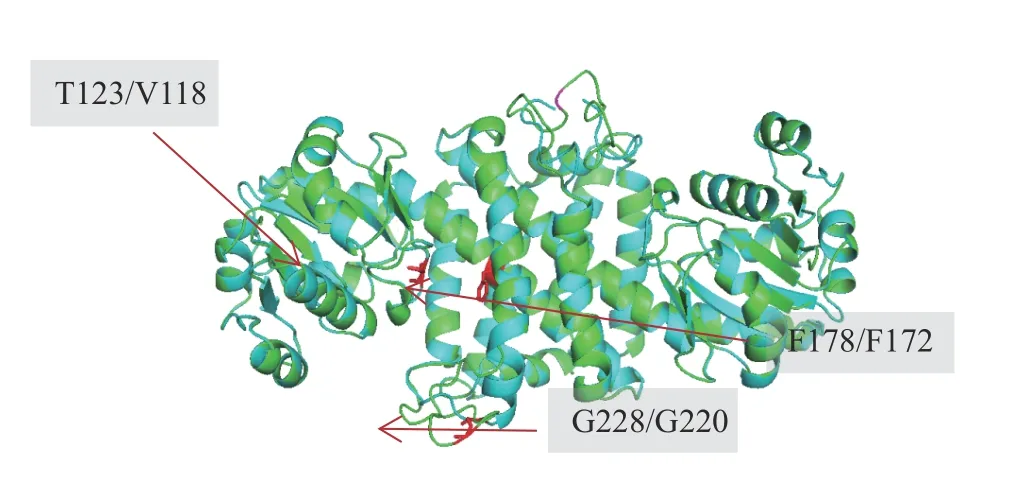
圖2 SnIR 與AdIR1 的結構比對結果Fig. 2 Results for structural alignment of AdIR1 with SnIR
以上比對結果表明,對于V118 與F172 位點,AdIR1 與SnIR 通過序列比對的定位結果與空間結構比對的結果一致。由于這兩個位點都位于序列保守區域,故可信度比較高。而另一方面對于G228 位點,一級序列的比對結果呈現出與空間比對不一致的結果。由于酶分子的半理性改造在原理分析上通常偏好基于空間比對的結果,故最終選擇了G220 作為改造位點。
2.3 AdIR1 的分子改造結果
通過分子改造,雖然在G220 殘基的突變中并未發現酶活有效提升的突變體,但在V118 和F172 位點的單突變嘗試中篩選到幾個活力可能提升的突變體。通過對其宿主細胞進行培養、誘導表達以及親和純化,獲得純酶。隨后,在30 ℃條件下測定純化后的突變酶(M1~M4)對1-苯基-3,4-二氫異喹啉(底物濃度為0.1 mmol/L)活力,結果如表5 所示。

表5 AdIR1 突變體的測活結果Table 5 Activity assay results of AdIR1 mutants
通過測活發現,突變酶M1 與M3 并沒有有效地增加AdIR1 對底物的催化活力,甚至在M2 中,分子改造使得其活力下降。而M4 突變有效增加了酶活,提升了1.9 倍。因此,通過對F172 位點的改造,本文得到了使AdIR1 活力顯著提升的突變體。隨后通過基因測序發現該突變型為F172Y。
2.4 亞胺還原酶突變體的酶學性質表征
2.4.1 pH 對酶活力的影響 通過使用不同的緩沖液,得到了在不同pH 條件下母本(WT)與突變體F172Y的催化活力特征曲線,結果如圖3 所示。與母本相似,突變體F172Y 對pH 呈現出典型的鐘形曲線,且最適pH 同樣為 7.0。
