miR-92a靶向PTEN促進宮頸癌細胞增殖*
2022-08-29 02:34:18蔣佩佩蔡徐山張春利齊結華吳守樂樂江漫
國際檢驗醫學雜志 2022年16期
關鍵詞:研究
蔣佩佩,蔡徐山,2△,張春利,宦 宇,齊結華,吳守樂,樂江漫
1.上海市嘉定區婦幼保健院檢驗科,上海 201821;2.同濟大學生命科學與技術學院,上海 200092
宮頸癌是最常見的婦科惡性腫瘤之一,其發病率僅次于乳腺癌、結直腸癌和肺癌[1]。據世界衛生組織不完全統計,全球每年新增宮頸癌病例50多萬例,發病率逐年上升,并呈年輕化趨勢,宮頸癌每年造成約30萬人死亡,其中85%發生在發展中國家,已嚴重影響女性的生存質量[2]。因此,研究宮頸癌的發病機制,尋找潛在的預后和靶向治療指標對提高宮頸癌患者的生存率具有重要意義。
微小RNA(miRNA)是在真核細胞及病毒體內發現的一類內源性染色體上的非編碼小分子單鏈RNA,長度為14~22個核苷酸[3]。miRNA通過與靶mRNA分子的3′-非翻譯區(3′-UTR)互補序列結合誘導mRNA 降解或抑制翻譯[4],調控基因表達。有研究表明,miRNA廣泛參與細胞發育、物質與能量代謝、免疫及炎性反應等各種生命代謝活動[5-8]。miR-92a屬于miR-17-92基因簇,據報道,miR-92a參與胃癌、骨肉瘤、鼻咽癌、肺癌、前列腺癌、結直腸癌和肝癌等多種腫瘤的發生和發展[9-13]。有研究表明,在宮頸癌組織、患者血清及細胞系中miR-92a均高表達[14-15]。ZHOU等[15]研究顯示,miR-92a可通過靶向抑癌基因FBXW7調控宮頸癌細胞周期,促進細胞增殖,增強細胞侵襲力。動物實驗證實,miR-92a可通過絲裂原活化蛋白激酶(MAPK)/細胞外信號調節激酶(ERK)途徑降低小鼠免疫力,增強宮頸成瘤能力[16]。
據生物信息學預測和功能分析,磷脂酰肌醇3激酶(PI3K)/蛋白激酶B(AKT)途徑的負調節因子第10號染色體同源丟失性磷酸酶張力蛋白基因(PTEN)是miR-92a的可能靶標[12]。然而,在宮頸癌中miR-92a經該通路靶向PTEN的調節作用鮮見報道。本研究通過體外干預宮頸癌細胞系miR-92a水平,初步探索其在PTEN/PI3K/AKT通路中對宮頸癌細胞的影響,并探討相關分子機制。
1 材料與方法
1.1材料 宮頸癌細胞HeLa和SiHa(上海攸碧艾細胞庫);DMEM-高糖培養基、胎牛血清(FBS)和胰蛋白酶(美國Gibco公司);逆轉錄試劑盒和PCR定量試劑盒(TaKaRa公司);抗磷酸化蛋白激酶B(p-AKT)、AKT、PTEN、GAPDH抗體(美國CST);LipofectamineTM2000(Lip2000,美國Invitrogen公司)。miR-92a-3p inhibitor及其陰性對照、PTEN、內參U6和GAPDH引物由上海生工生物工程有限公司合成。
1.2細胞培養 從液氮中取出HeLa與SiHa細胞,快速復蘇后立即重懸于含有10%FBS和1%雙抗(青霉素與鏈霉素各100 U/mL)的DMEM-高糖培養基中。于37 ℃、5% CO2恒溫培養箱中進行細胞培養。細胞隔天更換一次培養基,細胞長至密度80%~90%時進行細胞傳代。
1.3細胞分組與轉染 將細胞接種于培養板并用不含血清與雙抗的培養基培養,待細胞密度約60%時采用Lip2000進行RNA序列轉染,分別轉染miR-92a-3p inhibitor(inhibitor組)及其陰性對照(NC組),以未處理的宮頸癌細胞為空白對照組。轉染后將培養板置于37 ℃、5% CO2恒溫培養箱中培養,6 h后棄除含轉染液的培養基,更換為完全培養基,繼續培養。
1.4實時熒光定量PCR(RT-qPCR) 轉染48 h后提取總RNA。用磷酸鹽緩沖液(PBS)清洗細胞,加入Trizol 試劑,冰上裂解10 min,用移液槍吹打數次,充分裂解細胞后轉移至離心管中,加入氯仿,渦旋振蕩后4 ℃離心,取上層水相于離心管中,加入異丙醇后4 ℃離心,棄上清,收集RNA沉淀。沉淀用預冷75%乙醇洗滌后離心,棄上清,沉淀于冰上晾干,加入DEPC水充分溶解RNA。測定RNA濃度和純度。根據逆轉錄試劑盒逆轉錄miRNA(條件:42 ℃ 15 min,85 ℃ 5 s)和mRNA(條件:37 ℃ 15 min,85 ℃ 5 s),得到cDNA,然后以cDNA 為模板進行PCR反應。反應條件:95 ℃預變性30 s;95 ℃變性5 s,60 ℃退火延伸34 s,共40個循環。按照2-ΔΔCt公式,分別以GAPDH和U6為內參計算PTEN mRNA和miR-92a的相對表達量。基因及其引物序列見表1。

表1 引物序列
1.5細胞活率 采用CCK-8試劑盒檢測細胞活率。將細胞接種到96孔板中,待細胞過夜貼壁后進行轉染,分別于轉染24、48、72 h向每孔中加10 μL CCK-8試劑,每個時間段設3個復孔,放置培養箱內孵育1 h,用酶標儀測定450 nm處各組細胞的吸光度(A)。
1.6細胞凋亡率 采用Annexin V-FITC/PI雙染法聯合流式細胞儀檢測凋亡率。將每組細胞以2×105個/孔接種于六孔板,細胞培養24 h后行轉染,轉染后繼續培養。于培養36~48 h收集細胞,1×Binding Buffer洗滌3次,重懸細胞后加入5 μL Annexin V-FITC,室溫孵育15 min,上機前5 min加入5 μL PI。實驗重復3次。
1.7Western blotting檢測PTEN和磷酸化-AKT(p-AKT)蛋白表達 收集轉染48 h的各組細胞,加入PIPA裂解液冰上反應10 min,以低溫12 000 r/min離心10 min,提取上清蛋白裂解液。采用BCA法測定裂解液中蛋白濃度,調平蛋白濃度后加入上樣緩沖液,煮沸蛋白。行十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(SDS-PAGE)分離蛋白,濕轉法將蛋白轉至聚偏氟乙烯膜(PVDF膜)。2%牛血清清蛋白(BSA)封閉1 h,TBST洗膜3次。加入一抗(PTEN、p-AKT和GAPDH抗體的稀釋度均為1∶1 000),于4 ℃孵育過夜,加入熒光標記二抗室溫孵育1 h,TBST洗膜后置于紅外激光成像儀顯影。
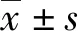
2 結 果
2.1RT-qPCR驗證轉染效率 HeLa 細胞中,inhibitor組miR-92a相對表達量(0.030±0.001)較NC組(0.980±0.007)明顯下降(P<0.01);SiHa細胞中,inhibitor組miR-92a相對表達量(0.010±0.001)較NC組(0.990±0.006)明顯下降(P<0.01),說明HeLa 細胞、SiHa細胞中miR-92a-3p inhibitor均轉染成功,可用于后續實驗。見圖1。

注:ns表示與空白對照組比較,差異無統計學意義(P>0.01);與NC組比較,aP<0.01。
2.2PTEN mRNA相對表達量 HeLa細胞中,inhibitor組PTEN mRNA相對表達量(1.830±0.050)較NC組(1.090±0.050)升高(P<0.01)。SiHa 細胞中,inhibitor組PTEN mRNA的相對表達量(1.670±0.040)較NC組(1.040±0.030)升高(P<0.01)。見圖2。
2.3抑制miR-92a表達對宮頸癌細胞活率的影響 CCK-8試驗結果顯示,與空白對照組相比,inhibitor組在轉染48、72 h細胞活率均降低(P<0.05),而NC組和空白對照組間差異無統計學意義(P>0.05)。見圖3。

注:ns表示與空白對照組比較,差異無統計學意義(P>0.01);與NC組比較,aP<0.01。

注:與NC組轉染48 h時比較,aP<0.05;與NC組轉染72 h時比較,aP<0.01。
2.4抑制miR-92a表達對宮頸癌細胞凋亡的影響 HeLa 細胞中,inhibitor組的細胞凋亡率[(16.54±0.78)%]高于空白對照組[(4.56±0.64)%]和NC組[(5.97±0.34)%],差異均有統計學意義(P<0.01),NC組和空白對照組比較,差異無統計學意義(P>0.05)。SiHa細胞中,inhibitor組的細胞凋亡率[(15.69±0.64)%]高于空白對照組[(4.37±0.74)%]和NC組[(5.32±0.41)%],差異均有統計學意義(P<0.01),NC組和空白對照組比較,差異無統計學意義(P>0.05)。見圖4。