2.4.2 熱穩定性表征結果 對母本和突變體F172Y稀釋酶液在30、40 ℃和50 ℃條件下保溫一定時間后取樣,用底物1-苯基-3,4-二氫異喹啉測定酶活,得到其在不同溫度下殘余活性的變化曲線,結果如圖4所示。半衰期結果由Excel 進行曲線模擬所得,結果如表6 所示。
結果表明,突變體F172Y 雖然僅突變了一個位點,但是熱穩定性有顯著提升,其在30 ℃和40 ℃條件下96 h 保溫后酶活力損失低于20%,并且擬合得到的t1/2在30、40 ℃以及50 ℃條件下分別是母本的4.6 倍、6.3 倍和3.9 倍。其中原母本在40 ℃條件下保溫12 h 殘留活性不足70%,而突變體F172Y 卻保持了91%的殘留活性,與30 ℃條件92%的殘余活性基本保持一致。因此后期的反應進程表征以及制備反應將選用40 ℃作為反應條件。

圖3 pH 對母本(a)與F172Y(b)酶活的影響Fig. 3 Effect of pH on the enzyme activity of WT(a) and F172Y(b)

圖4 母本(a)與F172Y(b)的熱穩定性Fig. 4 Thermostability of WT (a) and F172Y (b)
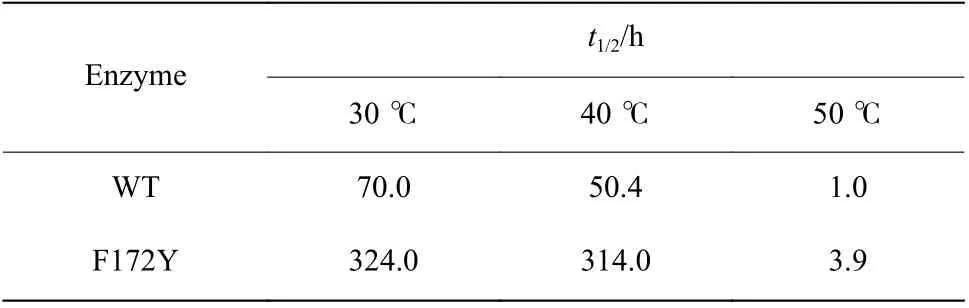
表6 F172Y 與母本的半衰期Table 6 Half-lives of WT and F172Y at different temperatures
2.4.3 動力學表征結果 利用不同濃度的1-苯基-3,4-二氫異喹啉為底物,本文分別測定了母本與突變體F172Y 的底物動力學特征曲線,結果如圖5 所示,并依據動力學特征曲線擬合計算出動力學參數,結果如表7 所示。
通過以上實驗結果表明,WT 與F172Y 對底物1-苯基-3,4-二氫異喹啉皆不存在底物抑制,意味著其在未來的工業生產上具有非常優質的應用前景。此前,通過F172Y 單點的突變,使該酶在Km保持不變的情況下,kcat有了3.4 倍的提升,即催化效率常數(kcat/Km)提升了3.4 倍。而溫度提高時催化效率常數提升不明顯,主要因為kcat有顯著提升的同時Km亦有提升。活力測定表明,在40 ℃且飽和的底物濃度條件下,比活可達46 mU/mg,這比30 ℃條件下的活力(15.3 mU/mg)提高了約3 倍。
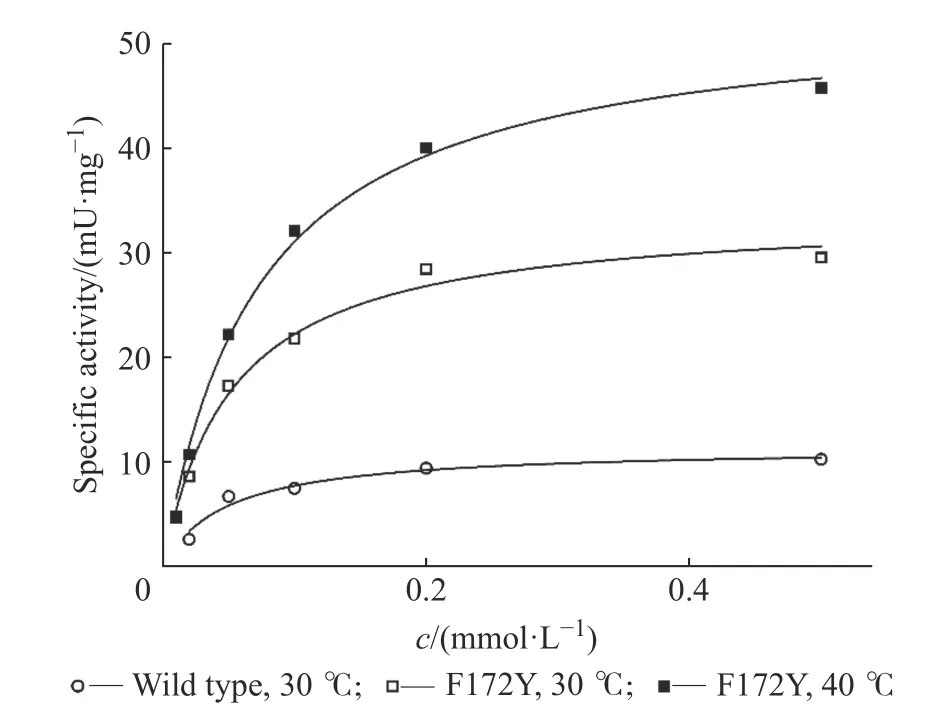
圖5 母本與F172Y 的動力學特征曲線Fig. 5 Kinetic characteristic curves of WT and F172Y
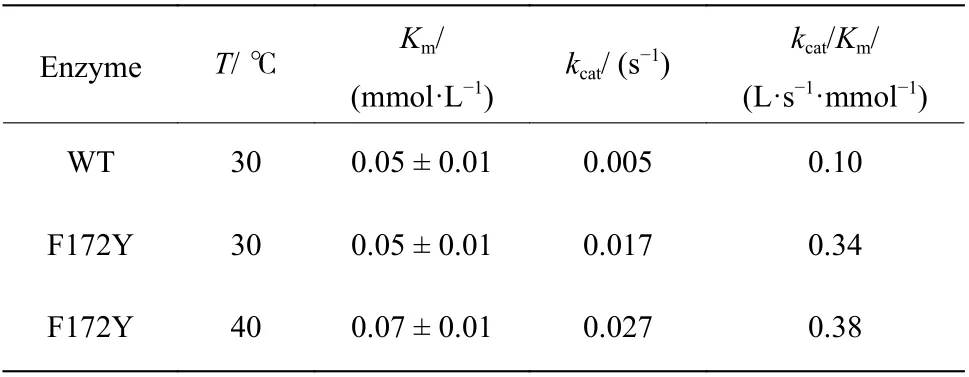
表7 母本與F172Y 的催化動力學參數Table 7 Kinetic parameters of WT and F172Y
2.5 亞胺還原酶母本與突變體的反應進程對比
通過對母本與突變體F172Y 在30 ℃、1 mmol/L底物濃度條件下的反應進程表征,本文得到了兩者的反應進程曲線,結果如圖6 所示。
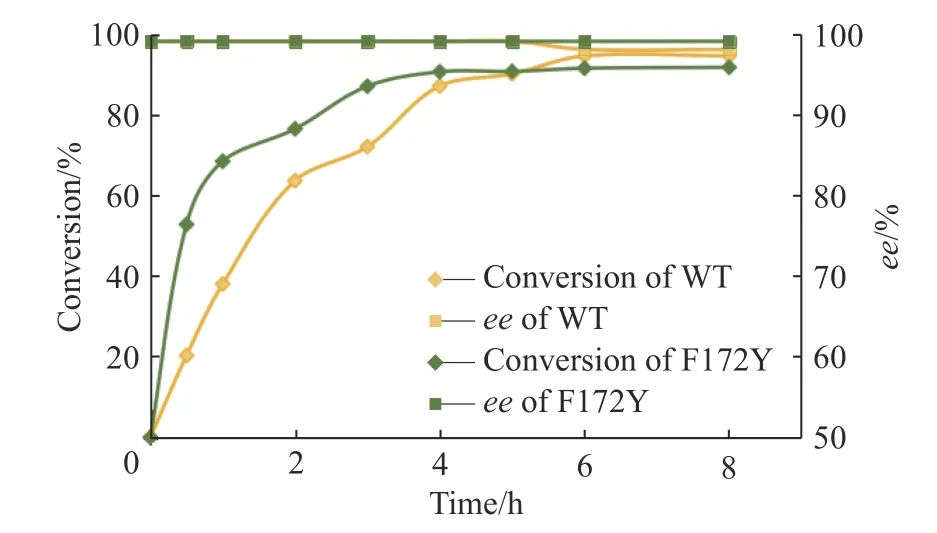
圖6 母本(黃色)與突變體F172Y(綠色)的反應進程曲線Fig. 6 Reaction process curves of WT (yellow) and F172Y(green) of AdIR1