圖4 抑制miR-92a表達后SiHa和HeLa 細胞凋亡檢測結果
2.5抑制miR-92a表達對宮頸癌細胞PTEN、p-AKT表達的影響 Western blotting檢測結果顯示,相比空白對照組,inhibitor組 PTEN蛋白表達升高,p-AKT的表達下降,NC組蛋白表達無明顯變化。見圖5。

圖5 抑制miR-92a表達后SiHa和HeLa細胞PTEN和p-AKT蛋白表達情況
3 討 論
miRNA是一類調控基因表達的非編碼RNA,僅占蛋白編碼基因的1%,而調節約1/3基因的表達[17]。生物信息學分析表明,一個miRNA有數百個靶基因,同時涉及多個信號通路,miRNA可能參與幾乎所有生命代謝活動[18]。有研究表明,位于腫瘤相關基因組區域的miRNA可以發揮癌基因或抑癌基因作用。
宮頸癌的發生、發展是多因素參與、多階段的過程,人乳頭瘤病毒(HPV)感染是其最重要的發病因素,miRNA在此過程中也發揮了重要作用。HPV-DNA整合到宿主染色體引起染色體結構不穩定及病毒致瘤蛋白的作用可能使miRNA的表達模式發生改變。miRNA的基因多態性和甲基化修飾也可增加癌癥的易感性[19]。此外,任何參與miRNA 加工的蛋白質的改變都可能干擾前體miRNA的有效成熟和(或)影響miRNA變體(isomiR)的生物發生,并可能徹底改變miRNA的功能。miRNA加工蛋白的失調已在HPV誘導的早期癌癥中被觀察到,如Drosha、AGO2和TENT2等[20],這表明miRNA可能參與了早期宮頸癌的發展。
有研究證實,miRNA在宮頸癌中表達失調,一些miRNA可促進宮頸癌的發生、發展,如miR-17、miR-21、miR-135、miR-150、miR-155、miR-196、miR-378、miR-425[21]。作為miR-17-92基因簇家族中的一員,miR-92a在多種腫瘤細胞的增殖、侵襲和轉移能力上扮演了不可或缺的角色。宮頸癌的體外實驗和動物實驗表明,miR-92a可經由相關信號通路和靶向基因促進細胞增殖,增強侵襲力,發揮致癌作用[15-16],但相關機制報道甚少。
為探討miR-92a在宮頸癌細胞中的作用,本研究向宮頸腺癌HeLa和鱗癌SiHa細胞轉染miR-92a inhibitor,RT-qPCR結果表明,miR-92a受到有效抑制。本研究中,CCK-8試驗和流式細胞術檢測結果顯示,抑制miR-92a表達后,宮頸癌細胞的活率下降,凋亡率升高,提示miR-92a對宮頸癌發生、發展具有潛在的促進作用。SU等[22]研究進一步說明,miR-92a可通過抑制p21的表達和促進細胞周期進程來促進宮頸癌細胞的增殖,突出了miR-92a在宮頸癌中的臨床意義。
PI3K/AKT通路是重要的細胞增殖信號通路,PTEN蛋白可使PI3K的產物三磷酸磷脂酰肌醇(PIP3)脫磷酸,從而下調該通路來維持生物體內細胞增殖與凋亡的動態平衡。PTEN/PI3K/AKT通路的失調已被證實參與多種腫瘤的發生、發展[9-13]。生物信息學預測PTEN是miR-92a的可能靶標,為了進一步探討miR-92a是否通過PTEN/PI3K/AKT通路對宮頸癌細胞增殖與凋亡發揮作用,本研究通過抑制miR-92a表達后觀察PTEN mRNA的表達情況,結果發現PTEN mRNA相對表達量升高,此結果與LI等[16]研究宮頸荷瘤小鼠的癌細胞轉染實驗結果一致。另外,本研究通過Western blotting實驗發現,抑制miR-92a表達后PTEN蛋白表達量增加,p-AKT蛋白表達量降低。本研究結果表明,miR-92a可能靶向PTEN調控PI3K/AKT信號通路,miR-92a在宮頸癌中有類似癌基因的作用。隨后,本課題組嘗試在宮頸癌細胞中過表達miR-92a以期進一步驗證其作用,但由于miR-92a過表達效果不理想,尚未得出有效的研究結果。
綜上所述,抑制miR-92a的表達可抑制宮頸癌細胞的增殖,促進其凋亡。miR-92a可能通過負向調控PTEN/PI3K/AKT信號通路實現類似癌基因的作用。miR-92a有可能成為宮頸癌治療的潛在靶點。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