以上結果表明,母本可以在6 h 實現94%的轉化率,產物ee值為98%。而突變體F172Y 并沒有因為點位突變而影響其對映體選擇性,其在前3 h 基本完成轉化,實現90%的轉化率和99%的ee值,后續過程中轉化率慢慢提升至92%,ee值始終保持在99%。
2.6 1-苯基-1,2,3,4-四氫異喹啉的克級制備反應
2.6.1 制備反應的最適溫度篩選 前期的酶學性質表征結果表明突變體F172Y 催化合成1-苯基-1,2,3,4-四氫異喹啉的最適催化溫度為40 ℃。由于制備級反應的底物濃度提升到了5 mmol/L,在此濃度下本文考察了突變體F172Y 不同溫度下催化合成1-苯基-1,2,3,4-四氫異喹啉的反應效果,結果如表8 所示,在1 mL 反應體系中,酶在40 ℃條件下的確表現出了比30 ℃更高的轉化率,其在40 ℃反應6 h 的結果與30 ℃反應12 h 的結果一致。因此,確定40 ℃為制備反應的溫度條件。
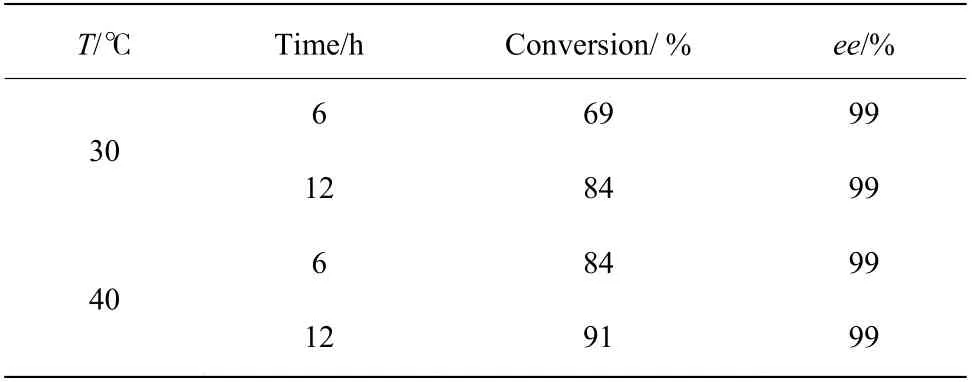
表8 F172Y 在30 ℃與40 ℃下的反應轉化結果Table 8 Reaction conversion results of F172Y at 30 ℃ and 40 ℃
2.6.2 最適助溶劑體積分數篩選結果 除了最適溫度,本文還對最適助溶劑體積分數進行了考察。為了降低DMSO 加入量的誤差,反應體系擴大到了10 mL級別,結果如圖7 所示。結果表明, φ =2%的DMSO加入量的轉化效果略優于1%與5%,且遠優于10%。因此將 φ =2%的DMSO 確定為最適助溶劑。另外,該反應條件下4 h 便完成了92%的轉化,該結果高于最適溫度篩選中6 h 的轉化率,原因可能是10 mL 反應器相對于振蕩反應器具有更佳的傳質、傳熱效果。

圖7 不同助溶劑濃度下的反應曲線Fig. 7 Reaction curves with different cosolvent volume fractions
2.6.3 克級制備反應 經過以上的反應條件優化,本文將反應體系擴大到2 L,在5 L 發酵罐中開展反應,底物1-苯基-3,4 二氫異喹啉的上載量為10 mmol(2.05 g),反應進程曲線如圖8 所示。由圖8 可以看出,12 h 后反應轉化率僅維持在85%,且基本不再變化;反應24 h 后對反應進行淬滅,通過柱層析純化得到0.34 g 的(S)-1-苯基-1,2,3,4-四氫異喹啉產物(得率為16.4%)。
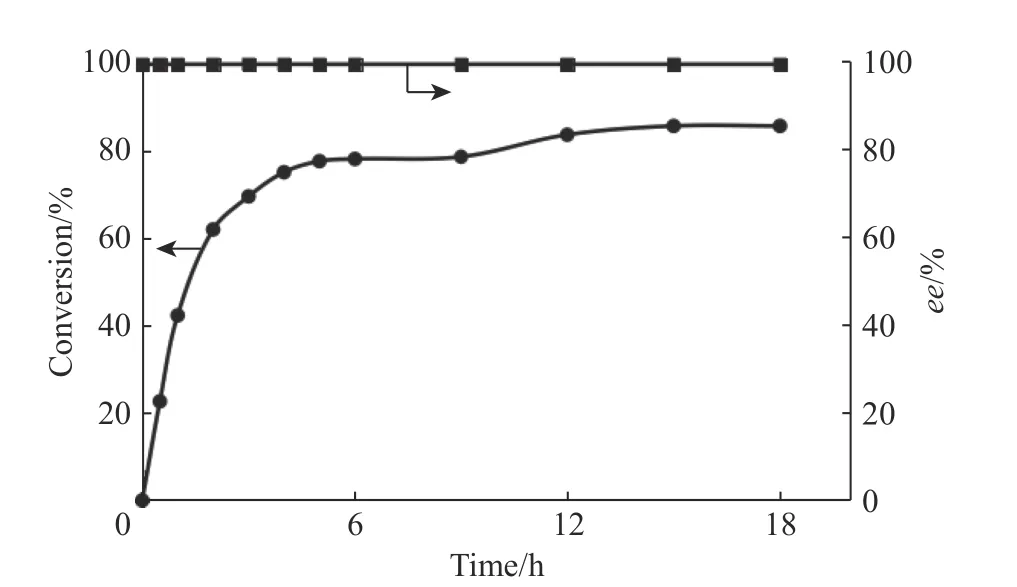
圖8 克級制備反應的反應進程曲線Fig. 8 Reaction process curves of gram-scale preparative reaction
3 結 論
以反應轉化率和對映體選擇性結果較優的AdIR1 作為分子改造的起始酶,得到了突變體F172Y,其在30 ℃條件下酶活為15.3 mU/mg,相較于母本提升了1.9 倍,催化效率(kcat/Km)相較于母本有3.4 倍的提升。同時突變體F172Y 具有更優的熱穩定性,在30 ℃和40 ℃條件下的t1/2分別是母本4.6 倍和6.3倍。另外,母本與突變體都未發現底物抑制情況。在催化合成1-苯基-1,2,3,4-四氫異喹啉的反應進程中,突變體F172Y 具有更快的轉化效果,相較于母本需要6 h 基本完成反應轉化,其僅需要3 h,且ee值維持在99%。在最適反應條件(40 ℃, φ =2% DMSO)下,利用突變體F172Y 作為催化劑開展了S構型1-苯基-1,2,3,4-四氫異喹啉的克級制備反應,反應12 h后可達85%的轉化率,通過柱層析分離得到了0.34 g產物,分離得率為16.4%,展示出所開發的突變酶F172Y 不對稱合成手性1-苯基-1,2,3,4-四氫異喹啉的實用性。
